
Kuldeep Dhama
Avian Diseases Section, Division of Pathology, Indian Veterinary Research Institute, Izatnagar, 243122, Bareilly, Uttar Pradesh, India
Shambhu Dayal Singh
Avian Diseases Section, Division of Pathology, Indian Veterinary Research Institute, Izatnagar, 243122, Bareilly, Uttar Pradesh, India
Rajamani Barathidasan
Avian Diseases Section, Division of Pathology, Indian Veterinary Research Institute, Izatnagar, 243122, Bareilly, Uttar Pradesh, India
P.A. Desingu
Avian Diseases Section, Division of Pathology, Indian Veterinary Research Institute, Izatnagar, 243122, Bareilly, Uttar Pradesh, India
Sandip Chakraborty
Department of Animal Resource Development, Pt. Nehru Complex, Agartala, 799001, Tripura, India
Ruchi Tiwari
Department of Veterinary Microbiology and Immunology, Uttar Pradesh Pandit Deen Dayal Upadhayay Pashu Chikitsa Vigyan Vishwavidhyalaya Evam Go-Anusandhan Sansthan (DUVASU), 281001, Mathura, Uttar Pradesh, India
M. Asok Kumar
Avian Diseases Section, Division of Pathology, Indian Veterinary Research Institute, Izatnagar, 243122, Bareilly, Uttar Pradesh, India
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 6 | Page No.: 751-767







ABSTRACT
Growth in poultry sector is being challenged due to increased incidence and re-emergence of diseases caused due to evolution of several viral pathogens and use of live vaccines. Piles of economic losses are encountered due to these diseases. Avian Infectious Bronchitis (IB), caused by Corona virus, is OIE-listed disease and characterized by respiratory, renal and urogenital involvements, causing high mortality. Economic losses are encountered due to loss of productive performance of both egg and meat-type chickens. Variant viruses evolve due to spontaneous mutations and recombinations, causing disease in vaccinated flocks of all ages. Serotyping and genotyping are the common methods of classification of IBV strains. The virus has 4 clusters, grouped into 7 serotypes and the most important strains are Massachusetts, Connecticut, Arkansas, Gray, Holte and Florida along with numerous others, distributed round the globe. Several conventional and molecular diagnostic methods have been described for the diagnosis of IB in chickens. 'All-in/all-out' operations of rearing along with good biosafety measures forms the basis of prevention, whereas vaccination forms the backbone of IB control programme. Both live and inactivated (oil emulsified) conventional vaccines are available. The new generation vaccines (recombinant and vector-based) developed against locally prevailing IBV strains may be more helpful and avoid the reversion of virulence in live vaccine viruses. The present review deals with all these perspectives of this important emerging poultry pathogen.
PDF Abstract XML References Citation
Received: June 26, 2013;
Accepted: December 19, 2013;
Published: January 29, 2014
How to cite this article
Kuldeep Dhama, Shambhu Dayal Singh, Rajamani Barathidasan, P.A. Desingu, Sandip Chakraborty, Ruchi Tiwari and M. Asok Kumar, 2014. Emergence of Avian Infectious Bronchitis Virus and its Variants Need Better Diagnosis, Prevention and Control Strategies: A Global Perspective. Pakistan Journal of Biological Sciences, 17: 751-767.
DOI: 10.3923/pjbs.2014.751.767
URL: https://scialert.net/abstract/?doi=pjbs.2014.751.767
DOI: 10.3923/pjbs.2014.751.767
URL: https://scialert.net/abstract/?doi=pjbs.2014.751.767
INTRODUCTION
The poultry farming in India was at its infancy during the 1960s which started as a small backyard venture. However, during the past three decades, the poultry farming has shown impressive performance powered by intensive system of rearing, scientific management, improved poultry breeds and ubiquitous use of vaccines. Poultry farming has developed into a vibrant, full-fledged organized commercial industry with the growth rate of 8-10% in egg production and 12-15 percent in meat production. As a result, India occupies 3rd, 4th and 6th rank in egg, layer and broiler production, respectively. However, presently the poultry sector faces challenges in several fronts. Increased incidence of disease outbreaks, re-emergence of various pathogens/diseases with higher virulence from time to time, evolution of variant strains due to recombination and simultaneous infection with multiple virus types and use of live vaccines are the reasons for such challenges. These diseases bring piles of economic loss in the form of inefficient feed conversion, decreased production, mortality, loss of market value, increased production cost etc (Morley and Thomson, 1984; Kataria et al., 2005; Rahul et al., 2005; Senthilkumar et al., 2003; Natesan et al., 2006; Bhatt et al., 2011; Dhama et al., 2008a, 2011a, 2013a; Singh et al., 2012; Gowthaman et al., 2012). One such common disease having all these features is avian Infectious Bronchitis (IB) (Hofstad, 1984; Ignjatovic and Sapats, 2000). The inclusion of this disease in the OIE (i.e., World Organization for Animal Health) list signifies its economic importance. It is a highly contagious and acute disease of chickens, caused by Corona virus (Cook, 2001; Lin et al., 2012). Avian Infectious Bronchitis Virus (IBV) is the causative agent. Infectious Bronchitis Virus is responsible for causing respiratory, renal and urogenital disease. Such disease is characterized by high mortality rates in affected flocks and severe economic losses due to production loss by reduction in weight gain by broilers and drop in egg qualityand production in laying poultry birds (Cavanagh, 2005, 2007; Almeida et al., 2012; Abro et al., 2012). Infectious bronchitis is prevalent worldwide, despite scientifically advanced poultry industry practices and availability of good vaccines and remains a global threat to poultry industry. It is possibly the most economically important viral respiratory disease of chickens in regions where there is no Highly Pathogenic Avian influenza Virus (HPAIV) or velogenic Newcastle disease virus. Antigenic shift and drift play important role in evolution of new strains. Infectious Bronchitis virus is known to affect multiple systems of the host involving lungs, oviduct and kidneys (Cavanagha and Gelb Jr., 2008; Cook et al., 2012; Jackwood, 2012). It causes respiratory problems, reduced performance, decreased hatchability and fertility, nephrosis and irreversible damage to reproductive organs (Cavanagh and Naqi, 2003; Jones, 2010). Vaccination is less successful because of continuous emergence of antigenic variance and less cross protection between the variance (Jackwood, 2012; Wang et al., 2012). Vaccination programmes should take into account the locally prevailing IBV strains to offer better protection against the field strains. Although the disease was first reported more than 80 years ago, control is yet to be complete (Cook, 2001).
The present review deals with various aspects of this important poultry virus having global significance and specifically describes the IB virus evolution and emerging variant strains, epidemiology and economic importance, the disease and pathogenesis. It also covers recent trends in diagnosis and vaccine development and appropriate prevention and control measures to be followed with an aim to combat IBV by designing/adapting effective disease control strategies. The review would be helpful for researchers and poultry producers for getting updated and comprehensive information about IB virus affecting the poultry industry.
METHODS
Infectious Bronchitis Virus (IBV), a Corona-like appearance-‘Crown virus’, is an enveloped, pleomorphic virus with a club-shape projection about 20 nm in length, approximately 120 nm in diameter, single stranded ribonucleic acid (ssRNA), 27.6 Kbp length genome with positive polarity (Jackwood, 2012). The virus is fairly labile and easily destroyed by disinfectants or organic solvents viz., ether and chloroform, sunlight, heat (56°C for 15 min), alkalis and other environmental factors (Van Roekel et al., 1941).
Classification and virus characteristics: Infectious Bronchitis Virus (IBV) belongs to order Nidovirales, family Coronaviridae, subfamily Coronavirinae, genus Gammacoronavirus and species Avian corona virus (ICTV, 2011). Among RNA viruses, the Corona virus has the largest genome, consisting of a 27.7 kb, single-stranded, positive-sense RNA (Kuo et al., 2013). IBV viral genome codes four major structural proteins: the Spike (S), Membrane (M), the internal nucleoprotein (N) and small membrane protein (E). Protective immunity, Hemagglutination-Inhibition (HI) and most of the Virus Neutralizing (VN) antibodies are induced by S1 protein that is formed by post translational cleavage of S protein. S1, the major antigen to which neutralizing antibodies develop, is the determinant of host species specificity and pathogenicity by determining susceptible cell range (tissue tropism) within a host. The N-terminal (S1) part of the S protein mediates attachment to cells via., Receptor Binding Domain (RBD) and is the most variable part of the S protein having unique amino acid sequences determining the serotype. Infectious Bronchitis virus serotypes (old and new) differ from one another, as well as from vaccine strains by approximately 50% of S1 amino acid sequence (Casais et al., 2003). The C-terminal S2 part triggers fusion of the virus envelope with host cell membranes. Important mechanisms of viral variation and diversity largely depend upon the large size of RNA genome, permitting diverse means of mutations and recombinations (Lai, 1996). At present, there is no definitive classification system. Serotypes are typed based on Virus Neutralization Test (VNT) after isolating them in embryonated eggs or cell culture system or in Tracheal Organ Cultures (TOCs), whereas protectotype of a strain is typed based on Cross-Immunization Study (CIS). Strains that induce protection against each other increase the efficacy of vaccination. Genotypes are grouped into strains based on genetic characterization using Reverse-Transcriptase-Polymerase Chain Reaction (RT-PCR), followed by Restriction Endonuclease (RE) cleavage site analysis and sequencing (Callison et al., 2006). Virus has the ability to mutate or change its genetic makeup very quickly. Numerous serotypes of IBV have emerged which complicate the control efforts through vaccination.
Replication strategies: The multifactorial regulation of translation during a IBV infection can be due to the N protein, causing down-regulation of host cell translation in IBV infected cells (Hilton et al., 1986; Siddell et al., 1981), with upregulation of the translation of virus-encoded proteins (Tahara et al., 1994). The N protein may interfere with host cell translation by disrupting the formation of new ribosomes and possibly the cell cycle in the nucleolus, followed by binding to the 5' end of virus-derived RNA (Nelson et al., 2000). This causes recruitment of ribosomes for translation of viral RNAs. Alternatively, interaction of ribosomes with the Corona virus core can cause destabilization and release of genomic RNA (Hiscox and Ball, 1997), thereby causing trafficking of virus particles to the nucleolus in association with ribosomal proteins.
Evolution of IBV: The evolution of IB virus is constantly being reported due to intensive poultry farming practices, rapidly growing poultry industry, increased global trading and immense pressure of vaccines (Abro et al., 2012). New variants of IBV have emerged due to spontaneous mutation and recombination during virus replication, followed by replication of those phenotypes favored by selection (Abdel-Moneim et al., 2012; He et al., 2012; Toro et al., 2012a, b; Liu et al., 2013b; Mo et al., 2013) and cause significant disease in vaccinated flocks of all ages. It is assumed that due to widespread vaccination, the immune selection pressure involving the S1 subunit of S gene and the high mutation rate of the viral genome together might have resulted in the emergence of many serotypes and variants (Abro et al., 2012). Existing IB vaccines do not provide protection against these (Kuo et al., 2010). A new live-attenuated vaccine is required mainly based on locally prevalent field strains. Important IBV variants are: D274 and D1466 (Netherlands), B1648 (Belgium), AZ20/97 (Italy), Arkansas (USA), 84084 and 88121(France), 4/91(UK). Different environmental determinants within the host viz., immune responses, affinity for cell receptors, physical and biochemical conditions are also implicated in the selection process (Toro et al., 2012a). Recombinations in IBV S1 and N genes have been documented in several field isolates (Cavanagh, 2007; Kuo et al., 2010, 2013; Abdel-Moneim et al., 2012; Liu et al., 2013a). Variants may attain increased virulence, efficient receptor binding, rapid transmission and survival in host system. It is important to isolate and type the IBV virus strains prevailing in a geographical area time to time and vaccinate birds accordingly with vaccine strains that offer maximum cross protection against the field virus in question or use combination of different strains to provide broad protection. Similarly, vaccines can be developed from recently isolated IBV field strain(s) from a particular region. Amino acid substitutions in epitopes responsible for development of neutralizing antibodies can result in immune evasion (Shi et al., 2006). Thus, nucleotide sequencing and identification of amino acid substitutions especially involving S1 and N gene help to identify vaccine failure due to antigenic variation (Kuo et al., 2013). Variants are generally related to the appearance of positively selected, single point mutations or recombination in the antigenic domain of the viral proteins which occurs continuously (Lee and Jackwood, 2001; Jackwood et al., 2005; Kuo et al., 2013). These mutations lead to the alteration of virulence and the escape of the viruses from host defenses (Lee and Jackwood, 2001). The average synonymous mutation rate in all coronaviruses including IBV is approximately 1.2x10-3 substitutions/site/year (Hanada et al., 2004; Holmes, 2009). The use of modified live vaccines has helped in rapid evolutionary rate of IBVs (Jackwood et al., 2012). Understanding the evolution of IBV is essential with regard to development of control and prevention strategies.
Classification of strains: Serotyping by Virus Neutralization (VN) and Haemagglutination Inhibition (HI) and genotyping are the common methods of classification of IBV strains. Serotype specific monoclonal antibodies are induced by S protein (S1 subunit) but are available for a few serotypes in a small quantity. Owing to the emergence of novel variants it is difficult to have serum against all the strains. New serotype may emerge due to mutation. Nowadays, to classify (genotyping) strains, S1-specific RT-PCR is followed by sequencing or restriction endonuclease analysis (Abdel-Moneim et al., 2006; Sumi et al., 2012). Deduced S1 amino acid sequences with Virus Neutralization (VN) tests have revealed many serotypes. These serotypes differ by about 20-25% (amino acid), or by 40% or more occasionally, example being 7.6% difference between Conn 46 and Mass 41 strains in the S1 region or amino acid identity (97%) with D274.4 clusters, grouped into 7 serotypes based on RT-PCR analysis of the N gene and neutralization tests, respectively, were also identified (Wang et al., 1994; Roussan et al., 2009).
The Connecticut (Conn) isolate neither cross neutralize nor cross protect against Massachusetts (Mass) isolate (Jungherr et al., 1956). Florida, Clark 333 and Arkansas were other variants identified along with Holte and Gray (Winterfield and Hitchner, 1962; Brown et al., 1987; Butcher et al., 1989; Kinde et al., 1991). In North America, Massachusetts, Connecticut and Arkansas 99 IB viruses are common serotypes. A large number of variant viruses (82) have been reported, however only GAV and GA98 were found to be implicated in widespread disease disseminations and persistent virus infections. Other IBV variants involved were Mass, Conn, Ark-DPI, CAV, DE072, MX97-8147, etc. (Jackwood et al., 2005). Three variants viz., CA557/03, CA706/03 and CA1737/04 were not related to each other or to Conn, Ark, or Mass vaccine strains, genetically or by cross-virus neutralization test (Ziegler et al., 2002; Kingham et al., 2000; Jackwood et al., 2007). Most of the Brazilian IBV field isolates recorded up to 1989 were classified as Mass (Massachusetts) serotype. Seroprevalence of the IBV in various species of birds in some of the countries (like Grenada) was reported, although vaccination against IBV is not practiced (Gutierrez-Ruiz, 2004; Sabarinath et al., 2011; Ramirez-Gonzalez et al., 2012). There are five distinct genotypes: A, B, C, D and Massachusetts based on the RFLP patterns of Twelve Brazilian isolates and one reference vaccine strain (De Fatima et al., 2008; Rimondi et al., 2009; Acevedo et al., 2012). Massachusetts-serotype vaccine is the only one type of live attenuated vaccine approved in Brazil, but the genotypes are greatly divergent and the most of them belongs to Non-Massachusetts types (Hipolito, 1957; Di Fabio et al., 2000; Villarreal et al., 2007, 2010; Brandao, 2010). Interestingly, in Argentina, putative genotypes have been observed (De Fatima et al., 2008; Rimondi et al., 2009; Acevedo et al., 2012). The Massachusetts type was first reported in 1948 in UK and subsequently in Holland new serotypes were reported, against which vaccines were developed (Kusters et al., 1987; Parsons et al., 1992; Cook et al., 1996).
In African continent, IB was observed from 1954 onwards (El Houadfi and Jones, 1985; Kelly et al., 1994; Thekisoe et al., 2003; Owoade et al., 2004, 2006; Ducatez et al., 2009; Mushi et al., 2006). Five field IBV genotypes with three new IBV genotypes different from vaccine strains Massachusetts type, the fourth genotype similar to vaccine strain MA5 (Massachusetts type) and the fifth genotype similar to vaccine strain 4/91 have been identified (El Bouqdaoui et al., 2005; Bourogaa et al., 2009, 2012; Susan et al., 2010). The Mass and H120 serotype vaccine isolates and D274 are the most common IBV strains found to be circulating in the Middle East (Roussan et al., 2009). Others variants reported were IBV variant 1 (IS/64714/96, IS/222/96, IS/251/96, 793/B) and variant 2 strains (IS/223/96, IS/572/98, IS/585/98, IS/589/98) (Callison et al., 2001; Meir et al., 2004; Mahmood et al., 2011; Ababneh et al., 2012). Mass, Ark, DE-072 and JMK serotypes of IBV have been documented, based on the results from the Hemagglutination Inhibition (HI) test (Gharaibeh, 2007) and D274 and 4/91 (793B) with the use of RT-PCR (Roussan et al., 2008). Variant viruses in Russia clustered into the six novel genotypes (Bochkov et al., 2006).
The Chinese isolates were separated into five genetic groups (genotypes) viz. (LX4, 4/91, JP, Gray and Mass) and seven serotypes based on S1 HVR I nucleotide sequences and virus-neutralization test, respectively (Chen et al., 2009; Li et al., 2012). Thirteen isolates belong to serotype I; two isolates, GX-NN10 and GX-YL2, vaccine strains H120 and Ma5 and one reference strain, M41, belong to serotype II; five isolates belong to serotype III; isolate GX-YL1 belongs to serotype IV; isolates GX-GL1 and GX-NN7 as well as one vaccine strain, 4/91, belonged to serotype V; isolates GX-YL8 and GX-YL9 belonged to serotype VI. Neutralization titers of isolate GX-NN12 were relatively low against all the antisera, so it belongs to serotype VII (Wang et al., 1997; Wu et al., 1998; Li and Yang, 2001; Yu et al., 2001a; Liu and Kong, 2004). In Thailand, analysis of S1 HVR I nucleotide sequences separated the viruses into two groups (I and II) (Pohuang et al., 2009). The sequence analysis of the S1 gene demonstrates that NIB Indian isolate PDRC/Pune/Ind/1/00 possesses a unique genotype compared to other reference strains of various countries and is unrelated to North American, European and Australian strains (Bayry et al., 2005). Recently, an Indian IBV strain (India/LKW/56/IVRI/08) revealed 99% homology with a Thailand strain (THA280252) and the other strain (India/NMK/72/IVRI/10) showed even greater homology with IBV strains from UK (4/91 pathogenic strain), Japan (JP/Wakayama/2/2004) and China (TA03) (Sumi et al., 2012).
The disease known as uraemia was first recognized in Australia in the late 1940s. Cumming (1963) isolated Nl/62 virus (synonym 'T') from one such case in 1962, a prototype of nephropathogenic strains of IBV (Cumming, 1963). They were found to differ antigenically from other IBV strains having no common epitopes on either the N or M proteins (Ignjatovic and McWaters, 1991). These findings have indicated that unusual changes are occurring in strains isolated in Australia. Infectious Bronchitis virus strains viz., Ql/88, Ql/89, N3/88, N6/88, Nl/89, N2/89, N2/90, N5/90, Nl/94, N6/94, V18/91, V19/91, V6/92, V9/92, Vl/93, V2/93 and V3/93, shared only minor antigenic similarity with the classical Infectious Bronchitis virus strains and clearly belonged to group 2 of novel strains. There is no clear-cut correlation between the S1 amino acid sequences and the nephropathogenicity of strains (Ignjatovic and McWaters, 1991; Sapats et al., 1996; Ignjatovic et al., 1997). In New Zealand, IBV was first isolated in 1967 and four serotypes (A, B, C and D) of the virus were described in 1976 that were different from those present in other countries, based on virus neutralization tests. Subsequently, a vaccine was produced from one of those serotypes (A) which was said to protect against all serotypes and is currently used in layer and breeder flocks and are having similarity to Australian Vic S strain (Pohl, 1967; Lohr, 1976; McFarlane and Verma, 2008). Percent cross protection offered by commonlu employed vaccine strains against various known field strains has been described by Gelb et al. (1991). Infectious Bronchitis virus strains prevailing worldwide are listed in Table 1. Emergence of IBV strains and timeline of events in their evolution are depicted in Fig. 1.
Host range, age susceptibility and tissue tropisms: Host range of IBV has been enlarging. The domestic chicken has been widely regarded as the exclusive host for IBV. Infectious bronchitis virus is also reported in pheasants, racing pigeons, peacock, partridge and mallard and associated with respiratory disease, egg production and shell quality disturbance. Infectious bronchitis virus grows well in 9-12 days old embryonated chicken eggs via., intra-allantoic route, producing curling and dwarfing of embryo. Infectious bronchitis virus has been propagated in chicken embryo tissue (kidney, lungs, liver), embryonic turkey kidney, Vero cells and chicken tracheal organ culture (Meir et al., 2004). Among all the age groups of birds the young chicks are most ill affected and resistance develops according to age. Female chicks may harbor certain genotypes of the virus in the oviductal epithelium due to lack of maternal antibodies and such hens are termed as ‘false layers’; the phenomenon being recently observed w ith a new variant predominant in Europe. Age-related susceptibility to infections is suspected to be under the control of an immunological response which in chicks develops fully at 2-5 weeks of age. Serotype-Massachusetts and Connecticut show more affinity for respiratory tract, whereas, serotypes-Holte, Gray and Australian ‘T’ strain are associated with nephrosis (Capua et al., 1999; Ignjatovic et al., 2002). Under certain circumstances, IBV is thought to persist for a considerable time in infected birds at some privileged sites like kidney and caecal tonsils (Meulemans and van den Berg, 1998; Almeida et al., 2012).
EPIDEMIOLOGY AND ECONOMIC IMPORTANCE
Infectious bronchitis is prevalent in intensive poultry production system, having a high incidence of infection with significant economic loss to the world poultry sector, despite proper vaccination.
| Table 1: | List of the strains of IBV prevalent in different continents across the world |
 | |
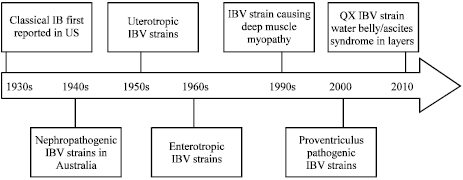 | |
| Fig. 1: | Chronological enlistment of emerging IBV stains isolated from certain continents from different affected organs |
The virus has got different serotypes, prominent ones being Massachusetts type (M41), the prototype IBV, first isolated in 1941 and Connecticut (Conn strain) (Jungherr et al., 1956). Reports regarding IB have since been given from different parts of the globe in several poultry raising areas caused by unique strains except Australia and New Zealand. More than 30 serotypes and dozens of virulent variants have also evolved at times (Zanella et al., 2003). The incidence of nephrosis that is caused by variants of IBV is on the increase in many countries which necessitates the development of new vaccines (Winterfield and Albassam, 1984). Outbreaks of IB are often due to infections with the strains serologically different from the strains used as vaccines. Outbreaks mainly occur during winter (Lopez and McFarlane, 2006). The seasonal use of live virus vaccine (Ark) has led a subpopulation of the vaccine to revert back to virulence. Losses from production inefficiencies are more than mortality. In broilers IB leads to poor weight gain and loss of profit at slaughter. In layers, IB causes loss of egg qualities and egg production may drop down to 10-50%. Nephropathogenic strains cause mortality of up to 30% in susceptible flocks. In United Kingdom (U.K.), IBV is the biggest single cause of infectious disease-related economic loss (Pennycott, 2000; Meulemans et al., 2001).
Transmission and spread: Coughing and sneezing are mainly responsible for spread of the virus in chicken flocks. A short incubation period of 18-36 h is observed after IBV infection and thereafter clinical signs develop. Carrier birds rather than vectors are the nidus of infection. Disease can be transmitted through infected semen (Gallardo et al., 2011). There is no vertical transmission, ruling out the chances of infection related to management of hatcheries. Transmission from farm to farm is related to movement of contaminated people, equipment and vehicles. During an active outbreak air-borne spread of the virus over a considerable distance occurs. It is assumed that poultry industry in different parts of the world can contract the disease via migratory birds, even though the role of wild birds in the spread of IBV in most of the instances is speculative and largely unknown, deserving more attention and research activities (Sjaak de Wit et al., 2011).
Pathogenesis and the disease: Infectious Bronchitis virus has wide tissue tropism including respiratory, urogenital and digestive system (Boroomand et al., 2012) and has pathogenic effects in tissues of respiratory tract, kidney and oviduct. Infection occurs via respiratory route regardless of tissue tropism of strains. Virus multiplies chiefly in upper respiratory tract, following which viraemia occurs and the virus gets widely disseminated to other tissues. After the acute phase of infection, a persistent infection can be established in specific organs and the virus can be excreted continuously. The virus is primarily epitheliotropic and replicates in many epithelial cells, including those of respiratory tract, kidney and gonads producing lesions and in alimentary tract, many times with little pathobiological clinical effect (Ignjatovic et al., 2002). Infectious bronchitis virus infection of female chicks less than 2 weeks of age can cause permanent damage. Respiratory-urinary organs-digestive tracts-generative organ based transfer of the virus is responsible for tissue tropism (Benyeda et al., 2009). Infectious bronchitis occurs in birds of all age groups, but severely affects chicks evincing malaise, depression and retarded growth. Young birds of less than 3 weeks of age are more susceptible than older ones. The age and immune level of the poultry flocks and the pathogenic potential of the causative viral strain affects the nature and disease severity. Signs include respiratory symptoms, decreased egg production due to permanent damage to the oviduct and deterioration in egg qualities like watery albumin, misshapened and soft-shelled eggs. Recovery from the typical respiratory phase may occur in birds infected with nephropathogenic viruses which subsequently show signs of depression, ruffled feathers and wet droppings along with increased water intake (Lee et al., 2004). Mortality, as high as 25% or more occurs in chicken above 6 weeks of age group.
Respiratory tract shows serous, catarrhal, or caseous exudates in the nasal passages, sinuses and trachea. Caseous plugs may be found in the lower trachea or bronchi of young birds. Air sac affections are characterized by thickening and opacity. Focal areas of pneumonia may appear. In urogenital system, middle third of the oviduct is most severely affected and may be non patent and hypo-glandula, continuous patent but underdeveloped structure to a blind sac. Swollen and pale kidneys with the tubules and ureters often distended with urates. Despite lack of gross lesions in kidney, microscopic changes of nephritis may still be present (Abdel-Moneim et al., 2005). The abdominal cavity of chickens in production may contain yolk substances, a condition also observed in diseases leading to marked fall in egg production (Ahmed et al., 2007). Pathology is not usually associated with infection of the alimentary tract by enterotropic IBV. Pathogenic strains cause thickened, haemorrhagic or ulcerative lesion in the proventriculus, haemorrhagic lesions in the caecal tonsils and thickening of duodenum (Escorcia et al., 2002).
DIAGNOSIS
Infectious bronchitis can be diagnosed on the basis of clinical manifestation of disease, rising antibody titres and detection of antigen and/viral DNA in the tissue sections and clinical material. Infectious bronchitis virus can be isolated from the trachea, lungs and kidneys of infected chickens (Pradhan et al., 1982; Verma and Malik, 1971; Sylvester et al., 2003a; Dhama et al., 2011a, b) while liver and pancreas are candidates for antigen detection; trachea and spleen for histological diagnosis (Fan et al., 2012). Two or three blind passages may be required for primary virus isolation which can be a cumbersome and time taking process. Therefore, embryos of Specific Pathogen Free (SPF) chicken or their Tracheal Organ Cultures (TOCs) are preferred for isolation of IB virus (Cook et al., 1976). Further, direct detection methods in infected tissues include immunohistochemistry (Nakamura et al., 1991; Chen et al., 1996) or in situ hybridization (Collisson et al., 1990). Confocal microscopy can be used to detect the viral N protein both from healthy and uninfected cells and also reveals the presence of this important protein (required in viral replication cycle) in the host cell (Hiscox et al., 2001). Serological test include group specific Enzyme Linked Immune-Sorbent Assay (ELISA) (including monoclonal antibody based technique) (Lin et al., 2012), Virus Neutralization Test (VNT) (type specific) and Haemagglutination Inhibition (HI), Indirect Immunofluorescence Antibody Test (IFAT) and agar gel immunodiffusion (AGID). Fluorescent Antibody Technique (FAT) is also a good test for antigen detection. Nucleocapsid phosphoprotein gene-specific reverse transcription loop-mediated isothermal amplification (RT-LAMP) assay has also been developed. However, for detection as well as serotyping, VNT is the gold standard test of choice while cross neutralization tests are used for detection of the variants (Elankumaran et al., 1999; De Wit, 2000; Chen et al., 2010). Tests involving detection of antigens should take into consideration that live IB vaccines are used ubiquitously and antibodies may fail to detect variant strains with modifies epitopes. Confirmation is made by nucleic acid-based methods: Reverse Transcription-Polymerase Chain Reaction (RT-PCR) using-specific oligonucleotide primers, producing DNA copies of the S1 part of the spike glycoprotein gene to determine the identity of a field strain; Restriction Fragment Length Polymorphism (RFLP) RT-PCR and sequencing can identify all known serotypes of IBV as well as variant viruses (Meir et al., 1998, Sylvester et al., 2003b, 2006; Almeida et al., 2012; Jackwood et al., 2012; Sumi et al., 2012). Reverse Transcription-Polymerase Chain Reaction product is digested with a set of specific restriction endonucleases; the generated fragments are separated by gel electrophoresis and the specific pattern of their separation in the gel is compared with those of the standard strains for identification. Genotype identification is achieved by sequencing of the S protein (S1 subunit) gene. In US and Israel, RFLP analysis and RT-PCR product cycle sequencing are being used to identify filed strains (Zwaagstra et al., 1992). Recently, the improvements in the PCR technique have created way for the application of real time-PCR for very rapid detection and quantification of virus (Jackwood et al., 2003); multiplex-PCR and multiplex nested RT-PCR technique for differential diagnosis of IB infection (Kataria et al., 2005; Dhama and Mahendran, 2008; Chen and Wang, 2010; Nguyen et al., 2013). RNase T1 fingerprinting analysis can be used for fingerprinting of the IBV genome, advantage being use of complete genome for generation of strain-specific fingerprints (De Witt, 2000; Alvarado et al., 2005; Dhama et al., 2011b).
Infectious bronchitis virus lesions and PCR based diagnosis are presented in Fig. 2.
DIFFERENTIAL DIAGNOSIS
Infectious Bronchitis may resemble other acute respiratory diseases viz., Newcastle Disease (ND), infectious laryngotracheitis (ILT) and Infectious Coryza (IC) that occur more severely and causes greater production losses when compared to IB. Moreover, nervous symptoms of ND, slow nature of spread of ILT, or facial swelling of Infectious coryza are absent in case of IB. Production declines and shell quality problems in flocks infected with the Egg Drop Syndrome (EDS) are similar to those seen with IB, except that internal egg quality is not affected in case of EDS. Nutritional deficiency disorders must also be taken into consideration while diagnosing IBV (Sylvester et al., 2005; Cavanagha and Gelb Jr., 2008).
PREVENTION AND CONTROL
Biosecurity for prevention of the infectious bronchitis virus: Prevention and control measures mainly include follow up of strict biosecurity, good hygiene and sanitation practices, along with judicious vaccination programme. A one-age system ('all-in/all-out' operations) of rearing, cleaning and disinfection between batches will reduce the level of infection. Good management practices comprise of strict isolation/quarantine, restocking/repopulation with disease free day old chicks and adapting appropriate cleaning, disinfection and hygienic measures in the poultry farm.
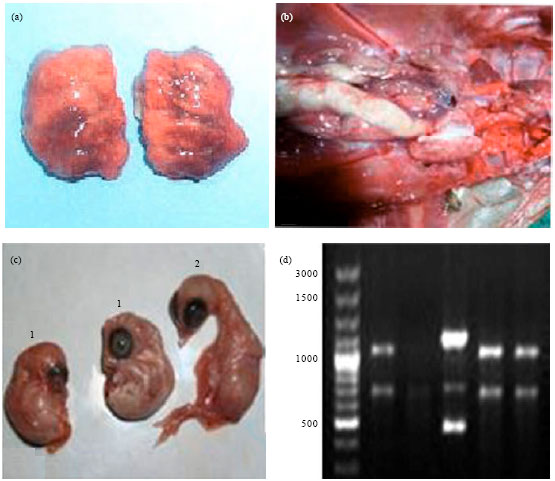 | |
| Fig. 2(a-d): | IBV lesions and PCR based diagnosis (a) Lungs of experimentally infected chicks showing areas of consolidation, (b) Swollen ureter distended with urates in a chicken naturally infected with nephropathogenic IBV, (c) IBV infected chicken embryos (1) Exhibiting dwarfing and curling compared with uninfected embryo (2) and (d) RFLP pattern of PCR-amplified S1 gene (1720 bp) of vaccine (lane 1, 2) and field (lane 3, 4, 5) IB viruses |
Steps to minimize the intensity of infectious virus and limiting its introduction in poultry houses include controlled visitor’s access of the farm premises; keeping separate clothing, footwear and equipments for each farm/unit; controlling movement of farm workers/personnel and equipments between farms and keeping appropriate footbaths with disinfectants at the entry points. These check points are very crucial for prevention and control of IBV on multi-age sites/farms and at times require a challenging job to be followed. All organic material should be removed or/and disposed from the house (top 4-5 cm of soil also removed from earth floor) and cleaning the houses at 35-55 Bar water pressure (adding detergents in the cleaning process is also advisable). Even though IBV is easily killed, use of suitable disinfectants (formaldehyde or chlorine releasing, quaternary ammonium compounds) with appropriate concentration and an optimum working time is very crucial to minimize infectivity of any residual virus particles. Disinfectants can be very helpful for preventing virus infections in a farm. Successive chicken flocks must be restocked with a minimum of 10-14 days downtime between them (Welchman et al., 2002; Sylvester et al., 2005; De wit et al., 2010; Dhama et al., 2011a).
VACCINATION
Vaccination is the main key for the prevention and control of infectious bronchitis virus. In most of the countries, day-old chicks are being vaccinated in hatcheries itself with low virulence IBV vaccines. Thereafter, a booster immunization is followed with virulent vaccines, usually in the drinking water. This low virulence is suitable for chicks with a lower level of maternal immunity and they do not cause the respiratory reactions which can occur with vaccines of higher virulence; disadvantage being the low level of immunity only enough to protect the respiratory tract (Kataria et al., 2005). Two types of vaccine viz., live attenuated and inactivated and killed are usually formulated in an oil emulsion adjuvant. Live vaccines are used in broilers and for the initial vaccination of breeders and layers while inactivated (oil emulsified) vaccines are administered in breeders and layers mainly during laying (Ladman et al., 2002; Jackwood et al., 2009). Nevertheless, for inactivated vaccines to be effective birds should have been previously "primed" with a live vaccine. Serial passages of IBV strains is followed in Embryonated Chicken Eggs (ECE) in order to achieve the required level of attenuation so as to be used in live vaccines. With optimal conditions, vaccination may give immunity for many months and this may be life-long (Cook, 2001; Bijlenga et al., 2004).
Live vaccines: These are Massachusetts strains: H120 vaccine (the most common representative of live vaccines) which is a mild vaccine and is known for the typical level of attenuation due to the number of passages it has undergone. It is usually used for vaccinations for the first time without inducing a long lasting immunity, in areas where there is increased level of field challenge with the intention of keeping the local protection of the respiratory tract at high level. Initial vaccination can be done individually by eye drop, intra tracheal or intranasal route or by mass vaccination (e.g., coarse spray or drinking water). Such procedure is usually inexpensive; induce local as well as systemic immunity. But unfortunately it can cause some vaccination reaction observed for a few days after vaccine application (Matthijs et al., 2003; Bijlenga et al., 2004). Ma5, a single component vaccine, is a mild one, used as a single component and can also be included in first vaccination programs with IB 4/91 vaccines and inactivated vaccines for broad protection against different IBV serotypes. Live vaccines are generally used in breeders and layers (young birds) to keep a good level of local protection of the respiratory tract and are advisable in areas of increased level of field challenge. However, selection of vaccine strain must be based on the strains prevalent in the area/country. Different vaccine strains in SPF chickens provides cross-protection against homologous and reference strains and variant field isolates (De Wit and van de Sande, 2009). Higher levels of cross-protection to some heterologous strains are given by combination of Mass and Conn or Mass and JMK. The occurrence of multiple serotypes of the virus has complicated and increased the cost of disease prevention and warrants the use of local strains in vaccines for their effective control. The inactivated vaccines are used primarily at the point of lay to avoid stress and loss of production. The Massachusetts (Mass or M41) strain is the most popular one as it is the representative of the initial isolates reported from many countries (Gelb et al., 1989, 1991, 2005; Cook et al., 1999; Terregino et al., 2008). In order to give specific protection against the IBV type, IB 4/91 variant virus (containing a strain of the 4/91serotype) or IB 274 vaccine virus (containing a strain of the D207(D274) serotype) are used; when combined with Ma5 and IB multi vaccines, they provide broad protection (Mase et al., 2008).
Inactivated vaccines: Inactivated vaccines induce long lasting immunity, show no vaccination reactions, usually cost more than live vaccines and combination of different antigens apart from IBV can be achieved when given individually. Elevated levels of circulating antibodies were stimulated by inactivated vaccines than live vaccines, therefore it is useful in a breeder program where maternal antibody protection is needed. Still, due to induction of better T cell responses and rendering a higher local antibody (IgA) stimulation, modified live vaccines play significant role in protecting commercial birds (layers). Chickens must be properly primed with live vaccines, in order to exploit the potential of the inactivated vaccine and by this way, highest titres will be obtained at an interval of at least 4-6 weeks (period of vaccination between last live and inactivated vaccine) (Ladman et al., 2002). Further, vaccination programs may be simplified by combining inactivated antigens against two or more serotypes (or two or more diseases) into one vaccine (Hong et al., 2012).
IB vaccines employed in various countries: Both modified live vaccines and inactivated water-in-oil emulsions are available for the Massachusetts, Connecticut and Arkansas serotypes in North America; California strains and Georgia 98 vaccines being used in USA. In Europe, "Holland variants" commonly designated by number (e.g., D-274, D-1466) are documented; IB H120 based vaccine being used in most parts of Europe. IB Ma5 and IB 4/91 (as mentioned earlier) are live, freeze-dried vaccine serotypes, giving long lasting protection. Infectious Bronchitis D274 is a live vaccine against strain D274 in poultry for vaccination of future breeding and layers stock. Vic S vaccine is used in most vaccination programs in Australia. The K2 vaccine might be useful for the control of newly evolving IBV recombinants (new cluster 1) and variants (new cluster 2) in Korea (Lim et al., 2012).
Future vaccines: With advances in molecular biology, novel vaccines like DNA vaccines, sub-unit vaccine and vectored vaccine using S1 glycoprotein gene, along with reverse genetics vaccines have been tried in different laboratories (Dhama et al., 2008b). Use of spike protein based DNA vaccine has revolutionized the concept of immunization against IB (Sylvester et al., 2005). Such tailor made-vaccines can be designed to suit the locally prevailing IBV strains and the problem of live attenuated strains reverting to virulence can be avoided. The recombinant or vector-based vaccines are also designed to introduce antigens from two or more viruses which act as multivalent vaccine giving protection against two or more diseases. DNA vaccine is another promising area as shown in initial clinical trials. These new generation vaccines can be administered safely in-ovo or to live chickens. The efficacy of these vaccines needs to be tested in large scale experiments before introduced for commercial purpose (Boots et al., 1992; Yu et al., 2001b).
It is the need of the hour to combat this economically important and emerging poultry pathogen by adapting and developing newer diagnostics, effective and safer vaccines, exploring novel therapeutics and following appropriate prevention and control strategies (Kataria et al., 2005; Sylvester et al., 2005; Cavanagh, 2007; Cavanagha and Gelb Jr., 2008; Dhama and Mahendran, 2008; Cook et al., 2012; Jones, 2010; Sjaak de Wit et al., 2011; Jackwood, 2012; Mahima et al., 2012; Deb et al., 2013; Dhama et al., 2008b, 2011a, b, c, 2013b, c, d, e, f, g, h; Tiwari et al., 2013).
CONCLUSION AND FUTURE PERSPECTIVES
Infectious bronchitis is an economically important disease of poultry having worldwide distribution, causing high morbidity (80%) and low mortality (20%). The economic losses are incurred upon by reduced egg production as well as worsen egg quality (internal and external); presence of silent layers; infertility and delay in growth; and increase in the susceptibility to secondary infections which altogether show the importance of the disease. Little or only partial cross-protection occurs between vaccine strains and new field strains which necessitate the development of new vaccines. The disease possesses a continuous threat to poultry despite of good vaccines, due to emergence of new IBV variants from time to time for which disease control by improved biosecurity, hygienic measures and vaccination programmes are required. For devising a strong IB control program, the presence of several serotypes, strains or variants of IBV and the possibilities of the emergence of novel ones have to be monitored. In this regard, detection and control strategies based on molecular biology and biotechnological tools and techniques are required to be developed. Alongside, upgradation of conventional techniques is also required to tackle the growing threat of IBV infection in order to effectively prevent and control it in the years to come. However, currently, better diagnostics and new generation vaccines are being developed along with the application of in ovo vaccination strategies so as to effectively check the spread of this ubiquitous and highly contagious viral infection of domestic birds.
REFERENCES
- Owoade, A.A., M.F. Ducatez and C.P. Muller, 2006. Seroprevalence of avian influenza virus, infectious bronchitis virus, reovirus, avian pneumovirus, infectious laryngotracheitis virus and avian leukosis virus in Nigerian poultry. Avian Dis., 50: 222-227.
PubMed - Ababneh, M., A.E. Dalab, S. Alsaad and M. Al-Zghoul, 2012. Presence of infectious bronchitis virus strain CK/CH/LDL/97I in the Middle East. ISRN Vet. Sci., Vol. 2012.
CrossRef - Abdel-Moneim, A.S., M.A. