
Yulia Irnidayanti
Departement of Biology, Jakarta State of University, 13220, Indonesia
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 7 | Page No.: 876-883







ABSTRACT
One of the plastic-based materials widely used in the plastics industry in various countries is ester phthalate. This compound will be oxidized in the body into 2-methoxyethanol (2-ME). The effect of 2-ME on human health and environment depends on the number, duration and the frequency of exposure. Recently, the incidence of brain damage tends to increase. In the last decade, it has been widely reported the negative effects of chemical pollutants to the environment. The aim of this study were to know the existence of the expression of Vimentin and GFAP proteins caused by 2-ME on the histological structure of the cerebral cortex of mice fetal during the prenatal period on gestation day 14 (GD 14) and day 18 (GD 18). The 2-ME compound was injected intraperitoneally with a dose of 7.5 mmol kg-1 of body weight at GD-10. The result showed that there is a change in existence Vimentin protein in the cerebral cortex fetal of treated mice at GD 14, which is very conspicuous. Meanwhile, a change in existence of GFAP protein in cerebral cortex fetal of treated mice at GD 14, have relatively no difference from controls and no impact on histological structure changes of the cerebral corteks at GD 14. The change in existence of Vimentin protein in the cerebral cortex fetal of treated mice at GD 14 have an impact on histological structure of the cerebral cortex of mice treated at GD 18. It is believed that the impact is due to the effects of 2-methoxyethanol.
PDF Abstract XML References Citation
Received: July 07, 2013;
Accepted: January 17, 2014;
Published: February 12, 2014
How to cite this article
Yulia Irnidayanti, 2014. Existence of Vimentin and GFAP Protein Expressions as a result of 2-Methoxyethanol
Administration in Cerebral Cortex Tissue of Swiss Webster Mice (Mus musculus): An Immunohistochemical Analysis. Pakistan Journal of Biological Sciences, 17: 876-883.
DOI: 10.3923/pjbs.2014.876.883
URL: https://scialert.net/abstract/?doi=pjbs.2014.876.883
DOI: 10.3923/pjbs.2014.876.883
URL: https://scialert.net/abstract/?doi=pjbs.2014.876.883
INTRODUCTION
The compounds of 2-Methoxyethanol (2-ME) can be obtained from the oxidation of phthalate esters, which is widely used as the basic material of plastic and known as ethylene glycol monoethyl ether (EGME) (CICADS, 2002). This plastic compound is generally used for a variety of daily living activities, such as household appliances, packing materials, bottles, food containers, toys, water pipes and even used for health purpose as blood storage for transfusion. People use plastics in their life every day, these chemicals are potentially dangerous to their health. Although these chemicals may compromise people’s health, they have been used in the largest industrial ranks (ATSDR, 2001).
In the body, phthalate esters can undergo oxidation. The oxidation result of these compounds can be detected in the urine, particularly found in female urine (Koch et al., 2004; Silva et al., 2004). Because of that, therefore, the possibility of poisoning and exposure to phthalate esters, most often found in women. Pregnant women, fetus and newborn babies, are considered as the very sensitive groups to the exposure of these compounds. Direct exposure of these compounds to a pregnant woman will affect the embryonic microenvironment since the substance can be passed through the placental barrier (Mose et al., 2007).
Globally, approximately more than 18 million pounds of phthalate esters has been used every year and the exposure of these compounds’ toxicity can occur every day (Adibi et al., 2008), through the respiratory air and skin (Dugard et al., 1984). Phthalate ester is metabolized in the body and synthesized by the enzyme of alcohol dehydrogenase (ADH) and aldehyde dehydrogenase (ALDH). One of the metabolites of phthalate esters is methoxyethanol (2-ME), which will be metabolized to methoxyacetic acid (MAA) are toxic and teratogenic compounds in rats and can cause physical abnormalities, reproductive disorders and skeletal and hematopoietic disorders (Brown et al., 1984; Miller et al., 1982). The developmental disorder caused by the MAA is apparently because of the content of fatty acid chains attached to the MAA. It is suspected that this fatty acid chain causes a covalent bond modification on cell chromatin proteins (Wade et al., 2008), which in turn can inhibit histone deacetylase activity that cause changes in gene expression (Henley et al., 2009).
