
Samy Ali Hussein Aziza
Department of Biochemistry, Faculty of Veterinary Medicine, Benha University, Moshtohor, Egypt
Mohammed El-said Azab
Department of Physiology, Faculty of Veterinary Medicine, Benha University, Moshtohor, Egypt
Soheir Kamal El-Shall
Department of Biochemistry, Faculty of Veterinary Medicine, Benha University, Moshtohor, Egypt
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 8 | Page No.: 964-977







ABSTRACT
Iron is an essential element that participates in several metabolic activities of cells; however, excess iron is a major cause of iron-induced oxidative stress and several human diseases. Natural flavonoids, as rutin, are well-known antioxidants and could be efficient protective agents. Therefore, the present study was undertaken to evaluate the protective influence of rutin supplementation to improve rat antioxidant systems against IOL-induced hepatic oxidative stress. Sixty male albino rats were randomly divided to three equal groups. The first group, the control, the second group, iron overload group, the third group was used as iron overload+rutin group. Rats received six doses of ferric hydroxide polymaltose (100 mg kg-1 b.wt.) as one dose every two days, by intraperitoneal injections (IP) and administerated rutin (50 mg kg-1 b.wt.) as one daily oral dose until the sacrificed day. Blood samples for serum separation and liver tissue specimens were collected three times, after three, four and five weeks from the onset of the experiment. Serum iron profiles total iron, Total Iron Binding Capacity (TIBC), Unsaturated Iron Binding Capacity (UIBC), transferrin (Tf) and Transferrin Saturation% (TS%)}, ferritin, albumin, total Protein, total cholesterol, triacylglycerols levels and aspartate aminotransferase (AST) and alanine aminotransferase (ALT) activities were determined. Moreover, total iron in the liver, L-malondialdehyde (L-MDA), glutathione (GSH), Nitric Oxide (NO) and Total Nucleic Acid (TNA) levels and glutathione peroxidase (GPx), catalase (CAT) and superoxide dismutase (SOD) activities were also determined. The obtained results revealed that, iron overload (IOL) resulted in significant increase in serum iron, TIBC, Tf, TS% and ferritin levels and AST and ALT activities and also increased liver iron, L-MDA and NO levels. Meanwhile, it decreased serum UIBC, total cholesterol, triacylglycerols, albumin, total protein and liver GSH, TNA levels and Gpx, CAT and SOD activities when compared with the control group. Rutin administration to iron-overloaded rats resulted in significant decrease in serum total iron, TIBC, Tf, TS%, ferritin levels and AST and ALT activities and liver total iron, L-MDA and NO levels with significant increases in serum UIBC, albumin, total protein and total cholesterol levels and in liver GSH, CAT and SOD activities compared with the IOL group. This study provides in vivo evidence that rutin administration can improve the antioxidant defence systems against IOL-induced hepatic oxidative stress in rats. This protective effect in liver of iron-loaded rats may be due to both antioxidant and metal chelation activities.
PDF Abstract XML References Citation
How to cite this article
Samy Ali Hussein Aziza, Mohammed El-said Azab and Soheir Kamal El-Shall, 2014. Ameliorating Role of Rutin on Oxidative Stress Induced by Iron Overload in Hepatic Tissue of Rats. Pakistan Journal of Biological Sciences, 17: 964-977.
DOI: 10.3923/pjbs.2014.964.977
URL: https://scialert.net/abstract/?doi=pjbs.2014.964.977
DOI: 10.3923/pjbs.2014.964.977
URL: https://scialert.net/abstract/?doi=pjbs.2014.964.977
INTRODUCTION
Iron is an essential micronutrient for all living organisms and is involved in fundamental enzymatic reactions, such as oxygen metabolism, electron-transfer processes and synthesis of DNA and RNA (Galleano et al., 2004). In the human body excessive amounts of iron may become very toxic due to it lacks effective mechanisms to protect cells against iron overload (Siah et al., 2005).
Iron overload is one of the most common metal-related toxicity (Zhao et al., 2005). It may be caused by: (1) Defects in iron absorption, as increase in iron absorption from the diet as in hereditary hemochromatosis (Beutler, 2007), in which genetic defects alter the regular absorption of iron from the gut or due to iron excess in diet as in Bantu siderosis (Gordeuk et al., 2003) (2) Parenteral iron administration in transfusion-dependent anaemias, as β-thalassemia; (3) Pathological conditions characterized by increases in iron (Crisponi and Remelli, 2008).
The liver is the main storage organ for iron, in iron overload, free radical formation and generation of lipid peroxidation (LPO) products, as iron excess generate oxidative stress through an increase rate of HO● generation by the Haber-Weiss reaction (Halliwell and Gutteridge, 1984), may result in progressive tissue injury as fibrosis (Arezzini et al., 2003) and eventually cirrhosis or hepatocellular carcinoma (Siah et al., 2005). In hepatocytes, as excessive iron deposition also lead to further injuries such as hepatocellular necrosis (Olynyk et al., 1995). Cases of acute iron toxicity are rare and mostly related to hepatoxicity (Tenenbein, 2001), which in turn cause the oxidation of lipids, proteins, nucleic acids (Papanastasiou et al., 2000) that may affect membrane fluidity and permeability and subsequently cell structure, function and viability (Maaroufi et al., 2011).
Iron-removal therapy may be achieved by antioxidants, iron chelators and/or free radical scavenging compounds, as flavonoids (polyphenols) (Blache et al., 2002), they exert multiple biological effects, including antioxidant and free radical scavenging activities (Negre-Salvayre and Salvayre, 1992), in which the metal chelation can be responsible, partly, both for the antioxidant capacity and some other activities (e.g., inhibition of lipooxygenases by chelation/reduction of iron in their active sites) (Vasquez-Martinez et al., 2007). Rutin is a kind of flavonoid glycoside known as vitamin P, a polyphenolic compound that is widely distributed in vegetables and fruits (Hertog et al., 1993), it is very effective free radical inhibitor in animal and human pathological states, such as IOL in rats (Afanas’ev et al., 1995), including hepatoprotective (Janbaz et al., 2002), in which its administration sharply suppressed free radical production in liver microsomes and by phagocytes in IOL animals (Afanas’ev et al., 1995) and also could reduce iron content in mouse liver (Gao et al., 2002). Accordingly, the aim of the present study was to evaluate the protective role of rutin administration against hepatic oxidative stress induced in iron-loaded rat models.
MATERIALS AND METHODS
Experimental animals: Sixty white male albino rats of 8-10 weeks old and weighing 180-220 g were used in the experimental investigation of this study. Rats were obtained from the Laboratory Animals Research Center, Faculty of Veterinary Medicine, Benha University, housed in separated metal cages and kept at constant environmental and nutritional conditions throughout the period of the experiment. The animals provided with a constant supply of standard pellet diet and fresh, clean drinking water ad-libitum.
Drug and antioxidants: The drug and antioxidant compounds used in the present study were:
| • | Haemojet(R): Haemojet ampoules were produced by Amriya Pharm. Ind. for European Egyptian Pharma. Ind., Alexandria, Egypt. Haemojet was obtained as pack of three ampoules of two ml solution. Each ampoule contains elemental iron (100 mg) as ferric hydroxide polymaltose complex |
| • | Rutin: Rutin is pale yellow crystalline powder (purity~99%). It was purchased from EIPICO ‘Egyption Pharmaceutical Industries Company, 10th of Ramadan city, Egypt. Rutin was dissolved in propylene glycol and administered to animals in daily oral dose of 50 mg kg-1 (b.wt.) (Fernandes et al., 2010) |
Induction of iron overload: Iron overload was induced by intraperitoneal injections of six doses (three doses per week) of ferric hydroxide polymaltose complex of 100 mg kg-1 b.w. (Zhao et al., 2005).
