
Manouchehr Safari
Department of Anatomy and Neural Stem, Cell Research Center, Semnan University of Medical Sciences, Semnan, Iran
Laya Ghahari
Department of Anatomy, Iran University of Medical Sciences and AJA University of Medical Sciences, Tehran, Iran
M.D. Babak Hossein Zadeh Zoroufchi
Department of Anesthesiology, Semnan University of Medical Sciences, Semnan, Iran
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 7 | Page No.: 931-936







ABSTRACT
Some growth factors, such as Epidermal Growth Factor (EGF), Growth Hormone (GH) and Platelet Derived Growth Factor (PDGF) have beneficial effects on keratinocyte proliferation and wound healing. Although the mechanism of these factors is unclear. In response to injury, growth factors are secreted by kinds of cutaneous cells. The goal of this project is to investigate the factors that could cause proliferate of the keratinocyte cells in vitro. The keratinocytes were removed from rat pups (10 days). Cultured in media with different concentration of GH, PDGF and EGF separately. The proliferation of cells was evaluated by the method of MTT and 3H-thymidine incorporation. Proliferation of keratinocytes was significantly higher in experimental groups than in control group. EGF maximally stimulated at 10 and 25 ng mL-1. PDGF-BB maximally stimulated at 50 ng mL-1, respectively. And maximal stimulation of GH was 2.5 IU L-1. GH, PDGF-BB and EGF stimulate keratinocyte cells proliferation in different concentration. These growth factors could play in healing of the skin.
PDF Abstract XML References Citation
Received: November 10, 2013;
Accepted: January 23, 2014;
Published: March 29, 2014
How to cite this article
Manouchehr Safari, Laya Ghahari and M.D. Babak Hossein Zadeh Zoroufchi, 2014. Effects of Epidermal Growth Factor, Platelet Derived Growth Factor and Growth Hormone
on Cultured Rat Keratinocytes Cells in vitro. Pakistan Journal of Biological Sciences, 17: 931-936.
DOI: 10.3923/pjbs.2014.931.936
URL: https://scialert.net/abstract/?doi=pjbs.2014.931.936
DOI: 10.3923/pjbs.2014.931.936
URL: https://scialert.net/abstract/?doi=pjbs.2014.931.936
INTRODUCTION
The outermost layer of skin is epidermis and contacts the outside environment directly. Rapid and efficient wound healing is essential for normal life. Wound repair is a multiple step process, included cell proliferation, differentiation, formation of granulation tissue and synthesis of various collagen. In wound healing keratinocyte migration causes re-epithelization of the skin (Lal et al., 2000). Several growth factors are essential for skin repair. Therefore, much research's have been done on these factors.
Growth Hormone (GH) widely used in children. There are serious effects, that this hormone has great effects on keratinocyte repair (Rasmussen et al., 1994; Jorgensen and Oxlund, 1996). Several studies reported that some factors such as insulin-like growth factor (IGF1) intercede the action of GH in wound repair (Pell and Bates, 1992).
These studies showed that the Epidermal Growth Factor (EGF), Platelet-Derived Growth Factor-BB (PDGF-BB) naturally produced peptides that important for preserving of skin integrity. EGF stimulates proliferation of human epithelium (Imanishi et al., 2000). Other researchers have shown than EGF have several therapeutic effects, including promoting vascularization, increasing of cell turnover and healing of epithelial and endothelial lesions (Knorr et al., 1997). EGF was found to increase epidermal thickness and stimulate proliferation of epidermal keratinocytes (Jost et al., 2000).
Locally release of PDGF promotes proliferation, vascularization and migration of connective tissue cells and then synthesis of extracellular matrix components. Cells of keratinocyte, endothelial, fibroblast and smooth muscle, contribute to PDGF generation in lesion site (Rollman et al., 2003). Mesenchymal cells in the skin express PDGF protein (Heldin and Westermark, 1999). Epidermal cells under physiological condition don’t express transmembrane receptors that recognize any type of classical PDGF isoform (Yan et al., 2007). Animal data have shown that delayed in skin repair is associated with reduce expression of PDGF protein and their receptors (Vavken et al., 2010). Human studies in diabetic ulcer have shown that topical performance of PDGF-BB promotes granulation tissue and increase vascularization (Wieman et al., 1998). PDGF is a principle mitogen for cell culture with Mesenchymal origin such as fibroblast, smooth muscle (Shin et al., 2012). The biological activity of PDGF is mediated via binding to cell-surface receptors. Two receptors for PDGF have been identified alpha and beta (Lederle et al., 2010).