Previous research has reported that the compounds of 2-ME are not only toxic in rat but also in several mammalian species (Feuston et al., 1990). People have been poisoned with 2-ME through penetration into the skin and respiration tube (Dugard et al., 1984). Exposure via medical equipments occurs only in a minority of the population. Approximately 100,000 people were poisoned by 2-ME per year. Predictably, they are women who are still fertile or able to produce offspring (Scott et al., 1989).
The compound of 2-ME has been known of its potential to cause abnormalities, such as exencephaly in mice fetus (Darmanto, 1998). Dam of mice that had been given at a dose of 2-ME 12.5 mmol kg-1 b.wt. on gestation days 11, have an impact on fetal development disorder on gestation days 18. Some disorders are anencephaly, thinning of the cerebral cortex, cerebellar abnormalities in foliation pattern and also reduce levels of total protein in the brains of fetal brain (Prihiyantoro et al., 2004). The toxic effects of 2-ME is not just the result of the secondary metabolites, MAA (Methoxyacetic acid), but the compound also has a direct target on the bases of purine and pyrimidine. It can cause moieties on the carbon of the purine and pyrimidine bases (Mebus and Welsch, 1989). As a result, availability for purine and pyrimidine bases, that required for the synthesis of protein, DNA and RNA, also decreased. Inhibition on RNA or protein synthesis in certain cells can give impact on the increase of the cell death (Ellis et al., 1991; Umansky, 1996). This condition affects the proliferation and differentiation of embryonic cells which can cause structural abnormalities (Ruyani et al., 2001). Since proteins have significant role in cellular development, disorder of protein expression in early embryonic development can result in developmental disorders. These disorders occur primarily due to the imbalance between cell proliferation and cell death.
A study by Hildebrand and Soriano (1999) have shown that the treatment with 2-ME can increase cell death in neuroepithelium. The cell death occurs due to an imbalance in cell proliferation, that required for neural tube closure. It certainly can inhibit neural tube closure. Therefore, the 2-ME compound has the potential cause of the developmental abnormalities in the brain. Brain is an essential organ for all body system coordination. Therefore, the failure of brain development will result in disruption of almost every function of the body.
The formation of the brain occurs in the early stage of development, although the process may complete in the late stage of development or after the birth. The development of brain involves many processes such as cell proliferation, migration, interaction, adhesion, differentiation and morphogenesis (Gilbert, 2000). All the mechanisms are also involved in the formation of extracellular matrix proteins. In the Central Nervous System (CNS), the extracellular protein substance composed of proteins that secreted primarily by astroglial cells, which is embedded in the substance of extracellular matrix (ECM). Vimentin and Glial Fibrillary Acidic Protein (GFAP), are extracellular proteins secreted by the astroglial cells, that has a role as a substrate for the cells to migrate and signaling in the brain development (Bruce et al., 2008). The pattern of the protein expression are temporary and depends on the tissue differentiation. The disruption of extracellular proteins in the brain can cause an impaired mechanism of cell interaction by which disruption of brain development process occurs. Additionally, the interference of protein synthesis in astroglial cells can inhibit the process of neuroblast migration and proliferation which, ultimately, also cause the abnormalities in brain development.