Experimental design: Rats were randomly divided into three equal groups: Each group containe twenty rats as follows:
| Group I: | (control group): Received saline only and served as control for all other groups |
| Group II: | (iron overload): Received six doses (three doses per week) of 100 mg kg-1 b.wt. of ferric hydroxide polymaltose administerated as IP-injections |
| Group III: | (iron overload+rutin): Received six doses (three doses per week) of 100 mg kg-1 b.wt. of ferric hydroxide polymaltose by IP-injections, followed by daily oral administration of rutin at a dose level of 50 mg kg-1b.wt. until the sacrificed day |
Sampling: Random blood samples and liver tissue specimens were collected from all animals groups (control and experimental groups) three times along the duration of experiment at three, four and five weeks from the onset of rutin administration.
Blood samples: Blood samples were collected by ocular vein puncture in dry, clean and screw capped tubes and serum was separated by centrifugation at 2500 rpm for 15 min. The clean, clear serum was separated and received in dry sterile samples tube, then kept in a deep freeze at -20°C until used for subsequent biochemical analysis
Liver tissues: At the end of the each experimental period, rats were sacrificed by cervical decapitation. The liver specimen was quickly removed and weighted, then perfused with cold saline to exclude the blood cells and then blotted on filter study and stored at -20°C. Briefly, half of liver tissues were cut, weighed and minced into small pieces, homogenized with a glass homogenizer in 9 volume of ice-cold 0.05 mM potassium phosphate buffer (pH7.4) to make 10% homogenates. The homogenates were centrifuged at 5,000 rpm for 15 min at 4°C then the supernatant was used for the subsequent biochemical analysis.
The other half of livers were weighed and putted into glass flask, then 5 volumes of mixed acid (nitric acid: Perchloric acid, 4:1) were added, heated until large amount of white vapors could be seen. The volumes of the digested samples were adjusted to 10 mL with double distilled water, then the obtained solutions were used to analyze iron contents.
Biochemical analysis: Serum iron and total iron binding capacity (TIBC), ferritin, albumin, total protein, total cholesterol, triacylglycerols levels and aspartate aminotransferase (AST) and alanine aminotransferase (ALT) activities were determined according to the methods described by Makino (1988), Dawson et al. (1992), Young (2001), Gendler (1984), Goodman et al. (1988), Stein (1987) and Young (2001), respectively.
Moreover, total iron in the liver, L-malondialdehyde (L-MDA), reduced glutathione (GSH), Nitric Oxide (NO) and Total Nucleic Acid (TNA) levels and Glutathione Peroxidase (GPx), catalase (CAT) and Superoxide Dismutase (SOD) activities were determined according to the methods described by Parker et al. (1967), Esterbauer et al. (1982), Beutler (1957), Montgomery and Dymock (1961), Spirin (1958), Gross et al. (1967), Sinha (1972) and Packer and Glazer (1990), respectively.
Statistical analysis: The obtained data were statistically analyzed by one-way analysis of variance (ANOVA) followed by the Duncan multiple test. All analyses were performed using the statistical package for social science 13.0 software, (SPSS, 2009). Values of p<0.05 were considered to be significant.
RESULT
The results presented in Table 1 and 2 revealed that, iron overload resulted in significant increase in serum iron, TIBC, Tf, TS% and ferritin levels and AST and ALT activities and also increased liver iron, L-MDA and NO levels. Meanwhile, it decreased serum UIBC, total cholesterol, triacylglycerols, total protein and albumin and liver GSH and TNA levels and Gpx, CAT and SOD activities, compared with the control group. Rutin administration to iron-loaded rats resulted in significant decreased in serum iron, TIBC, Tf, TS%, ferritin levels, AST and ALT activities and liver iron, L-MDA and NO levels, with significant increases in serum UIBC, total protein, albumin and total cholesterol levels and in liver GSH level, CAT and SOD activities, compared with the IOL group.
DISCUSSION
Iron overload in rats is an excellent model to study the in vivo LPO in which excess iron induced oxidative stress by increasing lipid peroxide levels in liver and in serum (Reddy and Lokesh, 1996). Subsequently, MDA and 8-IP adducts that were formed, significantly contributed to liver damage that is assessed by AST and ALT levels in the iron-supplemented rats (Asare et al., 2006). Excess hepatic iron may thus cause peroxidation of membrane lipids and oxidative liver injury (Abel and Gelderblom, 1998) in which IOL enhances liver injury and accelerates the process of fibrosis (Arezzini et al., 2003). This tissue injury can be relieved by the administration of an appropriate chelating agent which can combine with the iron and increase its rate of excretion (Zhao et al., 2005), as flavonoids that are substances with both chelating and free radical scavenging properties (Fraga and Oteiza, 2002) thus, rutin may be a very useful medicine for treatment (Reddy and Lokesh, 1996).
Serum and liver iron and serum TIBC, Tf, TS% and ferritin levels were significantly elevated in the iron-loaded rats, while serum UIBC was significantly decreased (Table 1, 2). These results came in accordance with the data of Silva et al. (2008) that reported, serum iron and TS% was 75% higher in rats with iron-dextran treatment when compared with the untreated control group. Also, Junge et al. (2001) reported that, acute iron overload elicited significant enhancement in iron levels of rats livers and Nahdi et al. (2010) observed that, IOL elicited significant enhancement in serum iron and significant increase (>10-fold) in liver iron in rats, TS was more than 100%, that most certainly results in the presence of Non Transferrin Binding Iron (NTBI) catalyzing the formation of reactive radicals (Zhang et al., 2006). Moreover, Crisponi and Remelli (2008) found that, when the iron load increases, the Iron Binding Capacity (IBC) of serum Tf is exceeded and a NTBI fraction of plasma iron appears which generates free hydroxyl radicals and induces dangerous tissue damage. Additionaly, Theurl et al. (2005) reported that, liver ferritin levels were increased with prolonged iron challenge as iron initially accumulated in spleen macrophages with subsequent increase in macrophage ferroportin and ferritin expression.
| Table 1: | Effect of treatment with rutin on some biochemical blood parameters in iron overloaded male rats |
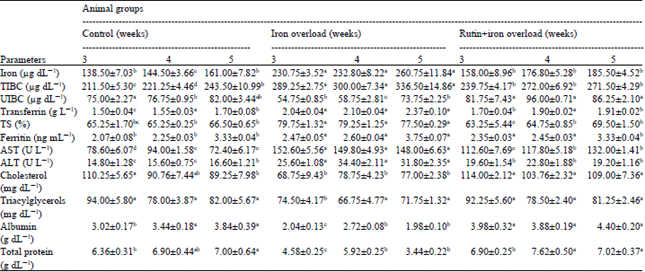 | |
| Data are presented as (Mean±SE), SE: Standard error, Mean values with different superscript letters in the same row are significantly different at (p<0.05) | |
Thus, iron overload treatment suggesting a novel mechanistic link between dopaminergic GSH depletion and increased Fe levels based on increased translational regulation of transferrin receptor 1(TfR1) (Kaur et al., 2009), in which iron deposition and related damages in liver indicate a strong relation between alterations of cellular redox condtion/increase in ROS generation due to GSH depletion with altered iron homeostasis in hepatic cell that led to iron deposition (Tapryal et al., 2010). However, exess iron induced increase in hepcidin mRNA level that was not sufficient to prevent increased intestinal iron absorption and onset of IOL compatible with the observation that serum iron was very high in that condition and TS was more than 100% that most certainly resulted in the presence of NTBI (Nahdi et al., 2010). Also, ferritin induction is expected to occur under conditions of chronic IOL when intracellular iron is abundant, due to lack of binding of the iron regulatory protein1 (IRP-1) to Iron-Responsive Elements (IRE) and IRP-2 degradation, resulting in significant translation of ferritin mRNA (Kim and Ponka, 2003). In cases of iron overload, the natural storage and transport proteins such as ferritin and transferrin become saturated and overwhelmed and then the iron spills over into other tissues and organs; At the same time, oxidative stress arises because of the catalytic activity of the metal ion on producing high reactive oxygen radicals and finally leads to tissue injury (Zhao et al., 2005).