Due to several activities which stimulated by the PDGF on cells, it has been implicated as a main components involved in dermal wound repair (Denk and Knorr, 2002). PDGF is a chemotactant factor for monocytes, neutrophils and fibroblast cells (Xian et al., 1997). It stimulates production of extra cellular matrix by fibroblast (Marikovsky et al., 1996). Studies have shown that PDGF-BB treatment accelerate the rate of ulcer healing in human and rats (Wong et al., 2012). All of these growth factors can stimulate proliferation of keratinocytes in vitro but the exact dose of these factors has not been identified. In this study it was assumed that adding of these factors with determine doses cause rapid proliferation of keratinocytes cells in vitro.
MATERIALS AND METHODS
Pups keratinocyte isolation and culture: All experiments in this study, especially separation and isolation of keratinocytes were accomplished according to the manifest of Helsinki. Ethical approval was taken from Semnan University of Medical Science. The cells of keratinocyte were isolated and cultured according to the protocol that described by Bevan et al. (1997). Briefly, Normal keratinocytes cells were obtained from dorsum of rat pups (10 days). They washed with warm Hanks balanced salt solution (HBSS) [sigma] containing 1.25 μg mL-1 of amphotericin. Samples were sliced into small pieces and washed again, then incubated at 37°C in 0.25% trypsin/EDTA Co2 5% for 1 h. The epidermis was removed from dermis. Resultant epidermis re-suspended in HBSS for 10 min. The remaining cells were centrifuged for 10 min at 1200 rpm and resuspended in culture medium (Dulbecco's modified Eagle’s medium [DMEM]/F12,3:1) [sigma] containing penicillin (100 IU L-1), streptomycin (100 μg mL-1), fetal bovine serum10%, hydrocortisone(0.525 μg mL-1), cholera toxin (10-10 M). The cell suspension (5x105 cells) was in 75 cm2 culture flasks. Culture flasks were sustained at 37°C containing 5% CO2. Culture medium was replaced every 3 days.
Cell proliferation (3H-thymidine incorporation) assay: Keratinocyte cells were treated with trypsin /EDTA 0/1% at 37°C for 10 min. The reaction was then stopped by adding Fetal Bovine Serum (FBS) [sigma]. The cells were counted by trypan blue exclusion and then re-plated into 24 well plates at a concentration (1x105) with respected medium. The plates were placed in incubator at 37°C overnight. The next day, culture medium was replaced with serum free medium. On the third day, 0.5 mL of serum free medium, 1 μCi 3H-thymidin, and respective growth factor with different concentration (PDGF 0, 1, 5, 10, 25, 50, 75 ng mL-1) (EGF: 0, 1, 2, 5, 10, 25, 50 ng mL-1) (GH: 0, 0.5, 2.5, 5, 10, 50 IU L-1) were added to each well of plate. The plates were placed in incubator for 24 h. On the fourth day, cells of each well were rinsed with HBSS, and then washed by 5% trichloroacetic acid. The cells of each well were dissolved with 5 mL well-1 of 1 N NaOH. Added 0.1 mL acid glacial acetic to each well. A sample aliquot of 0.1 mL was moved into scintillation vials in duplicate and counted in the scintillation counter.