It has been reported that migration disorders, particularly Purkinje cell migration can cause abnormal cerebral foliation patterns in mice (Darmanto, 1998, 2002). The abnormalities in migration are resulted from a decreased expression of an extracellular protein called Reelin. It is a type of extracellular proteins secreted by Cajal-Retzius cells located in the marginal zone of the cerebral cortex of the brain (Dutta et al., 2005). This protein is thought to function as a regulator of cell positioning and to direct the migration of nerve cells in the brain (Gotz et al., 1996). Apparently, Reelin is not the only protein that plays a role in the process of brain development. It has been recognized that brain development involves multiple processes and dynamic changes, therefore, a various proteins are required. Several proteins have been recognized to be involved in brain development including Fibronectin, Tenascin, Vimentin, Ncam and Neurofilament (Duprey and Paulin, 1995; Liu et al., 2004; Kolkova, 2008; Helfand et al., 2011).
Previously, researchs have reported that the mRNA levels of gene expression of extracellular Fibronectin, Tenascin, Vimentin, Neurofilament and neural cell adhesion molecules (Ncam) have been found on gestation days 10 (GD 10). Vimentin is expressed very high compared to the others (Irnidayanti et al., 2011). These data are supported by the result of previous studies. The brains of embryonic mice treatment on gestation days 12 (GD 12) have also showed a very high expression of mRNA level of Vimentin gene, but lower expression of Vimentin protein (Irnidayanti et al., 2010). After the recovery process, the cells undergo dedifferentiation or start proliferating again. Following the process, Vimentin protein has been expressed again in the cells undergoing the proliferation. It is believed that 2-ME inhibits the translation of Vimentin protein on gestation days 12 (GD 12) as a cellular response to toxic materials. These responds are characterized by higher levels of GFAP protein in the treatment group on gestation days 12 (GD 12) than in the control group and increased levels of absorption of Vimentin protein on gestation days 12 (GD 12). It means that the interference occurred at the stage of protein translation since the proteins have not been fully translated. This is evidenced by the low absorption level of Vimentin expression in the treatment fetal brain on gestation days 12 (GD 12), while the absorption level of GFAP expression in the fetal brain on GD 12 is very high. After this period, the expression of GFAP became lower again and Vimentin protein expression levels in the treatment group increase on gestation days 17 (GD 17) and on gestation days 18 (GD 18) (Irnidayanti et al., 2013).
Therefore, it is believed that extracellular proteins such as Vimentin plays an important role in brain development, particularly in the process of neurongenesis. In the early developmental stage of central nervous system, the extracellular matrix of immature astrocytes is mainly composed of Vimentin. This protein can be detected as early as GD 11 in radial fibers of the neural tube located in the ventricular zone (Oudega and Marani, 1991). Then, around birth, a switch from Vimentin to GFAP takes place. Vimentin is expressed as an extracellular protein in the early development and is used as a marker of glial cell differentiation. Vimentin plays a role in maintaining the integrity of cell structure as well as cell shape and stability. Vimentin also plays a role in replication, recombination and repair of DNA (Oudega and Marani, 1991). Vimentin disappears and progressively replaced by GFAP in differentiated astroglial cells that transiently co-express the two proteins. GFAP functions in the formation of the structural framework of the cell and the cell shape and organizes the organelles in the cytoplasm. A structural role of the protein is as the cytoskeleton, which is responsible not only for the movement inside the cell but also for the internal transport and cytoplasmic organelles. Both GFAP and Vimentin are components of intermediate filaments of astrocytes cells. Astrocytes are glial cells appear in the early development of the brain and acts to guide the growth of nerve cells.
Based on the above background, the research aimed to know the existence of expression of GFAP and Vimentin protein on the cortex cerebral layers caused by methoxyethanol on gestation day 14 (GD 14) and GD 18 by Immunohistochemistry technique.
MATERIALS AND METHODS
Experimental animals, material and sample collection: Female mice Mus musculus (Swiss Webster) used in the experiment came from Animal House and Toxicology Laboratories, University of Indonesia. Rearing the animals was done in a room of temperature at 23-27°C with 83% humidity. Food and water were given ad libitum. Ten to twelve weeks old female mice were mated with a male (1:1) overnight. A vaginal plug detected in the next morning was defined as day zero of gestation day (Rugh, 1968).