When rats were administerated with rutin serum and liver iron and serum TIBC, Tf, TS% and ferritin concentrations were significantly decreased and serum UIBC was significantly increased than that of iron-loaded group (Table 1, 2). Similarly, Gao et al. (1999) found that, iron contents were significantly decreased in the liver of rutin and baicalin fed rat.
Also, Gao et al. (2002) reported that, oral adminstration of higher doses of rutin in mice can cause a decrease of serum iron, copper and zinc concentrations. In addition to Gao et al. (2003) that reported, the iron contents, in the liver of rutin or baicalin containing diet (1%) fed rats were significantly decreased. Moreover, Zhao et al. (2005) observed that, when iron-loaded mice were supplemented with baicalin, there was a decrease of hepatic total iron, at the same time, serum nonheme iron was significantly increased, indicating that baicalin could gradually combine with hepatic non-heme iron and finally excreted it from the body. Moreover, Zhang et al. (2006) found that, the increased NTBI in quercetin supplemented mice caused no further oxidation indicating that the increased serum non-heme iron may come from flavonoids chelated Fe and although serum ferritin level was still higher than that of normal mouse, it was significantly decreased compared with IOL-mouse. Furthermore, Kostyuk et al. (2004) observed that, quercetin effectively released Fe from ferritin and Fe-flavonoid complexes are demonstrated to be antioxidants. These results can be attributed to metal chelating effects of rutin, which are involved in the Fenton reaction (Arjumand et al., 2011) that can be responsible for the documented antioxidant capacity of flavonoids (Mladenka et al., 2011), as these chelation effects of flavonoids are structure specific (Gao et al., 2003) and suggests that the high reducing power and metal chelating activities mechanisms may play a key role in the inhibition of oxidative processes (Lue et al., 2010).
| Table 2: | Effect of treatment with rutin on some biochemical liver parameters in iron overloaded male rats |
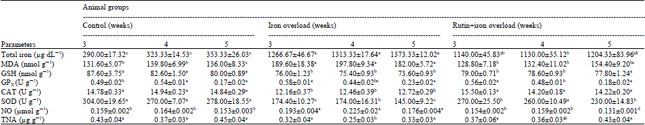 | |
| Data are presented as (Mean±SE), SE: Standard error, Mean values with different superscript letters in the same row are significantly different at (p<0.05) | |
However, although rutin form chelates with Fe ions, it is hydrolyzed by the intestinal flora to its corresponding aglycone, quercetin (Prince and Priya, 2010), which is responsible for its in vivo antioxidant activity, therefore, radical scavenging activity of rutin may be more important than their metal chelating activity (Kim et al., 2011).
Iron overload resulted in significant increase in serum AST and ALT activities compared with the normal control rats. The obtained results are nearly similar to data reported by Asare et al. (2006) that showed, in the iron-supplemented rats all of the indices of LPO, including AST and ALT, were increased significantly (~5-fold) compared with the control. Also, Silva et al. (2008) observed that, administration of iron dextran to rats increased AST activity, a marker for mitochondrial lesions and had no effect on ALT. As AST and ALT were used as sensitive indicators of liver damage (Mahmoud, 2012), the serum enzymes increased activities in iron-loaded rats can be attributed to the generation of ROS and oxidative damage by excess hepatic iron that may result in chronic necroinflammatory hepatic disease, which in turn generates more ROS and causes additional oxidative damage (Jungst et al., 2004). Rutin administration in iron-loaded rats decreased serum transaminases activities (Table 1). Similar results were reported by the data of Fernandes et al. (2010) that recorded, rutin administration to streptozotocin-diabetic rats decreased serum ALT and AST activities compared to untreated controls. Also, Mahmoud (2012) observed that, rutin administration significantly decreased the levels of AST and ALT activities in hyperammonemic rats, suggesting protection by preserving the structural integrity of the hepatocellular membrane against ammonium chloride. Rutin also scavenged free radicals (Cillard et al., 1990) and inhibiting LPO process (Karthick and Prince, 2006), in which the iron-rutin complexe studied not only retained the antioxidant properties of rutin, but in many cases exhibited enhanced free radical-scavenging activity (Ostrakhovitch and Afanas'ev, 2001).
The obtained results revealed that, iron overload resulted in significant decrease in serum total protein and albumin levels. The results agree well with those recorded by Asare et al. (2006) that found, iron accumulation disrupts the cell redox balance and geneates chronic oxidative stress, which damages DNA, lipids and protein in hepatocytes leading to both necrosis and apoptosis. That can be explained by protein oxidation that is known to give rise to alterations in both the backbone and side chains of the molecule, leading to the denaturation and loss of biological activities of various important proteins and cell death (Zhang et al., 2006), in which ROS may also cause oxidative damage to polyunsaturated fatty acids of membrane phospholipids, LPO enhanced by high-iron diet (Lafay et al., 2005), releasing cytotoxic and reactive aldehyde metabolites as MDA (Asare et al., 2006). Those cytotoxic products may impair cellular functions, including nucleotide and protein synthesis (Cheeseman, 1993). This suggestion was confirmed also by Youdim et al. (2005) that reported, ROS are capable of oxidizing cellular proteins, nucleic acids and lipids.
Rutin administration to iron-overloaded rats induced significant increases in serum total protein and albumin concentrations compared with the IOL-group. Similar results were reported by Zhang et al. (2006) that recorded ring the treatment with baicalin and quercetin in IOL-induced mice liver injury, the reduction of liver protein oxidation can be considered as a sign of protection under IOL. Kamalakkannan and Prince (2006) also observed that, oral administration of rutin to diabetic rats lead to significant increase in the plasma total protein and albumin concentrations when compared with the diabetic control. The antioxidant activity of rutin in Fenton reaction (Caillet et al., 2007) may explain the reduction of protein oxidation, as a mechanism of action preventing protein oxidation, in which the inhibitive effects of flavonoids on protein oxidation may come from the combination of both iron eliminating and direct free radical scavenging activities (Zhang et al., 2006).
Serum total cholesterol and triacyglycerols levels were significantly decreased in the iron loaded rats. These obtained results may be explained by Silva et al. (2008) that reported, hepatic injury triggered by iron excess may increase the concentration of secondary serum metabolites, such as cholesterol, triacylglycerols and glucose and also recorded that, treatment with Fe-dextran in male rats increased serum triacylglycerols level, but had no effect on the cholesterol evel. However, Turbino-Ribeiro et al. (2003) reported that, absence of alteration in serum cholesterol in rabbits receiving Fe-dextran injections.
When rats were administerated with rutin, serum total cholesterol and triacyglycerols levels were significantly increased than that of iron loaded control group. Contradictory results were obtained by Fernandes et al. (2010) reported that, serum total cholesterol and Low Density Lipoprotein (LDL)-cholesterol levels were lowered in the rutin-treated diabetics rats group. Also, Park et al. (2002) recorded that, supplementation of 0.1% rutin and tannic acid significantly lowered both plasma total cholesterol and triacylglycerols compared with control. These results may be explained by the lose of the amphiphilic properties of rutin, that be less capable of scavenging free radicals from the most lipophilic regions of the LDL particle (cholesterol esters and triacylglycerols) (Lue et al., 2010). In which the decrease in cholesterol level was due exclusively to the LDL and VLDL fraction (Aggarwal and Harikumar, 2009). On the other hand, Jiang et al. (2007) showed a dose-response effect of rutin in inhibiting LDL peroxidation and the reduced CAT activity in the rutin and tannic acid treatment might lead to less cholesteryl ester being available for VLDL packing, thereby resulting in a reduction in its secretion from the liver (Carr et al., 1992).
