
Narayan Mohapatra
Avian Diseases Section, Division of Pathology, Indian Veterinary Research Institute, Izatnagar, 243122, Bareilly (UP), India
Jag Mohan Kataria
Avian Diseases Section, Division of Pathology, Indian Veterinary Research Institute, Izatnagar, 243122, Bareilly (UP), India
Sandip Chakraborty
Department of Animal Resources Development, Pt. Nehru Complex, Agartala, Pin, 799006, India
Kuldeep Dhama
Avian Diseases Section, Division of Pathology, Indian Veterinary Research Institute, Izatnagar, 243122, Bareilly (UP), India
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 6 | Page No.: 821-828







ABSTRACT
Egg Drop Syndrome-76 (EDS-76) is a recognized disease of chickens and Japanese Quails, which is of high economic importance due to its drastic negative effects on egg production in laying birds. The aim of the present study was to better understand the EDS-76 viral disease process in Japanese quails (Coturnix coturnix japonica), since very limited studies have been conducted in this species of birds. For this purpose, an experimental study was conducted with infection of EDS-76 virus in laying Japanese quails to reveal pathology, effect on egg production/quality and immune responses of this virus in these birds. By 7, 9 and 13-15 Days Post Infection (DPI), drop as well as aberrant egg production and lower mean egg quality were observed compared to control birds. Significant histopathological changes were observed in genitalia and spleen. Haemagglutination Inhibition (HI) and Enzyme Linked Immunosorbant Assay (ELISA) titres rose rapidly by 2nd week when it became maximum; thereafter declined and maintained at low levels up to 10 week post infection. The mean total protein values in infected quail gradually increased to 4.10±0.05/100 mL without any change in mean albumen value at 12 DPI. In conclusion, the course of the EDS-76 is significant not only in chickens but also in quails even though it occurs occasionally in quails. Explorative pathological, blood biochemical and immunological studies are suggested during EDS-76 viral disease course in quails. This would aid in formulating effective disease prevention and control measures for this economically important disease of poultry.
PDF Abstract XML References Citation
Received: June 28, 2013;
Accepted: August 26, 2013;
Published: January 29, 2014
How to cite this article
Narayan Mohapatra, Jag Mohan Kataria, Sandip Chakraborty and Kuldeep Dhama, 2014. Egg Drop Syndrome-76 (EDS-76) in Japanese Quails (Coturnix coturnix japonica): An Experimental Study Revealing Pathology, Effect on Egg Production/Quality and Immune Responses. Pakistan Journal of Biological Sciences, 17: 821-828.
DOI: 10.3923/pjbs.2014.821.828
URL: https://scialert.net/abstract/?doi=pjbs.2014.821.828
DOI: 10.3923/pjbs.2014.821.828
URL: https://scialert.net/abstract/?doi=pjbs.2014.821.828
INTRODUCTION
Egg drop syndrome (EDS-76), reported by Van Eck et al. (1976) and McFerran et al. (1978), is a vertically transmitted and an economically important disease of laying hens (Kumar et al., 2003; Kataria et al., 2005; Ezeibe et al., 2008; Dhama et al., 2011). Fragility of eggs together with decreased rate of fertility and hatchability are the outcomes. The EDS-76 virus (EDSV) has been classified under group 3 of genus Aviadenoviridae. A large variety of wild as well as domestic vertebrate animals are infected by medium-sized Adenoviruses that are non-enveloped DNA viruses. In the mid 1970’s Egg Drop Syndrome (EDS) emerged throughout the world (Baxendale, 1978; Swain et al., 1997; McFerran and Smyth, 2000; Kumar et al., 2003). Serological evidence of EDS-76 virus infections in India has been found in chickens and quail (Kataria et al., 1991; Das and Pradhan, 1992; Das et al., 1995; Shaw et al., 1995) and in some instances the causative EDS-76 virus has been isolated and characterized biochemically and biologically (Swain et al., 1993; Chandramohan, 1996; McFerran and Adair, 2003; Woolcock, 2008). Many reports have been published regarding the etiology and pathogenesis of experimentally reproduced EDS-76 viz., (Taniguchi et al., 1981; Yamaguchi et al., 1981a; Van Eck 1986; Kumar, 1990; Swain et al., 1993, Chandramohan, 1996) and systematic study of this condition in chicken has been done. The disease occurs naturally in turkeys but the disease in ducks is more severe (Bidin et al., 2007; Su et al., 2011). The natural occurrence of this condition in Japanese quails (Coturnix coturnix japonica) has also been reported by Dash and Pradhan (1990) and Kataria et al. (1991) and a virus serologically indistinguishable from that of the chicken has been isolated for the first time. However, no systematic study of this disease condition in Japanese quail has been done (McFerran and Adair, 2003). In this study, results of experimental studies carried out in Japanese quails following infection with EDS-76 virus of quail origin has been reported with particular reference to pathological findings, changes in blood biochemical parameters during the course of the disease and immunological studies, which would help in better understanding of EDS-76 viral disease process in quails and prevent and control the disease in a better way.