Keratinocyte proliferation and effects of growth factors: Proliferation of keratinocyte cells was measured by tetrazolium bromide based colorimetric assay (MTT cell assay). All subcultures were done into the 96 well plate with 3x103 cell well-1. Above factors were added according to the protocolPDGF 0, 1, 5, 10, 25, 50, 75 (ng mL-1), EGF: 0, 1, 2, 5, 10, 25, 50 (ng mL-1), GH: 0, 0.5, 2.5, 5, 10, 50 (IU L-1). The final volume of media in each well was 100 μL. The cells were incubated for3 days in 37°C, 5% CO2 incubator. On day 3, 20 μL from MTT solution with a concentration of 5 mg mL-1 was added to the media of each well, and then was incubated for 4 h in a 37°C, 5% CO2 incubator. Culture medium was removed and then added 200 μL of dimethylsulphoxide (DMSO) to wells and stirred for 10 min in a shaker. Cell increasing was evaluated from the absorbance of 540-560 nm, using an Enzyme-Linked Immunosorbent Assay (ELISA) readerstat fox.
Statistical analysis: The results of this study are expressed as the Mean±standard error. Difference between proliferation of the cell responses following treatment with PDGF-BB, EGF and GH were compared together. Data were analysed using the one way ANOVA, and post hoc comparisons were done using Tukey’s test at 0.05 level of significance (SPSS ver.11).
RESULTS
Proliferation assay
Keratinocytes cells: Various concentrations of PDGF-BB, VEGF and GH on proliferation of keratinocytes were studied. PDGF-BB has a stimulatory effect on keratinocyte cells.
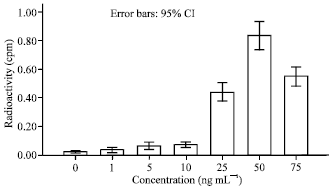 | |
| Fig. 1: | Keratinocytes cell response to PDGF. Concentration 25-50 and 75 ng mL-1 significantly increased cell number. No significant effect were observed in the concentration 1-10 ng mL-1. maximum proliferation was at 50 ng mL-1 concentration |
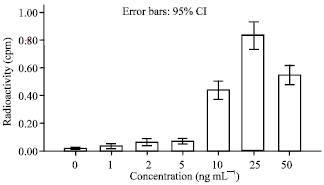 | |
| Fig. 2: | Keratinocytes cell response to EGF. Concentration 10-25 and 50 ng mL-1 significantly increased cell number. No significant effect were observed in the concentration 1-2-5 ng mL-1 maximum proliferation was at 25 ng mL-1 concentration |
Concentrations of 25-50 and 75 ng mL-1 had the most stimulatory effect for cell proliferation (p<0.05). Cell proliferation was dose dependent. The maximum proliferation was at 50 ng mL-1. There were no any significant differences between concentrations of 1-5 and 10 ng mL-1 (Fig. 1) Compared to control group, concentrations of 10-25 and 50 ng mL-1 of EGF caused a significant increase in cells proliferation. Cell proliferation was in a dose dependent manner. The maximal proliferation was at 25 ng mL-1. There was no any significant difference between concentrations 1-2 and 5 ng mL-1 (Fig. 2). The concentrations of 2.5-5 and 10 IU L-1 of GH significantly increase cell proliferation (p<0.05) (Fig. 3). The maximum stimulation was at 2.5 IU L-1 and there was no significant difference between concentrations 0/5 and 50 IU L-1. All groups were compared in Fig. 4.
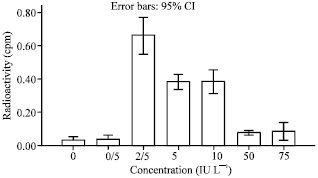 | |
| Fig. 3: | Keratinocytes cell response to GH. Concentration 2.5-5 and 10 IU L-1 significantly increased cell number. No significant effect were observed in the concentration 0/5-50 and 75 IU L-1 maximum proliferation was at 2.5 IU L-1 concentration |
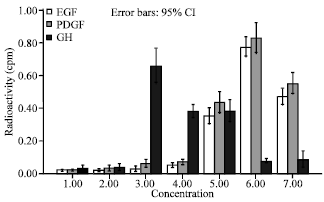 | |
| Fig. 4: | Keratinocytes cell response to GH, PDGF and EGF were compared to each other. Concentration 50 ng mL-1 PDGF have maximum effect on keratinocytes cell proliferation |
DISCUSSION AND CONCLUSION
The present study has confirmed that the proliferation of the keratinocyte cells can be increased or decreased, depending on the concentration and kind of growth factors. In this study some concentrations of EGF, PDGF-BB and GH significantly increased the proliferation of keratinocytes.