Liver L-MDA concentrations were significantly elevated in the iron-loaded rats as compared with control group. The obtained results are nearly similar to those of Kokoszko et al. (2008) who obseved that, the injection of FeCl or FeSO4 (3 mg Fe2+/100 g b.w.) significantly increased LPO products in the rat liver. Also, Nahdi et al. (2010) reported that, IOL in rats was accompanied by enhancement in LPO as shown by a significant increase in all tissue MDA concentration in iron supplemented group except the spleen and added that, in the liver there was a perfect correlation between MDA level and tissue iron content, suggesting production of oxidative stress. LPO generated by ROS was measured in terms of MDA (Arjumand et al., 2011), that is a secondary end product of the oxidation of polyunsaturated fatty acids (Yang et al., 2008). Focusing on the liver organ, the cytotoxic degradation products as MDA and 4-hydroxy-2-non-enal (4-HNE) (Esterbauer et al., 1991) can form covalent adducts with proteins, phospholipids and DNA (Guichardant et al., 1998), in which the formation of liver microsomal MDA protein adducts, during IOL in mice and the microsomal function impairment may alter protein function and might lead to cellular injury and Fe-associated hepatotoxicity (Valerio and Petersen, 1998). When rats were administerd with rutin hepatic L-MDA concentrations were significantly decreased than that of iron-loaded control group (Table 2). The obtained results agree with the data of Ostrakhovitch et al. (1995) that reported, rutin administration in IOL-rats sharply decreased microsomal LPO in the liver and spontaneous oxygen radical production by peritoneal macrophages. Also, Gao et al. (2003) reported that, LPO level in the liver of the rutin fed group was significantly decreased in comparison to the control group, As iron overload led to enhancement in LPO (Nahdi et al., 2010), some flavonoids can be both antioxidants and iron chelators; it means that flavonoids will be good candidates for curing IOL related diseases, as they can play a double role in reducing the rate of oxidation, one act as iron chelator (Borsari et al., 2001) and the other act as radical trap (Van Acker et al., 1998). Afanas’ev et al. (1995) also found that, rutin administration sharply suppressed free radical production in liver microsomes by phagocytes in iron overload rats. This antioxidant activity due to the inactive iron-rutin complexes of rutin that may be a good in vivo antioxidant and may be more effective free radical scavengers compared to the parent rutin (Ostrakhovitch and Afanas'ev, 2001). The sharp decrease in L-MDA in IOL-rats may be also attributed to the free radical scavenging property of rutin, in which IOL in rats induces the oxidative stress that was characterized by oxygen radical overproduction in liver microsomes, peritoneal macrophages and blood neutronphils (Mahmoud, 2012).
The obtained results revealed that, IOL resulted in significant decrease in liver GSH levels when compared with the control group. The obtained data are nearly similar to those reported by Poli et al. (2004) that observed, the increase in protein carbonylation and reduction in GSH content as well as in the GSH/GSSG ratio of the liver were observed after 6 weeks of treatment probably induced by iron-generated free radical activity (Pietrangelo, 2003). Also, Jagetia and Reddy (2011) reported that, introduction of Fe into mouse liver mitochondrial fraction caused a time dependent depletion in GSH (~2-fold) lower than control at 30 min post treatment. GSH is a major non-enzymatic tripeptide multifunctional intracellular antioxidant (Morimoto et al., 2008) that was often used as an estimation of the redox environment of the cell (Schafer and Buettner, 2001). Iron deposition and related damages in liver indicate a strong relation between alterations of cellular redox condtion/increase in ROS, e.g., Fe-induced free radical (Pietrangelo, 2003) due to GSH depletion, suggesting a novel mechanistic link between dopaminergic GSH depletion and increased iron levels (Kaur et al., 2009); In conclusion, reduced GSH functions intracellularly to reduce numerous oxidizing compounds, including ROS (Gong et al., 2010).
Rutin administration to iron-overloaded rats resulted in significant increase in liver GSH level compared with the IOL-group. Similarly, Korkmaz and Kolankaya (2010) observed that, rutin pretreatment significantly protected against the severe depletion of GSH content and MnSOD activity in the I/R-induced damage in rat kidney. Moreover, Arjumand et al. (2011) reported that, rutin treatment in cisplatin administered rats showed significant improvement in GSH concentration, suggesting its role in scavenging the free radicals generated by cisplatin-induced renal inflammation and apoptosis. The antioxidant imbalance was compensated by the prophylactic treatment of rutin as excessive LPO can cause increased GSH consumption (La Casa et al., 2000), this inhibitory effects of rutin on in vivo free radical production in IOL-rats are probably explained by its ability to form inactive iron-rutin complexes (Afanas'ev et al., 1989).
The obtained results revealed that, iron overload resulted in significant decrease in liver Gpx, CAT and SOD activities when compared with the control group. Iron overload can destruct the balance between prooxidants and antioxidants, leading to severe loss of total antioxidant status level. This phenomenon can be seen in most iron overload animal models (Dabbagh et al., 1994) as well as iron overload disease such as hereditary haemochromatosis (Young et al., 1994) and thalassemia (Livrea et al., 1996).
Catalase is an iron-containing antioxidant enzyme, it was reported that under iron overload, there was a significant decrease of catalase activity in rat liver (Galleano and Puntarulo, 1997). Likewise, Zhao et al. (2005) who showed that, Fe-dextran injection in mouse caused a significant decrease in hepatic CAT activity. Moreover, Valko et al. (2006) reported that, GPx significantly competes with CAT for H2O2 substrate and it is the major source of protection against low levels of oxidative stress-induced cancer. These results can be explained as the ROS generated during normal cellular processes are immediately detoxified by endogenous antioxidants like GSH, CAT, GPx, glutathione reductase, glutathione-S-transferase, etc. (Kim et al., 2006).
SOD work in conjunction with CAT and GPX (Michiels et al., 1994), preventing its interaction with Fe and therefore formation of the highly toxic ●OH, in which SOD and GPX are supportive enzyme system of the first line cellular defense against oxidative injury (Kalpravidh et al., 2010). GPx decomposes peroxides, that initiate a chain of free radical formation (Jagetia and Reddy, 2011) to H2O (or alcohol) while simultaneously oxidizing GSH; Significantly, GPx competes with CAT for H2O2 substrate, as it is the major source of protection against low levels of oxidative stress (Valko et al., 2006) and the increase of SOD/GPx ratio in Fe treated cells compared to control cells indicated that Fe-induced oxidative injury appeared, that might be indicative of ROS increase due to unefficient scavenging by enzymes (Formigari et al., 2007).
Rutin administration to iron-loaded rats resulted in significant increases in liver CAT and SOD activities compared with the IOL group (Table 2). As natural antioxidants, flavonoids intake may increase total antioxidant status level in living body, supplementation of baicalin or another flavonoid, rutin, could increase hepatic total antioxidant status level in rats and mice (Gao et al., 2003; Zhao et al., 2005). This effect may come from the chelation of free iron ion with stopping iron-catalyzed oxidative reaction and the direct increasing antioxidant status by baicalin and its metabolites that acte as strong antioxidant. Thus, the baicalin supplementation introduced a new source of antioxidant and could partly inhibit peroxidation-induced heme destruction and then provided a protection on catalase (Zhao et al., 2005). The significant increases in liver antioxidant enzymes in rutin treated iron-loaded rats are in conformity with the data reported by Park et al. (2002) that recorded, dietary rutin and tannic acid have a significant effect on SOD and GPx in rats. As a concomitant increase in CAT and/or GPx activity is essential if a beneficial effect from the high SOD be expected. Mahmoud (2012) found that, hyper-ammonemic rats pretreated with rutin significantly increased liver CAT and GPx. Chronic iron administration induced adaptive responses involving stimulation of the antioxidant defenses. Rutin significantly inhibited LPO in IOL-microsomes (Afanas’ev et al., 1995) that may be due to the free radical scavenging property (Mahmoud, 2012). The three enzymes can prevent damage by detoxifying ROS. The significant elevation of SOD activity also suggests that its free-radical scavenging activity is only effective when it is accompanied by an increase in activity of CAT and/or GPx activity, because SOD generates H2O2 as a metabolite (Park et al., 2002), which is more toxic than oxygen radicals in cells and needs to be scavenged by CAT or GPx (Pigeolet et al., 1990).