A liquid form of 2-ME (Product No.: 135-07762) produced by Wako Pure Chemical Industries, Ltd., Japan, was diluted with sterilized distilled water. A single dose of 7.5 mmol 2-ME kg-1 of body weight was administered by peritoneal injection to the mice dams on GD 10. The control dams were injected only with sterile distilled water with the same volume.
The mice dam on GD 14 and GD 18 were anaesthetized by chloroform and perfuse with 1% saline solution dan 4% Paraformaldehyde (PFA). Embryo from each dams was removed and put into a falcon tube containing buffer solution. Fetal brain was isolated and fixated in 4% PFA solution overnight.
Paraffin method: Uterus was opened, embryo was removed and the fetal brain was isolated under stereo microscope. Fetal brains were fixated in 4% PFA solution for 4 days. Further dehydration in alcohol gradually increased the concentration. Purification was done in a solution of alcohol-xilol and xilol-pure. Paraffin infiltration was done in an oven at a temperature of 57-60°C. Embedding the specimen was conducted in rectangular-metal containers containing paraffin. Brain specimen were sliced by a rotary microtome, at 5 μm thickness and then taped on the glass objects that have been poured by Mayer's albumen and water. The slides were stretched and dried in an oven at temperature of 42°C. In order to remove the paraffin, the slides were immersed in a xilol solution, hydrated in alcohol which concentration decreased gradually and then washed by distilled water. Finally, the slides were stained by a solution of Hematoxylin-Eosin (Conn et al., 1960).
Immunohistochemistry technique (manufactured protocols): All Sample Slides are dropped 3% solution of peroxide and absolute methanol for 10 min, drained and then washed with 0.01% Phosphate-buffered Saline (PBS) solution of pH 7.2 for 2 min. Blocking was done by dropping with blocking solution 2 drops for 10 min. Blocking solution was used for blocking non-specific antigens or proteins followed by dropping 2 drops of a monoclonal antibody, Vimentin and GFAP (Clone ZCG29, Lot No. 750727A). Let stand for 60 min. After that, the slide was soaked in a solution of 1 X PBS for 2 min and then drops 2 drops of secondary antibodies and let stand for 10-15 min. Drain and soak the slides in 1X PBS solution for 2 min and 2 drops of Horseradish Peroxidase (HRP)-Streptavidin for 2 min. Prepare a mixture of H2O2 solution, 3,3'-diaminobenzidine (DAB) and Buffer, are mixed each 1 drop into 1 mL aquadest, in a dark tube. Drop mixed solution on the object slide and leave it on for 3-5 min. Wash with sterile distilled water and stained with haematoxylin-Eosin. To obtain a blue color, slide immersed in 1X PBS solution after it was washed by sterile distilled water, dehydrated in increasing concentrations of alcohol. Slide was soaked in a solution of xilol, then mounting and covered. When the colors were brown, positive antigen and antibody reaction occured, that means, there are a protein Vimentin and GFAP.
RESULTS
Normally, the histology structure of mice fetal brain, particularly the cerebral cortex consists of VZ (ventricular zone), IZ (intermediate zone) and MZ (marginal zone). The gestation days 14 (GD 14) is not different from the structure of the cerebral cortex of mice fetal control on GD14. However, Immunohistochemistry (IHC) assay results showed a difference between control and treated of the cerebral cortex of mice fetal on GD 14. Result of Immunohistochemistry on the cerebral cortex of mice fetal control at GD 14 was dark brown colored (Fig. 1a). The dark brown color indicates that the existence of the protein Vimentin is relatively very much. In Fig. 1b, appears that the cerebral cortex of mice fetal treated on GD 14 was colored light brown. The light brown color indicates that the existence of the protein Vimentin is relatively fewer in number. Result of Immunohistochemistry on the cerebral cortex of mice fetal control on GD 18 was light of brown colored (Fig. 1c) and similar to the cerebral cortex of mice fetal treated on GD 18 (Fig. 1d). However structure of the cerebral cortex of mice fetal treated on GD 18 (Fig. 1d) was different from structure of the cerebral cortex of mice fetal treated on structure of the cerebral cortex of mice fetal control on GD 18.