The proliferative effects of growth factors are well known, for example fibroblast proliferation increased by PDGF and FGF (Lee et al., 2010; Sun et al., 2004). Study of Wu et al. (2010) demonstrated that growth factors enhance chemo taxis at proliferative concentrations. Some growth factors such as, PDGF-BB, EGF, GH to be very important peptides in the repair and maintenance of the in many species (Haber et al., 2003). EGF concentrations 10-25 and 50 ng mL-1 have a positive effect while other concentrations such as 1-2 and 5 ng mL-1 have no stimulatory effect. The results were similar to results from other researchers and other species. In some species such as rabbit model, EGF at 1 ng mL-1 increase keratinocyte cells proliferation (Hongo et al., 1992). After stimulation with 50 ng mL-1, human keratinocytes increased by 366% (Pancholi et al., 1998). In this study, rat keratinocytes number increased after stimulation with 25 ng mL-1 of EGF. These results imply that the factor EGF in cultured human keratinocytes is useful and can be used in clinical and may have beneficial effects in clinic. Other study has shown an increase in healing when using 50 μg mL-1 of EGF (Burling et al., 2000). Administration 5 μg mL-1 has neither inflammation nor improvement of wound healing. results have shown keratinocytes cells are stimulated at the dose between 10-25 and 50 ng mL-1 but maximum stimulation was 25 ng mL-1. So using these doses within drugs can have beneficial effects on repair of lesion site on skin. There are two types of receptors on the epithelial and endothelial cells (High and low affinity). However, other researchers have shown to have low affinity receptors that are found in the keratinocyte cells. This result accord with the growth promotion effects in keratinocyte cells (Hongo et al., 1993).
PDGF-BB similar to EGF significantly increased keratinocytes proliferation in some concentration. But maximum stimulation of keratinocytes was at a concentration of 50 ng mL-1. This concentration was more effective than those reported results of other studies (Hoppenreijs et al., 1993). They reported that PDGF-BB at concentration of 50 to 100 ng mL-1 maximally promote human keratinocytes proliferation between (150-246%).
The effects of PDGF-BB on keratinocyte cells depending on the type, distribution, number of receptor and receptor sites. In some species such as human and bovine, expression of receptors are limited but in rabbits, β subunit are more numerous, then wound repaired is accelerated by PDGF (Denk et al., 2003). Results of the study indicated that stimulation of PDGF-BB at higher concentration would be important.
The addition of PDGF to various wound healing models accelerate wound healing including re-epithelialization and granulation issue formation (Ansel et al., 1993; Penha-Goncalves et al., 1997). Platelets and macrophages that enter into the wound have been proposed as the largest sources of PDGF in wound healing. These studies have shown that some concentrations of PDGF accelerate proliferation of keratinocyte hence addition of these doses of PDGF can help in wound healing. Also cultured keratinocytes produce high level of PDGF suggesting that this cell is sensitive to exogenous PDGF (Pastore et al., 2008).
Treatment of ulcers in diabetic mellitus of rats with growth factor PDGF-BB increase wound repair (Li et al., 2008). PDGF-BB is produced by many different kind of cells. This production is in responses to some stimuli, such as damages, another cytokines and some growth factors. This exerts were done by binding of factors to transmembrane receptors of tyrosine kinase. After binding, tyrosine was phosphorylated in PDGF receptor. Then some proteins are produced in intracellular matrix Subsequently, PDGF receptor-mediated induction of extracellular matrix metalloproteinase production and these factors are essential for wound healing (Li et al., 2008). These actions of PDGF demonstrated proliferation of keratinocyte cell in culture medium.
