
Zhien Pu
College of Agronomy, Sichuan Agricultural University, Wenjiang, 611130, People�s Republic of China
Guoyue Chen
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, 611130, People�s Republic of China
Jirui Wang
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, 611130, People�s Republic of China
Yaxi Liu
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, 611130, People�s Republic of China
Qiantao Jiang
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, 611130, People�s Republic of China
Wei Li
College of Agronomy, Sichuan Agricultural University, Wenjiang, 611130, People�s Republic of China
Xiujing Lan
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, 611130, People�s Republic of China
Shoufen Dai
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, 611130, People�s Republic of China
Yuming Wei
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, 611130, People�s Republic of China
Youliang Zheng
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, 611130, People�s Republic of China
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 6 | Page No.: 792-801







ABSTRACT
In this study, the ARF genes were cloned, sequenced and located on the chromosomes. The gene expression of various stress conditions were analyzed through RT-PCR. Two important features of ARF in wheat were found: (1) High sequences homology among species in mammalian and plant and (2) Four exons and three introns were conserved in Poaceae. In this study the coding genes of ADP-ribosylation Factors (ARF) were characterized and they were located on chromosomes 3AL and 2DL in common wheat and its diploid progenitors. Forty-seven candidate SNPs in ARF were detected which were located in exons (17 SNPs) and introns (30 SNPs), respectively. As expected, most of the SNPs (66.34%) in ARF were transitions and the rest (33.66%) were transversions. The expression difference of ARF under various environmental stresses (low-temperature, Abscisic Acid (ABA), Polyethylene Glycol (PEG), NaCl, stripe rust), in two stages (seedling and maturity) and in different tissues (root, stem, flag leaf and immature embryo) of 15 days post-flowering were investigated. The results revealed that the expression levels of ARF were affected by environmental stresses. PEG stress induced the highest level of ARF expression, followed by the stripe rust and ABA stresses.
PDF Abstract XML References Citation
Received: January 30, 2013;
Accepted: August 31, 2013;
Published: January 29, 2014
How to cite this article
Zhien Pu, Guoyue Chen, Jirui Wang, Yaxi Liu, Qiantao Jiang, Wei Li, Xiujing Lan, Shoufen Dai, Yuming Wei and Youliang Zheng, 2014. Characterization and Chromosome Location of ADP-ribosylation Factors (ARFs) in Wheat. Pakistan Journal of Biological Sciences, 17: 792-801.
DOI: 10.3923/pjbs.2014.792.801
URL: https://scialert.net/abstract/?doi=pjbs.2014.792.801
DOI: 10.3923/pjbs.2014.792.801
URL: https://scialert.net/abstract/?doi=pjbs.2014.792.801
INTRODUCTION
ADP-ribosylation Factors (ARF) were GTP-binding proteins (Brown et al., 1993; Memon, 2004). ARF1 has been known that it plays an important role in vesicular transport between the ER and the Golgi and maintains the stability of the Golgi and ER organization (Takeuchi et al., 2002; Lee et al., 2002). ARF are a critical component in the regulation of vesicle-mediated secretary pathway in eukaryotic cells. The functions of ARF proteins played an important role in keeping the membrane traffic and organelle integrity, at the same time, the ARF proteins were tied to its reversible association with membranes and specific interactions with membrane phospholipids (Donaldson, 2003; Nickel and Wieland, 1997; Bonifacino and Glick, 2004). The activity of ARF is regulated by the binding and hydrolysis of GTP, as they cycled between active GTP-bound and inactive GDP-bound conformational states. The active GTP-bound state is regulated by the guanine nucleotide exchange factors (GEFs) which promote the exchange of GDP to GTP. The form change between inactive GDP-bound and active GTP-bound were selectively regulated by ARF cycle (Jackson and Casanova, 2000). Based on phylogenetic lines, ARF are abundant, ubiquitous and highly conserved in eukaryotes. Although the ARFS have been mainly studied in mammalian and yeast, several orthologues have been identified in plants.
In recent studies, most of ARFs were cloned and identified from various plant species, such as rice (Higo et al., 1994), wheat (Kobayashi-Uehara et al., 2001), carrot (Asakura et al., 2007), Arabidopsis (Regad et al., 1993) and so on. The features and functions of ARF genes in plant, yeast and mammalian are similar. They are found to be mainly active in roots and flowers (Kobayashi-Uehara et al., 2001). In Arabidopsis, overexpression of antisense ARFA1 reduces cell division and cell expansion which lead to antisense plants being severely stunted (Gebbie et al., 2005). ARF is important for wheat growth and development and the upregulated expression of this gene might contribute to the heterosis observed in wheat root and leaf growth in wheat (Yao et al., 2009). Although these studies on ARFs in different plants are not complete, the findings have so far provided a reliable view on the general functions of ARFs. In this study, ARF gDNA sequences, chromosomes location, splicing pattern in the wheat were determined. The results also revealed that the ARF expression was affected under various environmental stresses.
MATERIALS AND METHODS
Plant materials: It was thought that the genomes of cultivated wheat (Triticum aestivum, AABBDD) were derived from T. urartu (A), Aegilops speltoides (B) and Ae. tauschii (D), respectively. In this study, the cultivated wheat and its diploid putative progenitors were chosen to characterize ADP-ribosylation factor. Ten T. monococcum (2n = 2x = 14, AmAm), 10 T. urartu (2n = 2x = 14, AuAu), 5 Ae. tauschii (2n = 2x = 14, DD), 5 Ae. speltoides (2n =2x =14, SS), 5 spelt wheat and 6 cultivated wheat varieties were used (Table 1). These diploid wheat and Aegilops accessions were kindly supplied by the USDA-ARS. Plants grown in normal condition were used for gDNA cloning, cDNA cloning and RT-PCR.
DNA isolation and PCR amplification: CTAB method was used for the extraction of genomic DNA as previously described. Based on the alignment of 7ARF gene sequences, primers were designed to amplify the complete Open Reading Frames (ORF) of ARF gene. PCR amplification was performed with a GeneAmp PTC-200 cycler (MJResearch,USA) in 50 μL volume which included of 50 ng of genomic DNA, 0.2 mmol L-1 of dNTPs, 150 ng of oligonucleotide primers, 1.5 U ExTaqTM polymerase (TaKaRa), 2 mmol/MgCl2 and 5 μL 10xPCR buffer. PCR amplifications were conducted using the following thermal cycle: 95°C for 300 sec, followed by 35 cycles of 60 sec at 95°C, 45 sec at 57-60°C (depending on the primer sets and 45 sec at 72°C. The desired DNA fragments were recovered from gels and ligated to the pBluescript SK (+) T-vector plasmid (Stratagene). The positive clones were screened and sequenced.
The expression level of ARFs was analyzed by RT-PCR. RNA was prepared from the roots, leaves, stems and immature embryo of the cultivated wheat plants. Seedlings were treated separately with NaCl (200 mM), Abscisic Acid (ABA) (50 mM) and Polyethylene Glycol (PEG) (150 mM) for 48 h, low temperature (4°C) for 24 h and yellow rust for 72 h at the same time. At regular intervals, 100 mg of the seedlings were harvested and the total RNA was isolated from the samples as described above. The RNA extraction was checked by electrophoresis. Aliquots of RNA solutions (approximately 0.5 mg RNA equivalent) were added to the RT-PCR mixture prepared from the one step RT-PCR Kit (TakaRa).
ARFs sequence analysis: More than 90 complete and partial ARF Expressed Sequence Tag (EST) sequences retrieved from wheat, barley, maize, rice, human and Arabidopsis were obtained from the GenBank. To concentrate on the coding region alone, 5’ and 3’ non-translated regions and several ultra-short sequences were omitted. A total of 54 ARF sequences were left for alignment analysis using MEGA 3.0. The alignment results showed that ARFs were highly conserved.
The analysis of full-length sequence and the construction of subsequent nucleotide sequences were carried out under DNAman 4.0. The multiple sequence alignment software Clustal W (http://www.ebi.ac.uk/clustalw) was used for the SNP evaluation. ARFs were translated into amino acid sequences using cDNA sequences. The phylogenetic trees were constructed by MEGA, version 2 (Wang et al., 2004). ARF sequences were subject to nucleotide and amino acids alignment using CLUSTAL X program (ver.1.83).
Structure predictions: The three secondary structure elements are a helical region known as a-helix (H), a coil/loop and an extended region known as h-stand which joins a-helix together to form h-sheet (E). Fold recognition, disulfide bridges analysis and structural alignments were performed with ANTHEPROT software (Wang et al., 2004). To compare the secondary structure of ADP-ribosylation factors, 3D-PSSM protein fold recognition (threading) server (http://www.emblheidelberg.de/predictprotein) was used to assume the predicted secondary structure of these ADP-ribosylation factors.
| Table 1: | Material source and name |
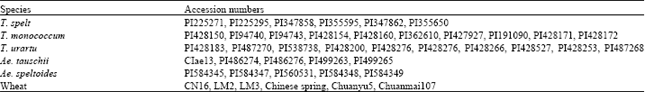 | |
| CN16: Chuannong 16; LM2: Liangmai 2; LM3: Liangmai 3. They are common wheat | |
RESULTS
Sequence analysis and ARF gene classification: In order to identify ARFs in wheat, a DNA segment 1195 bp was isolated from all samples. A number of gDNA clones was randomly selected and sequenced. A total of 73 sequences were obtained. The comparative analysis of the partial nucleotide sequences was carried out by BLAST for further characterization of gDNA encoding ARFs or ARF-like small GTP binding proteins. The sequencing results showed that these DNA amplified segments were indeed ARFs. A sample of gDNA exhibited a high homology (88-100%) among these clones. Compared with ARF sequences isolated from wheat and other species, such as yeast, human, carrot, rice, maize, A. thaliana and so on. The ARFs were isolated from barley and rice had higher sequence homology, up to 98%. The ARFs are also highly conserved in Arabidopsis (83%) and Brassica (84%). The results showed that the ARFs were conserved across species and also indicated that they may have important functional roles.
The sequences isolated from diploid species and hexaploid wheat also were analyzed. The results showed that the ARFs had two and three different sequences in diploid species and hexaploid wheat, respectively, so the ARFs might have two copies in diploid and three copies in hexaploid wheat which is consistent with previous studies (Kobayashi-Uehara et al., 2001).