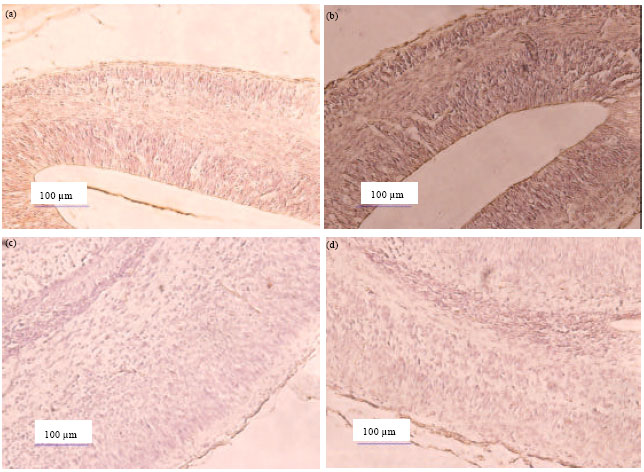 | |
| Fig. 1(a-d): | (a) Immunohistochemistry of vimentin protein in the cerebral cortex of mice fetal control at gestation days 14 (10x20), (b) In cerebral cortex of mice fetal treated at gestation days 14, (c) In cerebral cortex of mice fetal control at gestation days 18, (d) In cerebral cortex of mice fetal treated at gestation days 18 (Dark brown color indicated vimentin protein is showed a lot of protein vimentin and light brown color indicated is slight vimentin protein |
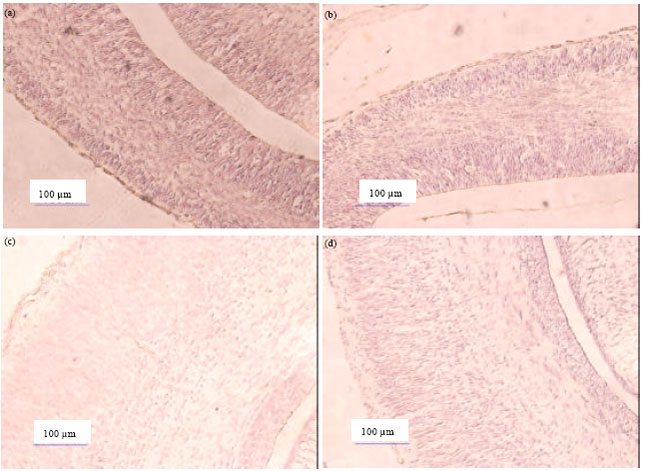 | |
| Fig. 2(a-d): | (a) Immunohistochemistry of glial fibrillary acidic protein (GFAP) in cerebral cortex of mice fetal control at gestation days 14 (10x20), (b) In cerebral cortex of mice fetal treated at gestation days 14, (c) In cerebral cortex of mice fetal control at gestation days 18, (d) In cerebral cortex of mice fetal treated at gestation days 18 (Dark brown color indicated vimentin protein is showed a lot of protein vimentin and light brown color indicated is slight vimentin protein |
It is believed that 2-methoxyethanol cause changes in the existence protein Vimentin and thus will give an impact on the changes in the histological structure of the cerebral cortex of mice fetal treated on GD 18.
The data is supported by the result of densitometry in previous studies (data not shown). Total absorption level of the Vimentin protein on brain of mice fetal control on gestation days 14 (GD 14) is 151.55. The existence of Vimentin protein on brain of mice fetal on gestation days 14 (GD 14) have change after administration of 2-ME, which is characterized by the decrease of light brown colour on the cortex cerebral. The total absorption level of the Vimentin protein on brain of mice fetal treated on gestation days 14 (GD-14) is 122.81 (Irnidayanti et al., 2013).