Iron overload resulted in significant increase in liver NO levels when compared with the control group. Similar results were recorded by Cornejo et al. (2001) that found, increased NO generation was evidenced in the liver under conditions of acute IOL. Moreover, Vadrot et al. (2006) explored that, increased production of NO by nitric oxide synthase-2 was demonstrated in patients with hepatic cell carcinoma complicating hereditary haemochromatosis. That can be explained as NO is an inorganic reactive nitrogen species synthesized in liver by inducible nitric oxide synthase (iNOS) found in hepatocytes, Kupffer cells and endothelial cells (Alderton et al., 2001), whose expression is controlled by the redox-sensitive transcription factor, nuclear factor-kappa B (Kleinert et al., 2004) and the complex interrelationships between Fe and NO (Galleano et al., 2004) can result in changes in in vivo NO production (Kagan et al., 2001). Also the increase in rat liver NOS activity due to chronic iron overload (Cornejo et al., 2007) is related to upregulation of iNOS expression (Cornejo et al., 2005). Rutin administration to iron-overloaded rats resulted in significant decreased liver NO levels compared with the IOL group. The data obtained are in harmony with Chen et al. (2001a, b) and Shen et al. (2002) that showed, rutin inhibit lipopolysaccharide-induced NO production. NO was proposed to act as a pro-oxidant at high conc. (Morand et al., 2000), or when it reacts with O2●B, forming the highly reactive peroxynitrite (Radi et al., 2001) that is suppressed by flavonoids by direct scavenging (Haenen et al., 1997).
Iron overload resulted in significant decreased in liver TNA levels when compared with the control group. Similarly, Youdim et al. (2005) explored that, iron is a major generator of ROS that lead to damage of lipids, proteins carbohydrates and nucleic acids. Park and Park (2011) reported that, Ferric-nitrilotriacetate markedly induced DNA damage in human leukocytes in vitro and rat leukocytes in vivo. The decrease in total nuclic acid level can be attributed to the primary generation of H2O2 and ●OH, due to Fe-induced oxidative stress, that damages DNA and other biomolecules (Huang, 2003). As ●OH may damage the nucleotide bases themselves, resulting in oxidized base products such as 8 oxo-guanine and fragmented or ring-opened derivatives (Lu et al., 2001). Iron also is thought to be involved in β-cleavage of lipid hydroperoxides, producing biogenic aldehydes that interact with DNA to form exocyclic products (Kew, 2009) that trigger free radical-mediated chain reaction including LPO, DNA damage and protein oxidation (Gutteridge and Halliwell, 2000).
Rutin administration to iron-overloaded rats, non significantly increased liver TNA concentration compared with the IOL-group. The obtained results are of the same harmony with the data of Undeger et al. (2004) that reported, rutin can prevent damage to DNA as it may modulate the enzymes necessary for activation of carcinogens. Also, Gong et al. (2010) observed that, rutin (50 μM) blocked H2O2-induced apoptosis in human umbilical vein endothelial cells and thus protecting DNA damage. Moreover, Omololu et al. (2011) reported that, rutin protect the stability of the genome. This protection against DNA damage is particularly important since oxidative damage to DNA, especially strand breaks, is highly dependent on the amount of Fe bound to DNA (Diaz-Castro et al., 2010), in which polyphenols, due to their ability to coordinate iron, are one large class of antioxidants that has been extensively examined for treatment and prevention of conditions associated with iron-generated ROS and oxidative stress (Perron and Brumaghim, 2009), as iron chelation was responsible for prevention of nuclear DNA damage by quercetin (Sestili et al., 1998).
CONCLUSION
From the obtained results it could be concluded that, the natural flavonoids, rutin inhibited the adverse effect of ferric ion induced oxidative stress by reducing protein and DNA oxidation and inhibition of lipid peroxidation in liver tissue due to its marked hepatoprotective role in the experimental rats. Rutin treatment improved the cytoprotective enzymatic and non-enzymatic antioxidants as revealead by elevated the GSH concentration, GPx, CAT and SOD activities and thus, might protect the cellular environments from iron-induced free radical damage. The results indicate that, rutin may have potential effects in inhibiting the iron-induced oxidative stress in human. The protective effect of rutin on livers of iron-overloaded rats may be due to its high antioxidant activity, including both its radical scavenging and iron chelation activities. So we recommended using rutin-enriched food regularly with additional research work for medicine manufacturing for protection against the bad complications of IOL-induced oxidative stress.
REFERENCES
- Abel, S. and W.C. Gelderblom, 1998. Oxidative damage and fumonisin B1-induced toxicity in primary rat hepatocytes and rat liver in vivo. Toxicology, 131: 121-131.
CrossRefPubMedDirect Link - Ostrakhovitch, E.A. and I.B. Afanas'ev, 2001. Oxidative stress in rheumatoid arthritis leukocytes: Suppression by rutin and other antioxidants and chelators. Biochem. Pharmacol., 62: 743-746.
CrossRef - Afanas'ev, I.B., A.I. Dorozhko, A.V. Brodshi, V.A. Kostyak and A.I. Potaporitch, 1989. Chelating and free radical scavenging mechanisms of inhibitory action of rutin and quercetin in lipid peroxidation. Biochem. Pharmacol., 38: 1763-1769.
PubMed - Afanas'ev, I.B., E.A. Ostrachovitch, N.E. Abramova and L.G. Korkina, 1995. Different antioxidant activities of bioflavonoid rutin in normal and iron-overloading rats. Biochem. Pharmacol., 50: 621-635.
CrossRefPubMedDirect Link - Aggarwal, B.B. and K.B. Harikumar, 2009. Potential therapeutic effects of curcumin, the anti-inflammatory agent, against neurodegenerative, cardiovascular, pulmonary, metabolic, autoimmune and neoplastic diseases. Int. J. Biochem. Cell Biol., 41: 40-59.
CrossRefDirect Link - Alderton, W.K., C.E. Cooper and R.G. Knowles, 2001. Nitric oxide synthases: Structure, function and inhibition. Biochem. J., 357: 593-615.
PubMedDirect Link - Arezzini, B., B. Lunghi, G. Lungarella and C. Gardi, 2003. Iron overload enhances the development of experimental liver cirrhosis in mice. Int. J. Biochem. Cell Biol., 35: 486-495.
CrossRefDirect Link - Arjumand, W., A. Seth and S. Sultana, 2011. Rutin attenuates cisplatin induced renal inflammation and apoptosis by reducing NFκB, TNF-α and caspase-3 expression in Wistar rats. Food Chem. Toxicol., 49: 2013-2021.
CrossRefPubMedDirect Link - Asare, G.A., K.S. Mossanda, M.C. Kew, A.C. Paterson, C.P. Kahler-Venter and K. Siziba, 2006. Hepatocellular carcinoma caused by iron overload: A possible mechanism of direct hepatocarcinogenicity. Toxicology, 219: 41-52.
CrossRefPubMedDirect Link - Junge, B., Y. Carrion, C. Bosco, M. Galleano, S. Puntarulo, G. Tapia and L.A. Videla, 2001. Effects of iron overload and lindane intoxication in relation to oxidative stress, kupffer cell Function and liver injury in the rat. Toxicol. Applied Pharmacol., 170: 23-28.
CrossRefPubMedDirect Link - Lue, B.M., N.S. Nielsen, C. Jacobsen, L. Hellgren, Z. Guo and X. Xu, 2010. Antioxidant properties of modified rutin esters by DPPH, reducing power, iron chelation and human low density lipoprotein assays. Food Chem., 123: 221-230.
CrossRefDirect Link - Beutler, E., 2007. Iron storage disease: Facts, fiction and progress. Blood Cells Mol. Dis., 39: 140-147.