Sixty six of the 73 sequences were not the same and the rest of the sequence is identical. The 66 different sequences were selected for further analysis using the Paup software. Phylogenetic analysis based on these sequences indicated that the 66 sequences could be divided into two groups. Most of sequences derived from A genome were clusted in Group I and Group I could be further branched into two subgroups again (Fig. 1). T. monococcum, Triticum urartu and part of the sequences isolated from hexaploid chromosome which had same haplotype with A chromosome were clustered in Group I.2. It is worth mentioning that the Iran and Europe spelt wheat did not differ from each other. Group I.1 included the sequences from PI225295 (Iran) and PI347862 (Europe). Group II included all sequences isolated from D and S chromosome, as well as sequences which had similarities with haplotype from the hexaploid chromosome. Furthermore, the deduced amino acid sequences were divided into two subgroups.
Intron variations among different species: These DNA clones had three introns and four exons compared with the AY736124 sequence that was isolated from common wheat. Due to sequence mutations, the total length of the introns varied between 754 and 779 bp. The three introns were located on 162-480, 539-731, 852-1118 bp and the length were 319, 192 and 266, respectively. A number of ADP-ribosylation factor genes from cultivated wheat and its diploid putative progenitors had the high sequence homology. Thus, 66 different ADP-ribosylation factor genes were selected for further analysis. The results showed that intron splicing had three different patterns. Intron 1 splicing occurred between GU-GA, intron 2 splicing occurred between AG-AC and intron 3 occurred between AG-AU. These three splicing patterns were fixed and not consistent with typical splicing patterns which were splicing between GU-AG and special splicing site AG in intron 2 and 3. Analyzing the splice site revealed that the sequence 2-3 bp upstream the splicing site was conserved. The sequence isolated from eukaryotes such as Saccharomyces cerevisiae, Arabidopsis thaliana, Drosophila melanogaster, Giardia lamblia and other lower organisms, mice, cows and other mammals and humans had demonstrated that the structure and function of the ARFs in plants and animals and evolution are highly conserved.
The ARF sequences of Arabidopsis thaliana, rice, Zea maize and Brachypodium distachyon and analyzed them using online BLAST. The results showed that the ARF in Arabidopsis thaliana also had three introns and four exons. A comparison of published gene ARFs sequences across species revealed an important variation in the number of introns (and consequently of exons) present in the gene (Fig. 2): A. thaliana, Zea maiz, Oryza sativa and Triticum aestivum L. contained 3 introns while Brachypodium distachyon contained 2 introns. Sequence alignment of the different exons was performed using ClustalW (data not shown) and the result suggested a reduction in the number of exons. Exon1 in Triticum aestivum L. could be aligned to exons (1+2) from Brachypodium distachyon (Fig. 2).
The intron splicing patterns observed in different species were listed in Table 2. The results showed that the patterns were different among different species. However, some common features in these patterns were found which were that intron 2 and 3 had the same splice sites AG and intron 2 had the splice sites C, intron 3 had splice sites U. That ADP-ribosylation factor might have the similar splice pattern in common wheat and its diploid ancestors.
gDNA SNP analysis in wheat: Although the ARFs in wheat were much conserved in their lengths and sequence homology, there were about 47 candidate SNPs in those sequences. Among the 47 SNPs, 17 were located in exons and 30 located in introns.
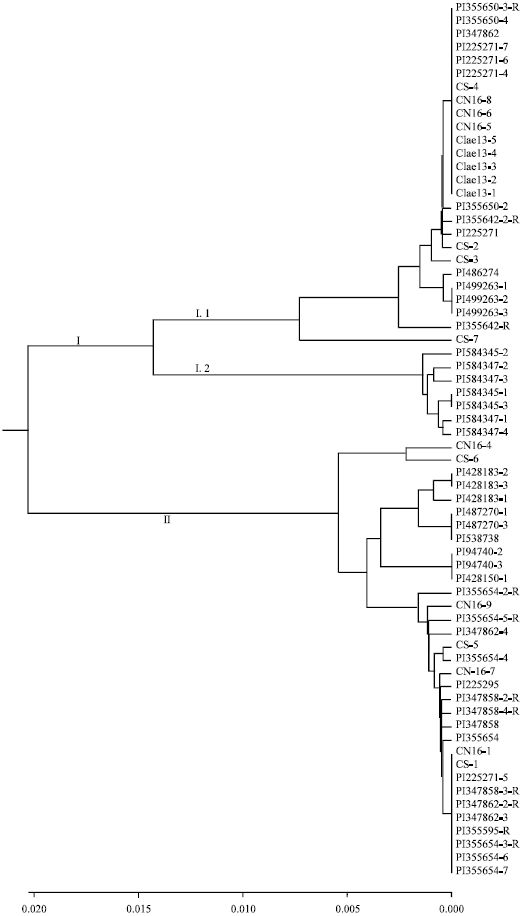 | |
| Fig. 1: | Homologous tree based on the ARF sequences |
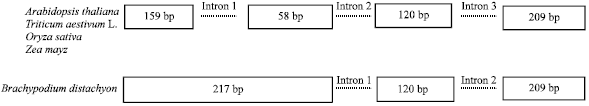 | |
| Fig. 2: | ARF genomic DNA structure inferred from published sequences from different species and alignments between the exons using Clustal W |
| Table 2: | ARF gDNA intron splicing patterns across species |
 | |
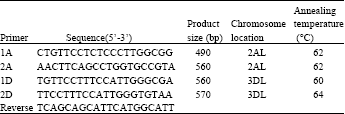
| Table 3: | Information of Group-specific primer sets of each group of ARF gene |
 | |
| ARF gene: ADP-ribosylation factors (ARFs) | |
| Table 4: | Haplotypes of ARF in the plant materials |
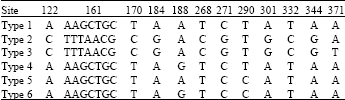 | |
The SNPs in ARFs were further investigated and the results showed that the transition was more common than the transversions among SNPs (Cooper and Krawczak, 1990). About 41% of the substitutions were found in the accurate CpG site, as adenine nucleotide was known for its high mutability (Cooper and Youssoufian, 1988). The distribution and proportion of transitions and transversions in the ARF SNPs were investigated (Table 4). As expected, most of the SNPs (66.34%) in ARFs were transitions and the rest (33.66%) were transversions. In the substitution type, the transversion type T→C was the most common while the transversions type A→G was second most common.