MATERIALS AND METHODS
Birds: One hundred and thirty five, 4 weeks old quail pullets seronegative to EDS-76 virus were obtained from Experimental Quail Farm of Central Avian Research Institute (CARI), Izatnagar. The birds were maintained in the Experimental Shed of the Division of Avian Diseases, Indian Veterinary Research Institute (IVRI) Izatnagar. The birds started laying at 6 weeks and at about 8 weeks the production increased up to 50%. Before start of the experiment the birds were serologically screened for Ranikhet Disease (RD), avian Infectious Bronchitis (IB), Chicken Embryo Lethal Orphan (CELO) virus, Salmonella and Mycoplasma and found negative to these avian pathogens.
Virus, antisera and conjugate: Virus and antisera of EDS-76 virus of quail origin (QEDS-76-1/AD-1/91) was obtained from the Virus Laboratory of the Division of Avian Diseases, Indian Veterinary Research Institute, Izatnagar in the form of freeze dried allantoic fluid. The virus was inoculated in 12-14 days old embryonated duck eggs for reactivation before the experiment. The standard serum against EDS-76 virus and its FITC conjugated globulin were also obtained from the above laboratory.
Experimental design: A total of 80 birds were inoculated with EDS-76 virus (109.5 EID50/mL/2 birds) by naso-oral route and rest 55 birds were treated as healthy. The infected and healthy birds were divided into 3 separate groups, viz., Group 1: For clinico-pathological, immunological, contact transmission and excretion studies-50 infected and 10 healthy birds were kept together, Group 2: For egg production study-30 infected birds were kept in colony cages and rest 30 healthy birds were kept in a separate cage and for control Group 3:15 healthy birds were kept as control in a separate shed. All the three groups of birds were reared under similar managemental conditions.
Egg production and egg quality studies: The eggs were collected from infected and healthy birds of group 2 up to 21 days. The egg production rate was noted. Randomly 10 eggs from control and 10 from infected birds were taken for study of egg quality viz., shell texture, shell colour, shell thickness, egg weight and Internal Quality Unit (IQU) determination. The IQU was used as a measure of egg quality in relation to egg weight and was calculated as per the formula of Kondaiah et al. (1981):
IQU = 100 Log (H + 4.18-0.898W 0.6674)
where, H is albumen height and W is weight of the egg in grams.
The statistical analysis of all the values was carried out by using Student’s ‘T’ test (Snedecor and Cochran, 1967).
Virus localization and histopathology: Three quails from the group-1 and one quail from group 3 were sacrificed at 3, 6, 9, 12, 15, 18, 21, 28, 35, 42 and 49 Days Post Infection (DPI) each. Serum was collected from each bird and tested for antibodies against EDS-76 virus by Haemagglutination Inhibition (HI) method (Swain et al., 1993). From the sacrificed birds pieces of uterus, vagina, spleen and ovary were collected in 10% formalin for histopathological examination and pieces of uterus, spleen, vagina and nasopharynx were also kept at -20°C for detection of viral antigen by Fluorescence Antibody Test (FAT). At 3, 4, 5 and 6 DPI, the buffy coat was separated from the blood of the infected birds and stained with anti-EDS-76 virus FITC conjugated globulin to demonstrate the presence of viral antigen in the circulating white blood cells as per the method of Van Eck (1986).