CrossRefPubMedDirect Link - Blache, D., P. Durand, M. Prost and N. Loreau, 2002. (+)-Catechin inhibits platelet hyperactivity induced by an acute iron load in vivo. Free Radical Biol. Med., 33: 1670-1680.
CrossRefDirect Link - Borsari, M., C. Gabbi, F. Ghelfi, R. Grandi, M. Saladini, S. Severi and F. Borella, 2001. Silybin, a new iron-chelating agent. J. Inorganic Biochem., 85: 123-129.
CrossRefPubMedDirect Link - Caillet, S., H. Yu, S. Lessard, G. Lamoureux, D. Ajdukovic and M. Lacroxix, 2007. Fenton reaction applied for screening natural antioxidants. Food Chem., 100: 542-552.
CrossRefDirect Link - Carr, T.P., J.S. Parks and L.L. Rudel, 1992. Hepatic ACAT activity in African green monkeys is highly correlated to plasma LDL cholesteryl ester enrichment and coronary artery atherosclerosis. Arterioscler. Thromb., 12: 1274-1283.
CrossRefPubMedDirect Link - Chen, Y.C., S.C. Shen, W.R. Lee, W.C. Hou, L.L. Yang and T.J.F. Lee, 2001. Inhibition of nitric oxide synthase inhibitors and lipopolysaccharide induced inducible NOS and cyclooxygenase-2 gene expressions by rutin, quercetin and quercetin pentaacetate in RAW 264.7 macrophages. J. Cell. Biochem., 82: 537-548.
CrossRefPubMedDirect Link - Cornejo, P., P. Varela, L.A. Videla and V. Fernandez, 2005. Chronic iron overload enhances inducible nitric oxide synthase expression in rat liver. Nitric Oxide, 13: 54-61.
CrossRefPubMedDirect Link - Cornejo, P., G. Tapia, S. Puntarulo, M. Galleano, L.A. Videla and V. Fernandez, 2001. Iron-induced changes in nitric oxide and superoxide radical generation in rat liver after lindane or thyroid hormone treatment. Toxicol. Lett., 119: 87-93.
CrossRefPubMedDirect Link - Cornejo, P., V. Fernandez, M.T. Vial and L.A. Videla, 2007. Hepatoprotective role of nitric oxide in an experimental model of chronic iron overload. Nitric Oxide, 16: 143-149.
Direct Link - Crisponi, G. and M. Remelli, 2008. Iron chelating agents for the treatment of iron overload. Coordination Chem. Rev., 252: 1225-1240.
CrossRefDirect Link - Dawson, D.W., D.I. Fish and P. Shackleton, 1992. The accuracy and clinical interpretation of serum ferritin assays. Clin. Lab. Haematol., 14: 47-52.
Direct Link - Diaz-Castro, J., S. Hijano, M.J.M. Alferez, I. Lopez-Aliaga, T. Nestares, M. Lopez-Frias and M.S. Campos, 2010. Goat milk consumption protects DNA against damage induced by chronic iron overload in anaemic rats. Int. Dairy J., 20: 495-499.
CrossRefDirect Link - Esterbauer, H., K.H. Cheeseman, M.U. Dianzani, G. Poli and T.F. Slater, 1982. Separation and characterization of the aldehydic products of lipid peroxidation stimulated by ADP-Fe2+ in rat liver microsomes. Biochem. J., 208: 129-140.
CrossRefPubMedDirect Link - Esterbauer, H., R.J. Schaur and H. Zollner, 1991. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med., 11: 81-128.
CrossRefPubMedDirect Link - Fernandes, A.A.H., E.L.B. Novelli, K. Okoshi, M.P. Okoshi, B.P. Di Muzio, J.F.C. Guimaraes and A. Fernandes Junior, 2010. Influence of rutin treatment on biochemical alterations in experimental diabetes. Biomed. Pharmacother., 64: 214-219.
CrossRefPubMedDirect Link - Formigari, A., P. Irato and A. Santon, 2007. Zinc, antioxidant systems and metallothionein in metal mediated-apoptosis: Biochemical and cytochemical aspects. Comp. Biochem. Physiol. C Toxicol. Pharmacol., 146: 443-459.
CrossRefPubMedDirect Link - Fraga, C.G. and P.I. Oteiza, 2002. Iron toxicity and antioxidant nutrients. Toxicology, 180: 23-32.
CrossRefDirect Link - Galleano, M. and S. Puntarulo, 1997. Dietary α-tocopherol supplementation on antioxidant defenses after in vivo iron overload in rats. Toxicology, 124: 73-81.
CrossRefDirect Link - Gao, Z., H. Xu and K. Huang, 2002. Effects of rutin supplementation on antioxidant status and iron, copper and zinc contents in mouse liver and brain. Biol. Trace Elem. Res., 88: 271-279.
CrossRefDirect Link - Gao, Z., K. Huang, X. Yang and H. Xu, 1999. Free radical scavenging and antioxidant activities of flavonoids extracted from the radix of Scutellaria baicalensis Georgi. Biochim. Biophys. Acta General Subjects, 1472: 643-650.
CrossRefPubMedDirect Link - Gao, Z., H. Xu, X. Chen and H. Chen, 2003. Antioxidant status and mineral contents in tissues of rutin and baicalin fed rats. Life Sci., 73: 1599-1607.
CrossRefPubMedDirect Link - Gong, G., Y. Qin, W. Huang, S. Zhou, X. Yang and D. Li, 2010. Rutin inhibits hydrogen Peroxide-induced apoptosis through regulating reactive oxygen species mediated mitochondrial dysfunction pathway in human umbilical vein endothelial cells. Eur. J. Pharmacol., 628: 27-35.
CrossRefPubMedDirect Link - Gordeuk, V.R., A. Caleffi, E. Corradini, F. Ferrara and R.A. Jones et al., 2003. Iron overload in Africans and African-Americans and a common mutation in the SCL40A1 (ferroportin 1) gene. Blood Cells Mol. Dis., 31: 299-304.
CrossRefPubMedDirect Link - Gross, R.T., R. Bracci, N. Rudolph, E. Schroeder and J.A. Kochen, 1967. Hydrogen peroxide toxicity and detoxification in the erythrocytes of newborn infants. Blood, 29: 481-493.
CrossRefPubMedDirect Link - Guichardant, M., P. Taibi-Tronche, L.B. Fay and M. Lagarde, 1998. Covalent modifications of aminophospholipids by 4-hydroxynonenal. Free Radical Biol. Med., 25: 1049-1056.
CrossRefPubMedDirect Link - Gutteridge, J.M.C. and B. Halliwell, 2000. Free radicals and antioxidants in the year 2000: A historical look to the future. Ann. N. Y. Acad. Sci., 899: 136-147.
CrossRefDirect Link - Halliwell, B. and J.M. Gutteridge, 1984. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J., 219: 1-14.
PubMed - Hertog, M.G., P.C. Hollman, M.B. Katan and D. Kromhout, 1993. Intake of potentially anticarcinogenic flavonoids and their determinants in adults in The Netherlands. Nutr. Cancer, 20: 21-29.
CrossRefPubMedDirect Link - Huang, X., 2003. Iron overload and its association with cancer risk in humans: Evidence for iron as a carcinogenic metal. Mutation Res./Fundam. Mol. Mech. Mutagen., 533: 153-171.
CrossRefDirect Link - Jagetia, G.C. and T.K. Reddy, 2011. Alleviation of iron induced oxidative stress by the grape fruit flavanone naringin in vitro. Chemico-Biol. Interact., 190: 121-128.
CrossRefDirect Link - Janbaz, K.H., S.A. Saeed and A.H. Gilani, 2002. Protective effect of rutin on paracetamol- and CCl4-induced hepatotoxicity in rodents. Fitoterapia, 73: 557-563.
CrossRefPubMedDirect Link - Jiang, P., F. Burczynski, C. Campbell, G. Pierce, J.A. Austria and C.J. Briggs, 2007. Rutin and flavonoid contents in three buckwheat species Fagopyrum esculentum, F. tataricum and F. homotropicum and their protective effects against lipid peroxidation. Food Res. Int., 40: 356-364.