The phylogenetic results and the substitution analysis showed that the SNPs were clustered into six haplotype and three kinds of haplotypes appeared only once. Haplotypes were associated with chromosomes. D chromosome had the haplotype type I, Au, Am and S chromosome had the same haplotype type 2. The only difference between type 2 and 3 was at site 371bp (T/A). There was only 1bp difference in the remaining type.
The variations deduced by insertion, deletion and SNPs in the introns were less than the exons. Introns had more variations and intron 1 was more conserved than the intron 2 and intron 3.119 SNPs in about 750 bp intron sequences and a part of the SNPs showed genome specificity.
ARF chromosome assignment by group-specific primer sets: Specific-primers based PCR assay was carried out to identify the chromosome locations of each group of ARFs. A total of 10 specific primer sets were designed on A and D chromosomes. The forward primers annealed at the beginning part of the SNP domain and the reverse primer annealed at the C-terminal domain. Due to the high sequence homology of ARFs across species, an additional primer-template mismatched base was added in a few primer sequences at their position 3’ end (Table 3). The gDNA sequence of A and D chromosomes in the ditelosomic lines of CS were used to carry out the specific PCR amplifications. Therefore, each primer set should yield uniformly sized products in ditelosomiclines. Furthermore, the chromosomal location would be consequently recognized by the absence of the PCR products from one particular ditelosomicline which had lost specific chromosome arms.
Two out of ten ARF groups were successfully assigned to specific chromosome arms on the A and D chromosomes (Table 3). The ARFs isolated from A chromosome were assigned to the long arm of chromosome 2A and the ARFs isolated from D chromosome were assigned to the long arm of chromosome 3D. The specific primer sets for S chromosome did not found. All primer had the same reverse primer.
ARF amino acid sequence analysis and protein structure prediction: The three conserved characteristic motifs (Kahn et al., 1995) unique to the GTP-binding protein super family involved in GTP-binding and/or hydrolysis are: GLDAAGKT (Fig. 1), DVGGQ and ANKQD. The structure of the ARFs includes the effector region binding to the GAPs (Amor et al., 1994; Greasley et al., 1995; Vitale et al., 1997); the switch 1 and switch 2 regions binding to GEF Sec7 domains (Mossessova et al., 1998) a potential myristoylation site at Gly-2 (Kahn et al., 1992; Antonny et al., 1997) the N-terminal 17 amino acids which impart ARF properties to a Drosophila (Kahn et al., 1992) residues 35-94 which activate phospholipase D and recruit adaptor protein AP-1 (Liang et al., 1997).
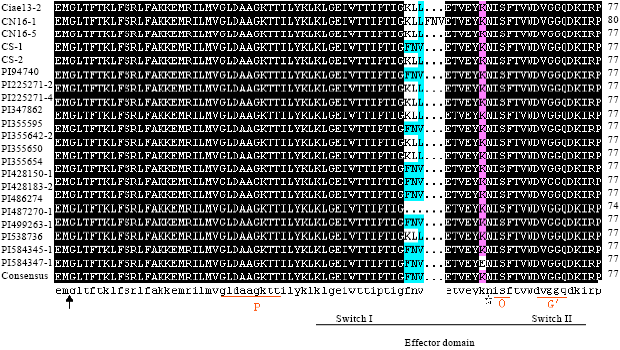 | |
| Fig. 3: | Partial alignment of deduced amino acids sequence |
The putative amino acid sequences of these ARFs showed significant sequence homology to the ARFs from Arabidopsis thaliana, rice and other species. The ARFs also were conserved in other functional site, such as the asparagine glycosylation site at position 61.