Virus excretion and contact transmission: The virus excretion study was performed by isolation of the EDS-76 virus from cloacal swabs collected at every alternate day from infected and healthy birds up to 3 weeks post infection (WPI). Contact transmission study was done on the basis of seroconversion in contact healthy birds. The cloacal swabs were processed for virus isolation in Chicken Embryo Liver (CEL) cell culture which was prepared by using method of Adair et al. (1979).
Biochemical study: The serum samples collected from the sacrificed birds at 3, 6, 9, 12, 15, 18 and 21, DPI were used for biochemical studies. The total serum protein, albumen, calcium and inorganic phosphorus contents were estimated by following directions mentioned in standard diagnostic kits (supplied by Span Diagnostics, Udharia, Surat, Gujrat, India). The serum globulin concentration was determined by subtracting the albumen quantity from total protein value. All the values were subjected to statistical analysis using Student’s ‘T’ test.
Immunological studies: Detection of humoral immune response in infected, in-contact and healthy birds against EDS-76 virus was done using haemagglutination (HI) test, Agar Gel Precipitation Test (AGPT) (Swain, 1991) and Enzyme Linked Immunosorbent Essay (ELISA) (Adair et al., 1986). At weekly intervals, 10 birds from infected and 10 birds from control group were bled and tested for serum antibodies against EDS-76 virus.
RESULTS AND DISCUSSION
Clinical sign and egg production study: Japanese quail layer birds, following artificial infection of EDS-76 virus via oral route, experienced drop in egg production with production of thin shelled eggs which was exhibited as the only clinical sign. The drop in egg production in infected quails was from 70-63% at 7 Days Post Infection (DPI). The lowest egg production was of 40% at 13 DPI. The birds remained apparently healthy during the entire observation period. The egg production increased gradually thereafter and at 21 DPI became 53%. The total hen day production and pattern of egg drop is shown in Fig. 1. The infected birds started laying abnormal eggs at 9 DPI and this remained upto 21 DPI. The number of abnormal eggs laid was maximum (42%) at 13 DPI. These abnormal eggs were soft shelled, thin shelled, shell less and smaller in size (Fig. 2). The number of abnormal eggs laid gradually decreased but it remained up to 6% even after 21 DPI, when the aberrant eggs were excluded from the total daily egg production, the lowest egg production was observed at 13 DPI and the drop was at level of 47% which remained up to 20% at 21 DPI. Internal egg quality was also affected. The effects on egg quality characters viz., egg weight, IQU, shell thickness are depicted in Table 1. The mean egg weight of the infected birds was significantly lower than those of the control at 13-15 DPI (p<0.01) and gradually there was improvement in egg weight after 19 DPI. The IQU of eggs laid were lower (57.16±0.54-58.16±0.44) during 11-16 DPI. The IQU value of eggs laid by the infected quails was lowest at 13 DPI (55.16±0.58) as compared to (61.42±0.26) in control birds, which gradually increased and became par with control group after 19 DPI.
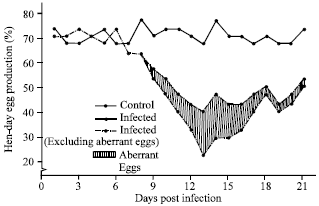 | |
| Fig. 1: | Effect of EDS-76 virus infection on egg production of quails |
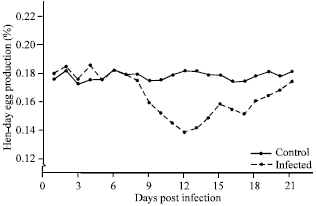 | |
| Fig. 2: | Effect of EDS-76 virus infection on egg shell thickness of quails |
| Table 1: | Effect of experimental EDS-76 virus infection on egg quality traits of Japanese quails |
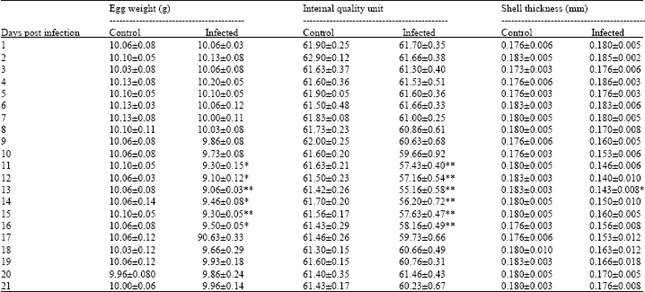 | |
| Each value is the Mean±SE of ten observations, Significance of difference from control values: *p<0.05, **p<0.01 | |
Similarly, the mean shell thickness of eggs laid by the infected birds was lowest (0.140±0.01 mm) at 10-13 DPI, which gradually increased and became normal at 20 DPI onwards.