CrossRefDirect Link - Jungst, C., B. Cheng, R. Gehrke, V. Schmitz and H. Dieter et al., 2004. Oxidative damage is increased in human liver tissue adjacent to hepatocellular carcinoma. Hepatology, 39: 1663-1672.
CrossRefDirect Link - Kagan, V.E., A.V. Kozlov, Y.Y. Tyurina, A.A. Shvedova and J.C. Yalowich, 2001. Antioxidant mechanisms of nitric oxide against iron-catalyzed oxidative stress in cells. Antioxidants Redox Signal., 3: 189-202.
PubMed - Kalpravidh, R.W., N. Siritanaratkul, P. Insain, R. Charoensakdi and N. Panichkul et al., 2010. Improvement in oxidative stress and antioxidant parameters in β-thalassemia/Hb E patients treated with curcuminoids. Clin. Biochem., 43: 424-429.
CrossRefDirect Link - Kamalakkannan, N. and P.S. Prince, 2006. Antihyperglycaemic and antioxidant effect of rutin, a polyphenolic flavonoid, in streptozotocin-induced diabetic Wistar rats. Basic Clin. Pharmacol. Toxicol., 98: 97-103.
CrossRefPubMedDirect Link - Karthick, M. and S.M.P. Prince, 2006. Preventive effect of rutin, a bioflavonoid, on lipid peroxides and antioxidants in isoproterenol-induced myocardial infarction in rats. J. Pharm. Pharmacol., 58: 701-707.
PubMed - Kaur, D., D. Lee, S. Ragapolan and J.K. Andersen, 2009. Glutathione depletion in immortalized Midbrain-derived dopaminergic neurons results in increases in the labile iron pool: Implications for Parkinson's disease. Free Radical Biol. Med., 46: 593-598.
CrossRefDirect Link - Kew, M.C., 2009. Hepatic iron overload and hepatocellular carcinoma. Cancer Lett., 286: 38-43.
CrossRefPubMedDirect Link - Kim, G.N., Y.I. Kwon and H.D. Jang, 2011. Protective mechanism of quercetin and rutin on 2, 2'-azobis (2-amidinopropane) dihydrochloride or Cu2+-induced oxidative stress in HepG2 cells. Toxicol. In vitro, 25: 138-144.
CrossRefDirect Link - Kim, S. and P. Ponka, 2003. Role of nitric oxide in cellular iron metabolism. Biometals, 16: 125-135.
CrossRefDirect Link - Kleinert, H., A. Pautz, K. Linker and P.M. Schwarz, 2004. Regulation of the expression of inducible nitric oxide synthase. Eur. J. Pharmacol., 500: 255-266.
CrossRefDirect Link - Korkmaz, A. and D. Kolankaya, 2010. Protective effect of rutin on the ischemia/reperfusion induced damage in rat kidney. J. Surg. Res., 164: 309-315.
CrossRefPubMedDirect Link - Kostyuk, V.A., A.I. Potapovich, E.N. Strigunova, T.V. Kostyuk and I.B. Afanas'ev, 2004. Experimental evidence that flavonoid metal complexes may act as mimics of superoxide dismutase. Arch. Biochem. Biophys., 428: 204-208.
CrossRefPubMedDirect Link - La Casa, C., I. Villegas, C.A. de la Lastra, V. Motilva and M.J.M. Calero, 2000. Evidence for protective and antioxidant properties of rutin, a natural flavone, against ethanol induced gastric lesions. J. Ethnopharmacol., 71: 45-53.
CrossRefPubMedDirect Link - Lafay, S., E. Gueux, Y. Rayssiguier, A. Mazur, C. Remesy and A. Scalbert, 2005. Caffeic acid inhibits oxidative Stress and reduces hypercholesterolemia induced by iron overload in rats. Int. J. Vitam. Nutr. Res., 75: 119-125.
PubMed - Livrea, M.A., L. Tesoriere, A.M. Pintaudi, A. Calabrese and A. Maggio et al., 1996. Oxidative stress and antioxidant status in β-thalassemia major: Iron overload and depletion of lipid-soluble antioxidants. Blood, 88: 3608-3614.
PubMedDirect Link - Lu, A.L., X. Li, Y. Gu, P.M. Wright and D.Y. Chang, 2001. Repair of oxidative DNA damage. Cell Biochem. Biophys., 35: 141-170.
CrossRefDirect Link - Maaroufi, K., E. Save, B. Poucet, M. Sakly, H. Abdelmelek and L. Had-Aissouni, 2011. Oxidative stress and prevention of the adaptive response to chronic iron overload in the brain of young adult rats exposed to a 150 kilohertz electromagnetic field. Neuroscience, 186: 39-47.
CrossRefPubMedDirect Link - Mahmoud, A.M., 2012. Influence of rutin on biochemical alterations in hyperammonemia in rats. Exp. Toxicol. Pathol., 64: 783-789.
CrossRefDirect Link - Silva, M., M.E. Silva, H. de Paula, C.M. Carneiro and M.L. Pedrosa, 2008. Iron overload alters glucose homeostasis, causes liver steatosis and increases serum triacylglycerols in rats. Nutr. Res., 28: 391-398.
CrossRefDirect Link - Michiels, C., M. Raes, O. Toussaint and J. Remacle, 1994. Importance of Se-glutathione peroxidase, catalase and Cu/Zn-SOD for cell survival against oxidative stress. Free Radic. Biol. Med., 17: 235-248.
PubMed - Mladenka, P., K. Macakova, T. Filipsky, L. Zatloukalova and L. Jahodar et al., 2011. In vitro analysis of iron chelating activ flavonoids. J. Inorg. Biochem., 105: 693-701.
CrossRefPubMedDirect Link - Galleano, M., M. Simontacchi and S. Puntarulo, 2004. Nitric oxide and iron: Effect of iron overload on nitric oxide production in endotoxemia. Mol. Aspects Med., 25: 141-154.
CrossRefDirect Link - Morand, C., C. Manach, V. Crespy and C. Remesy, 2000. Respective bioavailability of quercetin aglycone and its glycosides in a rat model. Biofactors, 12: 169-174.
PubMed - Morimoto, T., Y. Sunagawa, T. Kawamura, T. Takaya and H. Wada et al., 2008. The dietary compound curcumin inhibits p300 histone acetyltransferase activity and prevents heart failure in rats. J. Clin. Invest., 118: 868-878.
CrossRefPubMedDirect Link - Nahdi, A., I. Hammami, C. Brasse-Lagnel, N. Pilard, M.H. Hamdaoui, C. Beaumont and M. El May, 2010. Influence of garlic or its main active component diallyl disulfide on iron bioavailability and toxicity. Nutr. Res., 30: 85-95.
CrossRefDirect Link - Goodman, D.S., S.B. Hulley, L.T. Clark, C.E. Davis and V. Fuster et al., 1988. Report of the national cholesterol education program expert panel on detection, evaluation and treatment of high blood cholesterol in adults. Arch. Internal Med., 148: 36-69.
CrossRefDirect Link - Negre-Salvayre, A. and R. Salvayre, 1992. Quercetin prevents the cytotoxicity of oxidized LDL on lymphoid cell lines. Free Radical Biol. Med., 12: 101-106.
CrossRefPubMedDirect Link - Olynyk, J., P. Hall, W. Reed, P. Williams, R. Kerr and M. Mackinnon, 1995. A long-term study of the interaction between iron and alcohol in an animal model of iron overload. J. Hepatol., 22: 671-676.
PubMedDirect Link - Omololu, P.A., J.B.T. Rocha and I.J. Kade, 2011. Attachment of rhamnosyl glucoside on quercetin confers potent iron-chelating ability on its antioxidant properties. Exp. Toxicol. Pathol., 63: 249-255.