The correlation between SNPs and the amino acid alteration in the ARFs was limited. It was clear that the 73 SNPs only resulted in four amino acid variations. As 94.5% of the candidate SNPs resulted synonymous changes (Fig. 3). Most of these variations were a potential way to obtain information about the functional constraint via evolutionary comparisons, as it is generally believed that conserved residues are more likely to be functionally significant than non-conserved residues. The SNPs did not alter the amino acid sequences of the GTP-binding and the hydrolysis domain. The results illustrated the GTP-binding and hydrolysis functions of the ARFs were conserved. The secondary structure of the ARFs in wheat deduced from the Swiss-Model was not altered and remained similar with the secondary structure of the ARFs in human.
ARF expression under various stresses in wheat: In this study, the effects of various environmental stresses, including low-temperature, ABA, PEG, NaCl, stripe rust, on the accumulation of ARF transcription in wheat were investigated. At the same time, the ARF expression level of 15 days post-flowering in root, stem, flag leaf and immature embryo were analyzed. The ARF expression level during the seedling stage and the maturity phase also were measured.
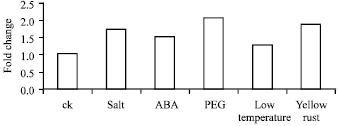
The ARF expression level was higher in root than other issue in the seeding stage and was higher in root and embryo in the maturity phase which suggested that the ARFs were differentially expressed in different tissue Fig. 4 (Kobayashi-Uehara et al., 2001). The results indicated that ARFs could be more active in the parts of wheat that had higher metabolism rate and the ARFs gene may be had higher expression in the meristem, such as root tip and young embryo. The ARF expression in the root was also up-regulated by 4.0 fold under the ABA stress and 2.3 fold under the PEG stress (Fig. 5 and 6). During the seeding stage, the ARF expression level was upregulated under various environmental stresses, such as low temperature, yellow stripe, ABA, PEG, NaCl treatment. PEG stress induced the highest ARF expression level compared to other environmental stresses, maybe the ARF gene regulate and control the growth of wheat in drought conditions. The ARFs expression level was higher in leafs and roots during the seeding stage than the maturity stage. The results showed that ARFs could be induced by several environmental stresses (Kobayashi-Uehara et al., 2001).
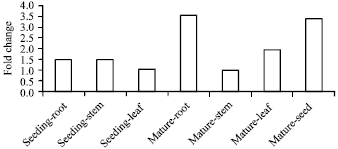 | |
| Fig. 4: | ARFs expression level of different stage in different issue. Seeding-root, seeding-stem, seeding-leaf represent the tissue originated from root, stem and leaf while the wheat was in seeding stage |
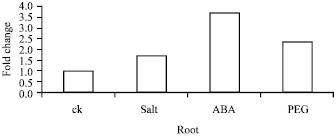 | |
| Fig. 5: | ARFs expression level of root under different stresses. The ARFs expression level of root under salt, ABA and PEG stresses |
 | |
| Fig. 6: | ARFs expression level of leaf under different stresses. The ARFs expression level of leaf under salt, ABA, PEG, low temperature and yellow rust stresses |
DISCUSSION
ARFs genomic characterization in wheat: In this study, two important features of the ARFs in wheat were found. The first feature was that ARF sequences in wheat had high homology with other species, such as mammalian, carrot, maize and so on. The second feature was that the ARFs in wheat contained four exons and three introns, consistent with the ARFs in A. thaliana, H. vulgare, Brachypodium, O. sativa and other species (Fig. 2). The genomic structure of the ARFs therefore, seemed to be very simple and conserved in the plant kingdom. The presence of intron 2 and 3 might be a common feature of Poaceae. Intron-1 was found in all species, but its bp length varied, even within the same Poaceae. The Poaceae family could be further divided based on the number of introns present: Three introns only in the sub-families Poaceae (Wheat, Arabidopsis thaliana, Oryza) and two introns in the sub-family Poaceae (Brachypodium). Interestingly, the length of intron-1 in Brachypodium distachyon was equal to the total length of intron 2 and 3 in Arabidopsis thaliana and wheat (Fig. 1, Table 2). Intron structure in wheat and Arabidopsis all contained three introns. The ancient species T. monococcum, T. urartu had three introns which implied that the three introns feature could represent an ancestral state.
The introns were removed precisely which is dictated by a splicing code integrated into the pre-mRNA. The 5’ splice site (GU) 3’ splice site (AG), polypyrimidine tract and the branch point adenosine residue near the 3’ splice site are core element of the splicing code during the splicing (Wang and Burge, 2008). Here, an alternative splicing site for ARFs which provided an interesting model for understanding different mechanisms involved in splice site selection. Though the cDNA sequences of the ARFs in Poaceae were conserved, the intron splicing sites were variable. The splicing sequences of the ARFs also were conserved. Conserved sequences in these introns are essential for the regulation of alternative splicing in response to stress (Singh et al., 2009). AG-rich enhancer sequence downstream of the three introns are defined and needed for efficient recognition of the exon splice site (Gallego et al., 1996). In this study, the splicing was chromosome-specific.