Along with the process of brain development and increasing age, the existence of Vimentin protein in the cerebral cortex of fetal mice is also changing. Previous studies show that total absorption level of the Vimentin protein of mice fetal treated on GD 18 was increasing from 133.00 become 163.83 (Irnidayanti et al., 2013). On the advanced development of the brain, the protein vimentin should be decreased, but after administration of 2-ME levels led to an increase in the total absorption level of the Vimentin protein. Then, around birth a switch from Vimentin to GFAP takes place.
Vimentin is expressed as extracellular protein in early development. The existence of GFAP protein has associated with the existence of protein Vimentin. In Fig. 2b shows that result of Immunohistochemistry on GFAP protein of the cerebral cortex of mice fetal treated at gestation days GD 14 (GD 14) has light of brown colored than result of Immunohistochemistry on GFAP protein in the cerebral cortex of mice fetal control on gestation days 14 (Fig. 2a). It mean that the existence of GFAP protein on cerebral cortex tends to decrease after administration 2-ME. However, the densitometry results showed an increase total absorption level of the GFAP protein on brain of mice fetal control at GD 14 from 133.50 become 163.58 on brain of mice fetal treated at GD 14. It is possible that the increased GFAP protein is not only derived from the cerebral cortex.
Result of immunohistochemistry on GFAP protein of cerebral cortex of mice fetal treated at gestation days 18 (Fig. 2d) was showed relatively no difference to controls (Fig. 2c). This data is supported by previous research that results of densitometry of GFAP protein total on mice brains fetal treated at GD-18 and GD-18 are 95.14 become 91.80 (Irnidayanti et al., 2013).
DISCUSSION
Histological structure of cerebral cortex on fetal of control mice at gestation days 14 (GD-14) and cerebral cortex on fetal of treated mice at gestation days 14 (GD-14) did not indicate a difference. However, the histological structure of the cerebral cortex on fetal of treated mice at gestation 18 (GD-18) shows the difference compared with controls. It is estimated that changes in the histological structure of cerebral cortex on fetal of treated mice at 18 days of gestation due to 2-ME compound. This compound causes the interference of Vimentin gene expression on levels of mRNA and Vimentin protein expression (Irnidayanti et al., 2010), which of course leads to the disorder of corticogenesis in the cerebral cortex and the existence of protein Vimentin in the cerebral cortex.
One stage in brain development is corticogenesis. During corticogenesis, the proliferation of neuroephitelium cells occur in the ventricular zone of the cerebral cortex and take on gestation days 11 to 17 (Takahashi et al., 1995). Therefore, administration of 2-ME on pregnant mice on gestation day 10 (GD 10), can disrupt the proliferation process of cells neuroepitelium in the cerebral cortex. The proliferation disorders may be caused by interference on expression of Vimentin protein, which is needed for the proliferation of neuroepithelium. The compound 2-ME in the body is metabolized to MAA. MAA as intermediate products can disrupt proteins of histon 1. The role proteins of histon 1 as histones compact Stabilize (HSC). Proteins of histon 1 are a protein associated with the higher chromatin structure, regulate gene expression and are involved in chromatin-based processes such as DNA replication and repair (Happel and Doenecke, 2009). Disorder on proteins of histone 1 that caused by MAA administration led to chromatin architecture becomes unstable and genetic disorders. The condition is a cellular response to toxic materials, thus causing changes in the existence of protein expression.
These data are supported by previous studies that 2-ME inhibits translation of Vimentin protein on gestation days 12 (Irnidayanti et al., 2013). Gestation days 12 (GD 12) is the starting point of recovery process. This is based on the assumption that astroglial cells are responsive to the toxic substance 2-ME, which causes tissue damage. As a response, astroglial cells are progressively transformed into reactive astrocytes. Changes in reactive astrocytes may alter gene expression and cause cellular changes (Brahmachari et al., 2006), which ultimately alter the protein translation process.