The pattern of egg drop and production of abnormal eggs was similar to the findings reported in chicken (Yamaguchi et al., 1981b; Brugh et al., 1984; Van Eck 1986; Kumar, 1990; Swain et al., 1993; Ezeibe et al., 2008). The egg production recovered after 21 DPI but the laying of abnormal eggs (46%) persisted till the end of the entire observation period.
Gross pathological changes: The gross changes in infected Japanese quails were confined to uterus and spleen were similar in nature as reported earlier in case of chicken (Van Eck 1986; Taniguchi et al., 1981; Kumar, 1990; Swain, 1991). The uterine folds were swollen during 9-18 DPI. Exudate in the uterine lumen was present during 12-15 DPI. At 2 and 6 DPI, the spleen was found to be congested. Splenomegaly was present during 6-15 DPI. However, no gross abnormalities were detected in vagina, ovary and any other organ.
Histopathology: Significant histopathological changes were observed in uterus, spleen, vagina and ovary. Early changes in uterus were detected at 3 DPI and characterized by oedema and congestion of the blood vessels which progressed further at 6 DPI when there was mononuclear cells infiltration into mucosa, focal hyperplasia of the surface epithelium and pyknosis of the nuclei of glandular cells. At 9-12 DPI, extensive hyperplasia of uterine epithelium, focal lymphoid cell infiltration into submucosa and muscular layer and atrophy of the tubular glands was marked (Fig. 3a). The development of lesions corresponded with the egg drop and egg shell abnormalities at 9-15 DPI.
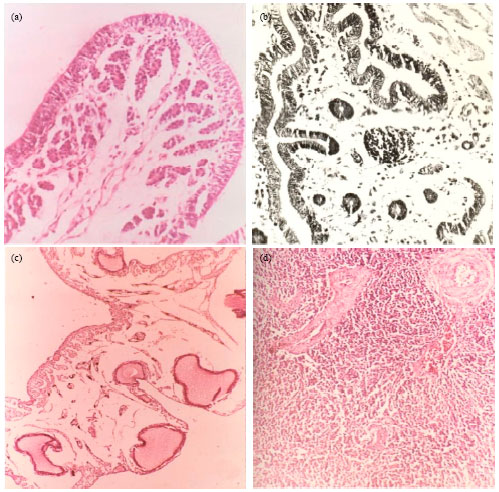 | |
| Fig. 3(a-d): | Histopathological changes in the tissue sections of EDS-76 virus infected quail (a) Uterus showing oedema and depletion of glands in submucosa at 9 DPI (H and Ex200), (b) Vagina showing lymphoid follicle and mononuclear cell infiltration in submucosa at 21 DPI (H and Ex200), (c) Ovary showing distorted follicles at 18 DPI (H and Ex50) and (d) Spleen showing a number of secondary follicles in Periarteriolar Lymphoid Tissue (PALT) around the central arteriole at 15 DPI (H and Ex200) |
Intranuclear inclusion bodies in goblet cells and ciliated cells of uterine epithelium and uterine glandular cells were present at 15-18 DPI. During this time more empty spaces were seen in submucosa due to depletion of glands. The condition gradually improved and by 28 DPI there was mild focal hyperplasia of epithelium, mononuclear cells infiltration and presence of lymphoid follicles. All these findings were similar to the observations made by earlier workers (Taniguchi et al., 1981; Yamaguchi et al., 1981b; Van Eck, 1986; Kumar, 1990; Swain et al., 1993) in case of experimentally infected chickens.