SNPs analysis in ARFs: In this study, sequence alignment for 73 genes derived from hexaploid and their closely related diploid wheat species were analyzed (Triticum and Aegilops). The alignment analysis revealed a relatively high sequence homology between different ARFs. Extron sequences were more variable than intron terminal domain. The ARFs were divided into two groups (Fig. 1) based on the exon sequences (Fig. 1). The ARFs also were classified into two groups based on the intron sequences (data not shown). A marker based PCR was employed to assign the chromosome location of each ARF group. The results indicated that two groups of ARFs were successfully assigned to specific chromosome arms. Moreover, two subgroups with relatively high-sequence identity were identified. The amplification results from diploid wheat and Aegilops accessions were in agreement with those in the Chines Spring ditelosomiclines. These results further supported the validity of ARF classification results. Therefore, the results in this study provided the possibility to predict the chromosome location of a new ARF using its cDNA sequence. The classification based on SNPs could simplify the characterization of ARF sequences. This was probably due to the limited number of genotypes of diploid species from which cultivated wheat was derived. Wheat is a hexaploid with genome constitution AABBDD which generally contains triplicated homologous genes derived from three diploid ancestral species (Feldman, 2001). There could be three possibilities for three chromosome-specific markers of homoeological genes in the polyploidy (Wendel, 2000). Although the chromosomal location of S-genome because S genome-specific PCR primer was unavailable, maybe the ARFs of B genome is homologous to ARFs of D genome located on based on the following three facts: Firstly, the ARFs of S genome showed high homology to D genome. Secondly, there were three homologous ARFs in the wheat genome (Kobayashi-Uehara et al., 2001) which was consistent to this results. Similar situations in WFLs in wheat have also been shown (Shitsukawa et al., 2006). Lastly, the phylogenetic analysis could not differentiate S and D genome and the conclusion also tested and verified the molecular marker results.
A new molecular marker system termed Intron Length Polymorphism (ILP) has been developed in rice by comparing the draft genomic sequences of indica cultivar and japonica cultivar (Wang et al., 2006). ILP is a codominant marker and can be conveniently detected by PCR with a pair of primers annealing at the flanking exons. ILPs are compatible among different species (Wang et al., 2006) because the exon-intron structures of ARFs are highly conserved (Rogozin et al., 2003; Roy and Gilbert, 2005; Lin et al., 2006; Yang et al., 2007). In addition, ILPs mainly exist in low frequency among (sub) species or higher taxonomic ranks. It is possible that these subspecies-specific ILP (SSILP) markers could well preserve the genetic information from wild progenitors and therefore could reflect the original genetic differentiation between the ancestors of wheat and its diploid putative progenitors. Moreover, introns usually have no biological functions. The above desirable features suggest that ILP markers should be quite suitable for studying wheat evolution. In the work described here, SSILP markers could be utilized to investigate the genetic differentiation of wheat in relation to the genetic differentiation among other species. Through the ARFs sequence analysis, that ARF genome-specific markers had two important features. First, they reflected the genetic differentiation between the two subspecies (Table 1). Second, the two subspecies-specific alleles were generally conserved. New markers might be developed for ARFs to investigate the evolution of wheat. In this study, the results indicated that the A- and D-genome had been clearly differentiated into two distinct groups. This suggested that genome-specific extron sequences could be found as neutral marker. Now Subspecies-specific Intron Length Polymorphism (SSILP) is developed as a neutral marker for selection in general because introns usually do not have biological functions in rice (Zhao et al., 2009).
ARF expression: The expressions of the ARFs were probed by antisense suppression. Antisense suppressed plants showed lowered stature and some plants with severe phenotypes died or were infertile (Gebbie et al., 2005). As for wheat, the protein level of ARFs was higher in root and flowers than in shoots, suggesting that wheat roots and flowers accumulated a relatively abundant amount of ARFs (Kobayashi-Uehara et al., 2001). The disruption of ARF1 could cause cold sensitivity and retarded growth which indicated that ARF1 played important roles in promoting cell survival (Stearns et al., 1990). Northern analysis of total RNA showed that ARFs were highly expressed in young seedlings and in seeds at the early developmental stage, but to a lesser extent in cultured cells (Higo et al., 1994). In this regard, the effects of various environmental stresses on ARF expression were focused and found that ARF expression level could be altered by different conditions, including cold stress. These results were not consistent with an earlier finding that the ARFs wheat was not cold inducible (Kobayashi-Uehara et al., 2001).
In this study, PEG stress induced the highest ARF expression level, followed by the stripe rust and ABA stress. RT-PCR revealed that ARFs were expressed at 2.0, 1.8 and 1.4 fold higher under PEG, stripe rust and ABA stressed, respectively. The ARFs could play an important role in DNA repair by self-modifying the transcripts. Cell division and cell expansion both depend on vesicle trafficking to deposit new wall material, to secrete proteins (e.g., expansins) that promote cell wall biogenesis and to deliver new plasma membrane proteins for cellulose synthesis and other needs (Samuels et al., 1995; Thiel and Battey, 1998; Cosgrove, 2000).
The relative abundance of ARF mRNA in roots and flag-leafs may indicate a high level of vesicular transporting activity in these tissues of wheat plants. The ARFs have been proposed to function through direct activation of enzyme activities, local alterations in lipid composition, or recruitment of proteins to the membrane (Donaldson et al., 1992). Therefore, the relative abundance of ARF mRNA in roots and embryos may indicate a high level of vesicular transporting activity in these tissues (Yao et al., 2009). These theories can explain why the expression level of ARFs was upregulated under ABA and PEG stresses, since cell production rate and final cell size were both reduced. Changed time-to-flowering, apical dominance and fertility may reflect alterations to hormonal and other signaling pathways with which ARFs may be associated with (Gebbie et al., 2005).