Based on the research before, the Vimentin expressed very high, but the function of this protein can be occurred maximally, if its presence together with protein GFAP, the timetable and its amount should be appropriate (Irnidayanti et al., 2013). If the recovery process at the level of cells and tissues have been occurred, but the situation can not improve the positioning of cells that have occurred in the brain tissue, this situation certainly may cause interference in all these mechanisms, thus disturbing formation of the layers of the cerebral cortex. The process of cell positioning on the cortex layers is affected by the existence and amount of GFAP and Vimentin protein in a certain degree. Nevertheless, the amount of protein expression and the existence of protein vimentin is very high, it is not enough to repair neuroephitellium cells of the cerebral cortex through the process of proliferation mechanism.
A switch of the expression of Vimentin protein by protein GFAP occur around birth, but in this study the existence of GFAP protein was detected gestation days 14 (GD 14). In this study, the existence of expressed GFAP is not enough to recover the process corticogenesis. Accuration of timetable of GFAP is extremely needed for the recovery process. After this period, the expression of GFAP became lower again and associated with the expression of Vimentin. After the recovery process, all the cells undergo redifferentiation and start proliferating again.
CONCLUSION
The results of this study conclude that 2-methoxyethanol causes the changes in the existence of Vimentin protein in the cerebral cortex of the fetal of treated mice at GD 14. 2-methoxyethanol inhibits the translation of Vimentin protein and give an impact on histological structure of the cerebral cortex of treated mice at GD 18. The reduced existence of GFAP protein in the cerebral cortex of treated mice on gestation day 14 (GD 14), believed to be a cellular response to 2-methoxyethanol.
REFERENCES
- Adibi, J.J., R.M. Whyatt, P.L. Williams, A.M. Calafat and D. Camann et al., 2008. Characterization of phthalate exposure among pregnant women assessed by repeat air and urine samples. Environ. Health Perspect, 116: 467-473.
Direct Link - Brahmachari, S., Y.K. Fung and K. Pahan, 2006. Induction of glial fibrillary acidic protein expression in astrocytes by nitric oxide. J. Neurosci., 26: 4930-4939.
CrossRef - Brown, N.A., D. Holt and M. Webb, 1984. The teratogenicity of methoxyacetic acid in the rat. Toxicol. Lett., 22: 93-100.
CrossRef - Dugard, P.H., M. Walker, S.J. Mawdsley and R.C. Scott, 1984. Absorption of some glycol ethers through human skin in vitro. Environ. Health Perspect., 57: 193-197.
Direct Link - Duprey, P. and D. Paulin, 1995. What can be learned from intermediate filament gene regulation in the mouse embryo. Int. J. Dev. Biol, 39: 443-457.
PubMed - Ellis, R.E., J. Yuan and H.R. Horvitz, 1991. Mechanisms and functions of cell death. Annul. Rev. Cell Biol., 7: 663-698.
CrossRef - Feuston, M.H., S.L. Kerstetter and P.D. Wilson, 1990. Teratogenicity of 2-methoxyethanol applied as a single dermal dose to rats. Toxicol. Sci., 15: 448-456.
CrossRef - Gotz, B., A. Scholze, A. Clement, A. Joester and K. Schutte et al., 1996. Tenascin-C contains distinct adhesive, anti-adhesive and neurite outgrowth promoting sites for neurons. J. Cell Biol., 132: 681-699.
CrossRef - Happel, N. and D. Doenecke, 2009. Histone H1 and its isoforms: Contribution to chromatin structure and function. Gene, 431: 1-12.
CrossRef - Helfand, B.T., M.G. Mendez, S.N.P. Murthy, D.K. Shumaker and B. Grin et al., 2011. Vimentin organization modulates the formation of lamellipodia. Mol. Biol. Cell, 22: 1274-1289.