GH stimulates the development of multiple organs, not by cellular hypertrophy but by cell proliferation. GH acts both directly and indirectly on the specific cells. It induces the differentiation of progenitor cells and also increases the level of IGF-I which coincides with the results of this study, where there is a GH increase cell mitogenic effect. This implies that GH causes promotion of wound healing. Other research studies demonstrated that fibroblast proliferation decreases above the concentration of 5 IU L-1 and Li et al. (2008) have shown that GH has a dose dependency on fibroblasts (Reuterdahl et al., 1993; Cook et al., 1999). In the present study, it was found that keratinocyte proliferation increased between 2.5 to 10 IU L-1. However, the effective dose of GH on cell proliferation requires further study.
In conclusion, growth factors such as PDGF-BB, GH and EGF facilitate keratinocyte proliferation in vitro. Much remains to be studied in vivo. However, this study provides evidence that PDGF-BB, EGF and GH could be applied topically in clinic to accelerate wound repair and severe lesion.
REFERENCES
- Ansel, J.C., J.P. Tiesman, J.E. Olerud, J.G. Krueger and J.F. Krane et al., 1993. Human keratinocytes are a major source of cutaneous platelet-derived growth factor. J. Clin. Invest., 92: 671-678.
PubMed - Bevan, S., B. Woodward, R.L. Ng, C. Green and R. Martin, 1997. Retroviral gene transfer into porcine keratinocytes following improved methods of cultivation. Burns, 23: 525-532.
PubMed - Burling, K., M.A. Seguin, P. Marsh, K. Brinkman and J. Madigan et al., 2000. Effect of topical administration of epidermal growth factor on healing of corneal epithelial defects in horses. Am. J. Vet. Res., 61: 1150-1155.
PubMed - Cook, J.R., M.K. Mody and M.E. Fini, 1999. Failure to activate transcription factor NF-κB in corneal stromal cells (keratinocytes). Invest. Ophtalmol. Visual Sci., 40: 3122-3131.
PubMed - Denk, P.O., J. Hoppe, V. Hoppe and M. Knorr, 2003. Effect of growth factors on the activation of human Tenon's capsule fibroblasts. Curr. Eye Res., 27: 35-44.
PubMed - Denk, P.O. and M. Knorr, 2002. Differential regulation of expression of PDGF receptors on corneal epithelial cells. Ophthalmologe, 99: 15-19.
PubMed - Heldin, C.H. and B. Westermark, 1999. Mechanism of action and in vivo role of platelet-derived growth factor. Physiol. Rev., 79: 1283-1316.
PubMed - Hongo, M., M. Itoi, N. Yamaguchi and J. Imanishi, 1992. Distribution of Epidermal Growth Factor (EGF) receptors in rabbit corneal epithelial cells, keratinocytes and endothelial cells and the changes induced by transforming growth factor-β1. Exp. Eye Res., 54: 9-16.
PubMed - Hongo, M., M. Itoi, Y. Yamamura, N. Yamaguchi and J. Imanishi, 1993. Distribution of epidermal growth factor receptors in rabbit lens epithelial cells. Invest. Ophthalmol. Visual Sci., 34: 401-404.
PubMed - Hoppenreijs, V.P., E. Pels, G.F. Vrensen, P.C. Felten and W.F. Treffers, 1993. Platelet-Derived growth factor: Receptor expression in corneas and effects on corneal cells. Invest. Ophthalmol. Visual Sci., 34: 637-649.
PubMed - Jost, M., C. kari and U. Rodeck, 2000. The EGF receptor-an essential regulator of multiple epidermal functions. Eur. J. Dermatol., 10: 505-510.
PubMed - Knorr, M., M. Volker, P.O. Denk, K. Wunderlich and H.J. Thiel, 1997. Proliferative response of cultured human tenon's capsule fibroblasts to platelet-derived growth factor isoforms. Graefes Arch. Clin. Exp. Ophthalmol., 235: 667-671.
CrossRefPubMedDirect Link - Lederle, W., N. Linde, J. Heusel, J. Bzyl and E.C. Woenne et al., 2010. Platelet-derived growth factor-B normalizes micromorphology and vessel function in vascular endothelial growth factor-A-induced squamous cell carcinomas. Am. J. Pathol., 176: 981-994.