In vagina, the changes were observed during 9-28 DPI. At 9-12 DPI, mononuclear cells infiltration was marked in submucosa but during 15-18 DPI it was also noticed in the muscular layer. The lymphoid follicle formation in vaginal submucosa was seen during 21-28 DPI (Fig. 3b). The changes were similar to those observed by Kumar (1990). In ovary, the mononuclear cells infiltration was observed as early as 6 DPI which persisted upto 12 DPI during which the lymphoid follicles were present in ovarian tissue. During 9-18 DPI the number of developing primary ovarian follicles was few and during 15-18 DPI these follicles were distorted and collapsed (Fig. 3c). Badstue and Smidt (1978) and Taniguchi et al. (1981) also reported similar changes in ovary of chickens infected with EDS-76 virus.
In spleen, the microscopic changes were first noticed after 3 DPI which consisted of congestion, presence of disrupted follicles, large lymphoblasts in follicles and karyorrhexis of macrophages in red pulp. At 6-9 DPI, these changes were present along with focal areas of lysis of macrophages and presence of secondary follicles in Peri-arteriolar Lymphoid Tissue (PALT). During 12-15 DPI these secondary follicles were seen in PALT (Fig. 3d) which were found to be devoid of reticular capsule and consisted of large number of mesenchymal cells and macrophages by 5-18 DPI. In spleen the changes were same as the report of Swain et al. (1993). No microscopic changes were observed in any tissues of healthy birds throughout the observation period. The changes in ovary were marked by infiltration of mononuclear cells and lymphoid follicle formation during 6-12 DPI with presence of few and distorted follicles.
Localization of viral antigens: In FAT, granular fluorescence was detected in the nuclei of uterine epithelium as early as 6 DPI. At 9 DPI, the number of fluorescing cells increased and at 12 DPI the glandular cells showed fluorescence. At 21 DPI, few epithelial cells were still showing fluorescence. In circulating blood leukocytes, fluorescent antigen was detected at 4-6 DPI. In spleen, granular fluorescence was seen in macrophages of red pulp as early as 3 DPI. At 6 DPI, the intensity was maximum and more cells were involved. In vagina, fluorescence was marked in epithelial cells only at 12 DPI, where as in nasopharynx it was only observed at 3 DPI. Granular immunofluorescence was also demonstrated in nuclei of epithelial cells which persisted till 21 DPI. No fluorescing antigen could be detected in any of the three organs of the control birds. These findings are in accordance with those observed in chickens infected with CEDS-76 virus (Yamaguchi et al., 1981b; Van Eck, 1986; Kumar, 1990).
Virus excretion and contact transmission: EDS-76 virus, following naso-oral infection, was re-isolated from cloacal swabs of experimental birds upto 6 DPI. However, no virus was isolated from control birds. Transmission of the virus to in-contact birds was evidenced by sero-conversion in healthy birds. The healthy birds kept in contact with the infected layers developed HI-antibodies at 5 WPI. However, the antibody titre was very low and persisted only for a short duration. Out of 10 birds only two birds showed HI-antibody titre of 21-22, these birds were found positive by ELISA and AGPT. These findings are indicative that the degree of contact transmission is fully dependent on the housing system of the birds. This study suggest contagiousness of EDS-76 virus in quails as agreed with the reports of Heffels et al. (1982) and Swain et al. (1993).
Serology: The infected birds also developed antibodies as early as 1 WPI, which was successfully detected using HI, ELISA and AGPT (Adair et al., 1986; Swain et al., 1993; Piela and Yates, 1983; Bishop and Cardozo, 1996). The result of humoral immune response of experimentally infected birds is presented in Table 2. From the table it is evident that the HI-antibody (mean HI titre 26.1) was detected at 1 WPI which increased slightly to 27.2 at 2 WPI but gradually declined and during 6-18 WPI the antibody titre was as low as 22-23.