CrossRef - Henley, D.V., S. Mueller and K.S. Korach, 2009. The short-chain fatty acid methoxyacetic acid disrupts endogenous estrogen receptor-alpha-mediated signaling. Environ. Health Perspect., 117: 1702-1706.
PubMed - Hildebrand, J.D. and P. Soriano, 1999. Shroom, a PDZ domain-containing actin-binding protein, is required for neural tube morphogenesis in mice. Cell, 99: 485-497.
CrossRef - Irnidayanti, Y., W. Darmanto and A. Abadi, 2010. Ekspresi level gen mRNA protein ekstraseluler otak embrio mencit black-6 UK-12 akibat induksi 2-methoxyethanol: Analisis secara real time RT-PCR [Expression of level gen mRNA of extracellular protein embrio brain mice black-6 at GD-12 caused by induced 2-methoxy-ethanol: Analysis by real time RT-PCR]. Berkala Penelitian Hayati UNAIR, 15: 171-179, (In Indonesian).
Direct Link - Irnidayanti, Y., W. Darmanto, A. Abadi, Y. Hattori and Y. Yamashiro, 2013. Differential expression of vimentin and GFAP protein during brain development of mouse fetuses after treated with 2-Methoxyethanol. ITB J. Sci., 44: 346-357.
CrossRefDirect Link - Koch, H.M., H.M. Bolt and J. Angerer, 2004. Di(2-ethylhexyl)phthalate (DEHP) metabolites in human urine and serum after a single oral dose of deuterium-labelled DEHP. Arch. Toxicol., 78: 123-130.
CrossRef - Liu, Q., F. Xie, S.L. Siedlak, A. Nunomura and K. Honda et al., 2004. Neurofilament proteins in neurodegenerative diseases. Cell. Mol. Life Sci., 61: 3057-3075.
CrossRef - Mebus, C.A. and F. Welsch, 1989. The possible role of one-carbon moieties in 2-methoxyethanol and 2-methoxyacetic acid-induced developmental toxicity. Toxicol. Applied Pharmacol., 99: 98-109.
PubMedDirect Link - Miller, R.R., R.E. Carreon, J.T. Young and M.J. McKenna, 1982. Toxicity of methoxyacetic acid in rats. Fundam Applied Toxicol., 2: 158-260.
PubMedDirect Link - Mose, T., G.K. Mortensen, M. Hedegaard and L.E. Knudsen, 2007. Phthalate monoesters in perfusate from a dual placenta perfusion system, the placenta tissue and umbilical cord blood. Reprod. Toxicol., 23: 83-91.
CrossRefDirect Link - Oudega, M. and E. Marani, 1991. Expression of vimentin and glial fibrillary acidic protein in the developing rat spinal cord: An immunocytochemical study of the spinal cord glial system. J. Anatomy, 179: 97-114.
PubMed - Silva, M.J., J.A. Reidy, A.R. Herbert, J.L. Preau Jr, L.L. Needham and A.M. Calafat, 2004. Detection of phthalate metabolites in human amniotic fluid. Bull. Environ. Contam. Toxicol., 72: 1226-1231.
PubMed - Scott, W.J., R. Fradkin, W. Wittfoht and H. Nau, 1989. Teratologic potential of 2-methoxyethanol and transplacental distribution of its metabolite, 2-methoxyacetic acid, in non-human primates. Teratology, 39: 363-373.
CrossRef - Takahashi, T., R.S. Nowakowski and V.S. Caviness Jr., 1995. The cell cycle of the pseudostratified ventricular epithelium of the embryonic murine cerebral wall. J. Neurosci., 15: 6046-6057.
Direct Link - Wade, M.G., A. Kawata, A. Williams and C. Yauk, 2008. Methoxyacetic acid-induced spermatocyte death is associated with histone hyperacetylation in rats. Biol. Reprod., 78: 822-831.
CrossRefPubMedDirect Link