CONCLUSION
In conclusion, the ARFs gene sequences, SNP site, splicing pattern and gene expressions level under stresses were investigated in this study. These results showed that the ARFs gene sequences were conserved in the Poaceae family and the introns might have the similar splice pattern in common wheat and its diploid ancestors. The chromosome locations of the gene were located on 3AL and 2DL. The GTP-binding area of the ARFs was conserved and the secondary structure of ARFs in wheat was not altered with the nucleotide changing. Under environmental stresses, the gene expression level was higher than control. The ARFs were more active in the parts of wheat that had higher metabolism rate, such as root tip and young embryo.
ACKNOWLEDGMENTS
This study was supported by the National Basic Research Program of China (973 Program 2010CB134400) and China Transgenic Research Program (2011ZX08002).
REFERENCES
- Amor, J.C., D.H. Harrison, R.A. Kahn and D. Ringe, 1994. Structure of the human ADP-ribosylation factor 1 complexed with GDP. Nature, 372: 704-708.
CrossRef - Antonny, B., S. Beraud-Dufour, P. Chardin and M. Chabre, 1997. N-terminal hydrophobic residues of the G-protein ADP-ribosylation factor-1 insert into membrane phospholipids upon GDP to GTP exchange. Biochemistry, 36: 4675-4684.
CrossRef - Asakura, Y., E. Ishigaki, R. Sugiyama and F. Kurosaki, 2007. Cloning and expression of cDNAs encoding ADP-ribosylation factor in carrot seedling. Plant Sci., 172: 189-195.
CrossRef - Bonifacino, J.S. and B.S. Glick, 2004. The mechanisms of vesicle budding and fusion. Cell, 116: 153-166.
CrossRef - Brown, H.A., S. Gutowski, C.R. Moomaw, C. Slaughter and P.C. Sternwels, 1993. ADP-ribosylation factor, a small GTP-dependent regulatory protein, stimulates phospholipase D activity. Cell, 75: 1137-1144.
CrossRef - Cooper, D.N. and M. Krawczak, 1990. The mutational spectrum of single base-pair substitutions causing human genetic disease: Patterns and predictions. Hum. Genet., 85: 55-74.
CrossRefDirect Link - Cooper, D.N. and H. Youssoufian, 1988. The CpG dinucleotide and human genetic disease. Hum. Genet., 78: 151-155.
CrossRef - Cosgrove, D.J., 2000. Loosening of plant cell walls by expansins. Nature, 407: 321-326.
CrossRefDirect Link - Donaldson, J.G., 2003. Multiple roles for Arf6: Sorting, structuring and signaling at the plasma membrane. J. Biol. Chem., 278: 41573-41576.
CrossRefDirect Link - Donaldson, J.G., D. Cassel, R.A. Kahn and R.D. Klausner, 1992. ADP-ribosylation factor, a small GTP-binding protein, is required for binding of the coatomer protein β-COP to Golgi membranes. Proc. Natl. Acad. Sci. USA., 89: 6408-6412.
Direct Link - Gallego, M.E., P. Sirand-Pugnet, P. Durosay, B. Clouet d'Orval and Y. d'Aubenton-Carafa et al., 1996. Tissue-specific splicing of two mutually exclusive exons of the chicken β-tropomyosin pre-mRNA: Positive and negative regulations. Biochimie, 78: 457-465.
CrossRef - Gebbie, L.K., J.E. Burn, C.H. Hocart and R.E. Williamson, 2005. Genes encoding ADP-ribosylation factors in Arabidopsis thaliana L. Heyn.; genome analysis and antisense suppression. J. Exp. Bot., 56: 1079-1091.
CrossRef - Greasley, S.E., H. Jhoti, C. Teahan, R. Solari and A. Fensome et al., 1995. The structure of rat ADP-Ribosylation Factor-1 (ARF-1) complexed to GDP determined from two different crystal forms. Nat. Struct. Biol., 2: 797-806.
CrossRef - Higo, H., N. Kishimoto, A. Saito and K.I. Higo, 1994. Molecular cloning and characterization of a cDNA encoding a small GTP-binding protein related to mammalian ADP-ribosylation factor from rice. Plant Sci., 100: 41-49.
CrossRef - Jackson, C.L. and J.E. Casanova, 2000. Turning on ARF: The Sec7 family of guanine-nucleotide-exchange factors. Trends Cell Biol., 10: 60-67.
CrossRef - Kahn, R.A., J. Clark, C. Rulka, T. Stearns and C.J. Zhang et al., 1995. Mutational analysis of Saccharomyces cerevisiae ARF1. J. Biol. Chem., 270: 143-150.
CrossRefDirect Link - Kahn, R.A., P. Randazzo, T. Serafini, O. Weiss and C. Rulka et al., 1992. The amino terminus of ADP-Ribosylation Factor (ARF) is a critical determinant of ARF activities and is a potent and specific inhibitor of protein transport. J. Biol. Chem., 267: 13039-13046.