| Table 2: | Humoral immune response and virus excretion pattern in Japanese quails experimentally infected with EDS-76 virus |
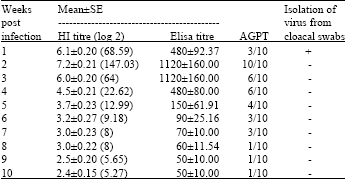 | |
| Values in parentheses indicate the geometric mean of HI titres, In AGPT column, Numerator: No. of sera samples found to be positive; -- Denominator: No. of sera samples tested, -: Negative, +: Positive | |
| Table 3: | Effect of experimental EDS-76 virus infection on serum total protein, albumin and globulin |
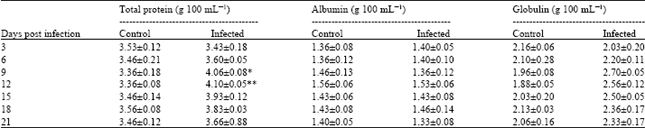 | |
| Each value is the Mean±SE of three observations, Significance of difference from control values, *p<0.05, **p<0.01 | |
| Table 4: | Effect of experimental EDS-76 virus infection on serum calcium and inorganic phosphorus content of Japanese quails |
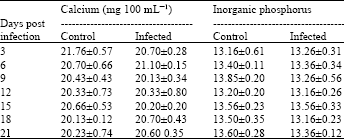 | |
| Each value is the Mean±SE of three observations | |
All the infected birds were positive by ELISA and highest mean ELISA titre of 1120±160 was detected at 2 WPI. The ELISA titre raised parallel to the HI-antibody and remained 10-12 times higher than the later during the observation period. The precipitating antibodies to EDS-76 virus were first detected in 3 out of 10 birds in 1st week. At 2 WPI, all the birds developed precipitins. The titre of precipitins was very low and only at 2 WPI all the birds were screened positive by AGPT. The precipitins level also declined afterwards and after 8 WPI the birds were found negative for the presence of precipitins. The poor immune response to EDS-76 virus in Japanese quails might be due to species factors associated with it. Das and Pradhan (1992) also detected low HI-antibody titre in the quails of old ages.
Biochemical studies: The mean values of total serum protein, calcium, phosphorus, albumen and globulin are depicted in Table 3 and 4. The mean total protein values in infected quail gradually increased and the highest value of 4.10±0.05/100 mL was observed at 12 DPI, thereafter the value declined gradually and remained higher than that of the control birds. No change in the mean albumen value (1.56±0.06/100 mL) was observed at 12 DPI. The values of calcium and phosphorus of blood serum of the infected birds was nearly the same as those of control. There are conflicting reports available regarding the changes of these two minerals in the serum. Van Eck and Vertommen, (1984) and Asi et al. (1987) on one hand did not find any difference in these values, where as Kumar (1990) reported significant increase in serum phosphorus and calcium level, respectively. However, there is no significant change in these values in the Japanese quails in the present report.
CONCLUSION
In conclusion, the course of EDS-76 disease was found equally significant both in chickens and quails inspite of the fact that it occasionally occurs in quails. The pathology and pathogenesis, changes in blood biochemical parameters during disease course and immunological studies for EDS-76 virus need to be elaborately explored in this species of birds. This will altogether help in understanding the disease process in quails in a better way, which would aid in designing effective vaccines and formulating appropriate disease prevention and control measures for this economical important poultry disease.
REFERENCES
- Adair, B.M., J.B. McFerran, T.J. Connor, M.S. McNulty and E.R. McKillop, 1979. Biological and physical properties of a virus (strain 127) associated with the egg drop syndrome 1976. Avian Pathol., 8: 249-264.
CrossRef - Badstue, P.B. and B. Smidt, 1978. Egg-drop syndrome 76 in Danish poultry. Nordisk Veterinaermedicin, 30: 498-505.
PubMed - Bidin, Z., I. Lojkic, M. Mikec and B. Pokric, 2007. Naturally occurring egg drop syndrome infection in turkeys. Acta Veterinaria Brno, 76: 415-421.
CrossRef - Bishop, S.C. and P. Cardozo, 1996. Egg drop syndrome 76 in Bolivia. Trop. Anim. Health Prod., 28: 199-206.
CrossRef - Brugh, M., C.W. Beard and P. Villegas, 1984. Experimental infection of laying chickens with adenovirus 127 and with a related virus isolated from ducks. Avian Dis., 28: 168-178.