CrossRefPubMedDirect Link - Ostrakhovitch, E.A., L.G. Korkina, N.E. Abramova and I.B. Afanas'ev, 1995. Effect of rutin on oxygen radical production in normal and iron-overloaded rats. Toxicol. Lett., 1178: 1-88.
Direct Link - Papanastasiou, D.A., D.V. Vayenas, A. Vassilopoulos and M. Repanti, 2000. Concentration of iron and distribution of iron and transferrin after experimental iron overload in rat tissues in vivo: Study of the liver, the spleen, the central nervous system and other organs. Pathol. Res. Pract., 196: 47-54.
CrossRefPubMedDirect Link - Park, J.H. and E. Park, 2011. Influence of iron-overload on DNA damage and its repair in human leukocytes in vitro. Mutat. Res. Genet. Toxicol. Environ. Mutagenesis, 718: 56-61.
CrossRefPubMedDirect Link - Park, S.Y., S.H. Bok, S.M. Jeon, Y.B. Park, S.J. Lee, T.S. Jeong and M.S. Choi, 2002. Effect of rutin and tannic acid supplements on cholesterol metabolism in rats. Nutr. Res., 22: 283-295.
CrossRefDirect Link - Parker, M.M., F.L. Humoller and D.J. Mahler, 1967. Determination of copper and zinc in biological material. Clin. Chem., 13: 40-48.
Direct Link - Perron, N.R. and J.L. Brumaghim, 2009. A review of the antioxidant mechanisms of polyphenol compounds related to iron binding. Cell Biochem. Biophys., 53: 75-100.
CrossRefPubMedDirect Link - Pietrangelo, A., 2003. Iron-induced oxidant stress in alcoholic liver fibrogenesis. Alcohol, 30: 121-129.
CrossRefPubMedDirect Link - Pigeolet, E., P. Corbisier, A. Houbion, D. Lambert and C. Michiels et al., 1990. Glutathione peroxidase, superoxide dismutase and catalase inactivation by peroxides and oxygen derived free radicals. Mechanisms Ageing Dev., 51: 283-297.
CrossRefPubMedDirect Link - Poli, G., G. Leonarduzzi, F. Biasi and E. Chiarpotto, 2004. Oxidative stress and cell signalling. Curr. Med. Chem., 11: 1163-1182.
CrossRefPubMedDirect Link - Reddy, A.C.P. and B.R. Lokesh, 1996. Effect of curcumin and eugenol on iron-induced hepatic toxicity in rats. Toxicology, 107: 39-45.
CrossRefDirect Link - Radi, R., G. Peluffo, M.N. Alvarez, M. Naviliat and A. Cayota, 2001. Unraveling peroxynitrite formation in biolog systems. Free Radic. Biol. Med., 30: 463-488.
CrossRefPubMedDirect Link - Schafer, F.Q. and G.R. Buettner, 2001. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic. Biol. Med., 30: 1191-1212.
CrossRefDirect Link - Sestili, P., A. Guidarelli, M. Dacha and O. Cantoni, 1998. Quercetin prevents DNA single strand breakage and cytotoxicity caused by tert-butylhydroperoxide: Free radical scavenging versus iron chelating mechanism. Free Radic. Biol. Med., 25: 196-200.
CrossRefDirect Link - Shen, S.C., W.R. Lee, H.Y. Lin, H.C. Huang, C.H. Ko, L.L. Yang and Y.C. Chen, 2002. In vitro and in vivo inhibitory activities of rutin, wogonin and quercetin on lipopolysaccharide-induced nitric oxide and prostaglandin E(2) production. Eur. J. Pharmacol., 446: 187-194.
CrossRefPubMedDirect Link - Siah, C.W., D. Trinder and J.K. Olynyk, 2005. Iron overload. Clin. Chim. Acta, 358: 24-36.
CrossRefPubMedDirect Link - Sinha, A.K., 1972. Colorimetric assay of catalase. Anal. Biochem., 47: 389-394.
CrossRefPubMedDirect Link - Spirin, A.S., 1958. Spectrophotometric determination of total nucleic acids. Biokhimiia, 23: 656-662.
PubMed - Tapryal, N., C. Mukhopadhyay, M.K. Mishra, D. Das, S. Biswas and C.K. Mukhopadhyay, 2010. Glutathione synthesis inhibitor butathione sulfoximine regulates ceruloplasmin by dual but opposite mechanism: Implication in hepatic iron overload. Free Radical Biol. Med., 48: 1492-1500.
CrossRef - Tenenbein, M., 2001. Hepatotoxicity in acute iron poisoning. J. Toxicol. Clin. Toxicol., 39: 721-726.
PubMed - Theurl, I., S. Ludwiczek, P. Eller, M. Seifert, E. Artner, P. Brunner and G. Weiss, 2005. Pathways for the regulation of body iron homeostasis in response to experimental iron overload. J. Hepatol., 43: 711-719.
CrossRefPubMedDirect Link - Turbino-Ribeiro, S.M.L., M.E. Silva, D.A. Chianca Jr., H. de Paula, L.M. Cardoso, E. Colombari and M.L. Pedrosa, 2003. Iron overload in hypercholesterolemic rats affects iron homeostasis and serum lipids but not blood pressure. J. Nutr., 133: 15-20.
Direct Link - Undeger, U., S. Aydin, A.A. Basaran and N. Basaran, 2004. The modulating effects of quercetin and rutin on the mitomycin C induced DNA damage. Toxicol. Lett., 151: 143-149.
CrossRefPubMedDirect Link - Vadrot, N., A. Legrand, E. Nello, A.F. Bringuier, R. Guillot and G. Feldmann, 2006. Inducible Nitric Oxide Synthase (iNOS) activity could be responsible for resistance or sensitivity to IFN-γ-induced apoptosis in several human hepatoma cell lines. J. Interferon Cytokine Res., 26: 901-913.
CrossRefPubMedDirect Link - Valerio Jr., L.G. and D.R. Petersen, 1998. Formation of liver microsomal MDA-protein adducts in mice with chronic dietary iron overload. Toxicol. Lett., 98: 31-39.
CrossRefDirect Link - Valko, M., C.J. Rhodes, J. Moncol, M. Izakovic and M. Mazur, 2006. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact., 160: 1-40.
CrossRefPubMedDirect Link - Vasquez-Martinez, Y., R.V. Ohri, V. Kenyon, T.R. Holman and S. Sepulveda-Boza, 2007. Structure-activity relationship studies of flavonoids as potent inhibitors of human platelet 12-hLO, reticulocyte 15-hLO-1 and prostate epithelial 15-hLO-2. Bioorganic Med. Chem., 15: 7408-7425.
CrossRefDirect Link - Yang, J., J. Guo and J. Yuan, 2008. In vitro antioxidant properties of rutin. LWT-Food Sci. Technol., 41: 1060-1066.
CrossRefDirect Link - Youdim, M.B., M. Fridkin and H. Zheng, 2005. Bifunctional drug derivatives of MAO-B inhibitor rasagiline and iron chelator VK-28 as a more effective approach to treatment of brain ageing and ageing neurodegenerative diseases. Mech. Ageing Dev., 126: 317-326.
CrossRefDirect Link - Young, I.S., T.G. Trouton, J.J. Torney, D. McMaster, M.E. Callender and E.R. Trimble, 1994. Antioxidant status and lipid peroxidation in hereditary haemochromatosis. Free Radic. Biol. Med., 16: 393-397.
CrossRefDirect Link - Zhang, Y., H. Li, Y. Zhao and Z. Gao, 2006. Dietary supplementation of baicalin and quercetin attenuates iron overload induced mouse liver injury. Eur. J. Pharmacol., 535: 263-269.
CrossRefDirect Link - Zhao, Y., H. Li, Z. Gao and H. Xu, 2005. Effects of dietary baicalin supplementation on iron overload-induced mouse liver oxidative injury. Eur. J. Pharmacol., 509: 195-200.
CrossRef