Direct Link - Kobayashi-Uehara, A., E. Shimosaka and H. Handa, 2001. Cloning and expression analyses of cDNA encoding an ADP-ribosylation factor from wheat: Tissue-specific expression of wheat ARF. Plant Sci., 160: 535-542.
CrossRef - Lee, M.H., M.K. Min, Y.J. Lee, J.B. Jin and D.H. Shin et al., 2002. ADP-ribosylation factor 1 of Arabidopsis plays a critical role in intracellular trafficking and maintenance of endoplasmic reticulum morphology in Arabidopsis. Plant Physiol., 129: 1507-1520.
CrossRefDirect Link - Liang, J.O., T.C. Sung, A.J. Morris, M.A. Frohman and S. Kornfeld, 1997. Different domains of mammalian ADP-ribosylation factor 1 mediate interaction with selected target proteins. J. Biol. Chem., 272: 33001-33008.
CrossRefDirect Link - Lin, H., W. Zhu, J.C. Silva, X. Gu and C.R. Buell, 2006. Intron gain and loss in segmentally duplicated genes in rice. Genome Biol., Vol. 7.
CrossRefDirect Link - Memon, A.R., 2004. The role of ADP-ribosylation factor and SAR1 in vesicular trafficking in plants. Biochim. Biophys. Acta, 1664: 9-30.
PubMedDirect Link - Nickel, W. and F.T. Wieland, 1997. Biogenesis of COPI-coated transport vesicles. FEBS Lett., 413: 395-400.
CrossRef - Regad, F., C. Bardet, D. Tremousaygue, A. Moisan, B. Lescure and M. Axelos, 1993. cDNA cloning and expression of an Arabidopsis GTP-binding protein of the ARF family. FEBS Lett., 316: 133-136.
CrossRef - Rogozin, I.B., Y.I. Wolf, A.V. Sorokin, B.G. Mirkin and E.V. Koonin, 2003. Remarkable interkingdom conservation of intron positions and massive, lineage-specific intron loss and gain in eukaryotic evolution. Curr. Biol., 13: 1512-1517.
CrossRef - Roy, S.W. and W. Gilbert, 2005. Rates of intron loss and gain: Implications for early eukaryotic evolution. Proc. Nat. Acad. Sci., 102: 5773-5778.
CrossRef - Samuels, A.L., T.H. Giddings Jr. and L.A. Staehelin, 1995. Cytokinesis in tobacco BY-2 and root tip cells: A new model of cell plate formation in higher plants. J. Cell Biol., 130: 1345-1357.
CrossRef - Shitsukawa, N., A. Takagishi, C. Ikari, S. Takumi and K. Murai, 2006. WFL, a wheat FLORICAULA/LEAFY ortholog, is associated with spikelet formation as lateral branch of the inflorescence meristem. Genes Genet. Syst., 81: 13-20.
CrossRef - Singh, R.K., A. Tapia-Santos, T.W. Bebee and D.S. Chandler, 2009. Conserved sequences in the final intron of MDM2 are essential for the regulation of alternative splicing of MDM2 in response to stress. Exp. Cell Res., 315: 3419-3432.
CrossRef - Takeuchi, M., T. Ueda, N. Yahara and A. Nakano, 2002. Arf1 GTPase plays roles in the protein traffic between the endoplasmic reticulum and the Golgi apparatus in tobacco and Arabidopsis cultured cells. Plant J., 31: 499-515.
CrossRef - Vitale, N., J. Moss and M. Vaughan, 1997. Interaction of the GTP-binding and GTPase-activating domains of ARD1 involves the effector region of the ADP-ribosylation factor domain. J. Biol. Chem., 272: 3897-3904.
CrossRef - Wang, J.R., Z.H. Yan, Y.M. Wei and Y.L. Zheng, 2004. A novel high-molecular-weight glutenin subunit gene Ee1. 5 from Elytrigia elongata (Host) Nevski. J. Cereal Sci., 40: 289-294.
Direct Link - Wang, X., X. Zhao, J. Zhu and W. Wu, 2006. Genome-wide investigation of intron length polymorphisms and their potential as molecular markers in rice (Oryza sativa L.). DNA Res., 12: 417-427.
CrossRef - Wang, Z. and C.B. Burge, 2008. Splicing regulation: From a parts list of regulatory elements to an integrated splicing code. RNA, 14: 802-813.
CrossRef - Yang, L., G. Jin, X. Zhao, Y. Zheng, Z. Xu and W. Wu, 2007. PIP: A database of potential intron polymorphism markers. Bioinformatics, 23: 2174-2177.
CrossRef - Yao, Y., Z. Ni, J. Du, Z. Han, Y. Chen, Q. Zhang and Q. Sun, 2009. Ectopic overexpression of wheat adenosine diphosphate-ribosylation factor, TaARF, increases growth rate in Arabidopsis. J. Integrat. Plant Biol., 51: 35-44.
CrossRef - Zhao, X., L. Yang, Y. Zheng, Z. Xu, W. Wu, 2009. Subspecies-specific intron length polymorphism markers reveal clear genetic differentiation in common wild rice (Oryza rufipogon L.) in relation to the domestication of cultivated rice (O. sativa L.). J. Genet. Genom., 36: 435-442.
CrossRef