CrossRef - Li, H., X. Fu, L. Zhang, Q. Huang, Z. Wu and T. Sun, 2008. Research of PDGF-BB gel on the wound healing of diabetic rats and its pharmacodynamics. J. Surg. Res., 145: 41-48.
PubMed - Marikovsky, M., P. Vogt, E. Eriksson, J.S. Rubin, W.G. Taylor, S. Joachim and M. Klagsbrun, 1996. Wound fluid-derived heparin-binding EGF-like growth factor (HB-EGF) is synergistic with insulin-like growth factor-I for Balb/MK keratinocyte proliferation. J. Invest. Dermatol., 106: 616-621.
PubMed - Pancholi, S., A. Tullo, A. Khaliq, D. Foreman and M. Boulton, 1998. The effects of growth factors and conditioned media on the proliferation of human corneal epithelial cells and keratocytes. Graefes Arch. Clin. Exp. Ophthalmol., 236: 1-8.
PubMed - Pastore, S., F. Mascia, V. Mariani and G. Girolomoni, 2008. The epidermal growth factor receptor system in skin repair and inflammation. J. Invest. Dermatol., 128: 1365-1374.
PubMed - Pell, J.M. and P.C. Bates, 1992. Differential actions of growth hormone and insulin-like growth factor-I on tissue protein metabolism in dwarf mice. Endocrinology, 130: 1942-1950.
PubMed - Penha-Goncalves, M.N., D.E. Onions and L. Nicolson, 1997. Cloning and sequencing of equine transforming growth factor-β1 (TGF β-1) cDNA. DNA Sequence, 7: 375-378.
PubMed - Rasmussen, L.H., C. Garbarsch, D. Schuppan, D. Moe, K. Horslev-Pedersen, F. Gottrup and H. Steenfos, 1994. Influence of human growth hormone on granulation tissue formation, collagen deposition and the amino terminal propeptide of collagen type III in wound chambers in rats. Wound Rep. Reg., 2: 31-36.
CrossRefPubMed - Reuterdahl, C., C. Sundberg, K. Rubin, K. Funa and B. Gerdin, 1993. Tissue localization of β receptors for Platelet derived growth factor B chain during wound receptor in humans. J. Clin. Invest., 91: 2065-2075.
PubMed - Rollman, O., U.B. Jensen, A. Ostman, L. Blound, S.M. Gustafsdottir and T.G. Jensen, 2003. Platelet Derived Growth Factor (PDGF) responsive epidermis formed from human keratinocytes transduced with the PDGFβ receptor gene. J. Invest. Dermatol., 120: 742-749.
PubMed - Sun, X.Q., X.B. Fu, W. Chen and T.Z. Sun, 2004. Effects of fibroblast growth factor-10 on the secretions of transforming growth factor-α, platelet-derived growth factor-AB and vascular endothelial growth factor by normal adult human keratinocytes in culture. Zhongguo Wei Zhong Bing Ji Jiu Yi Xue., 16: 13-18.
PubMed - Vavken, P., F.A. Saad and M.M. Murray, 2010. Age dependence of expression of growth factor receptors in porcine ACL fibroblasts. J. Orthop. Res., 28: 1107-1112.
PubMed - Wieman, T.J., J.M. Smiell and Y. Su, 1998. Efficacy and safety of a topical gel formulation of recombinant human platelet-derived growth factor-BB (becaplermin) in patients with chronic neuropathic diabetic ulcers. A phase III randomized placebo-controlled double-blind study. Diabetic Care., 21: 822-827.
PubMed - Xian, W., M.P. Rosenberg and J. DiGiovanni, 1997. Activation of erbB2 and c-src in phorbol ester-treated mouse epidermis: Possible role in mouse skin tumor promotion. Oncogene, 14: 1435-1444.
PubMed - Yan, F., Y.N. Hui, Y.J. Li, C.M. Guo and H. Meng, 2007. Epidermal growth factor receptor in cultured human retinal pigment epithelial cells. Ophthalmologica, 221: 244-250.
PubMed