Direct Link - Das, B.B. and H.K. Pradhan, 1992. Outbreaks of egg drop syndrome due to EDS-76 virus in quail (Coturnix coturnix japonica). Vet. Rec., 131: 264-265.
PubMed - Ezeibe, M.C., O.N. Okoroafor, J.I. Eze and I.C. Eze, 2008. Seroprevalence of egg drop syndrome-76 virus as cause of poor egg productivity of poultry in Nsukka, South East Nigeria. Trop. Anim. Health Prod., 40: 137-140.
PubMed - Heffels, U., S.E.D. Khalaf and E.F. Kaleta, 1982. Studies on the persistence and excretion of egg drop syndrome 1976 virus in chickens. Avian Pathol., 11: 441-452.
CrossRef - Kataria, J.M., P. Swain, B.B. Dash and K.C. Verma, 1991. Egg drop syndrome-76 (EDS-76) virus infection in Japanese quail. Proceedings of the Souvenir 12th Annual Conference of IAVMI and National Symposium on Important Infectious Diseases of Livestock and Poultry, September 12-14th, 1991, Tirupati, India, pp: 6.
- Kataria, J.M., C.M. Mohan, S. Dey, B.B. Dash and K. Dhama, 2005. Diagnosis and immunoprophylaxis of economically important poultry diseases: A review. Indian J. Anim. Sci., 75: 555-567.
Direct Link - McFerran, J.B., R.M. McCracken, E.R. McKillop, M.S. McNulty and D.S. Collins, 1978. Studies on a depressed egg production syndrome in Northern Ireland. Avian Pathol., 7: 35-47.
CrossRef - Piela, T.H. and V.J. Yates, 1983. Comparison of enzyme-linked immunosorbent assay with hemagglutination-inhibition and immunodiffusion tests for detection of antibodies to a hemagglutinating duck adenovirus in chickens. Avian Dis., 27: 724-730.
Direct Link - Snedecor, G.W. and W.G. Cochran, 1967. Statistical Methods. 6th Edn., Iowa State University Press, Ames, Iowa, ISBN: 9780813815602, Pages: 593.
Direct Link - Su, J., S. Li, X. Hu, X. Yu and Y. Wang et al., 2011. Duck egg-drop syndrome caused by byd virus, a new tembusu-related flavivirus. PloS ONE, Vol. 6.
CrossRef - Swain, P., J.M. Kataria, K. Dhama and K.C. Verma, 1997. Purification of egg drop syndrome-76 virus by velocity density gradient centrifugation. A comparative study. Acta Virol., 41: 303-304.
PubMed - Swain, P, J.M. Kataria and K.C. Verma, 1993. Biological characterisation of an Indian isolate of egg drop syndrome-76 virus. Res. Vet. Sci., 55: 396-397.
PubMed - Taniguchi, T., S. Tamaguchi, M. Maeda, H. Kawamura and T. Horiuchi, 1981. Pathological changes in laying hens inoculated with the JPA-1 strain of egg drop syndrome-1976 virus. Nat. Inst. Anim. Health Q., 21: 83-93.
PubMed - Van Eck, J.H., F.G. Davelaar, T.A. Heuvel-Plesman, N. van Kol, B. Kouwenhoven and F.H. Guldie, 1976. Dropped egg production, soft shelled and Shell-less eggs associated with appearance of precipitins to adenovirus in flocks of laying fowls. Avian Pathol., 5: 261-272.
PubMedDirect Link - Van Eck, J.H.H. and M. Vertommen, 1984. Biochemical changes in blood and uterine fluid of fowl following experimental EDS'76 virus infection. Vet. Q., 6: 127-134.
CrossRef - Yamaguchi, S., T. Imada, H. Kawamura, T. Taniguchi and M. Kawakami, 1981. Pathogenicity and distribution of egg-drop syndrome-1976 virus (JPA-1) in inoculated laying hens. Avian Dis., 25: 642-649.
PubMed - Yamaguchi, S., T. Imada, H. Kawamura, T. Taniguchi, H. Saio and K. Shimamatsu, 1981. Outbreaks of egg-drop syndrome-1976 in Japan and its etiological agen. Avian Dis., 25: 628-641.
Direct Link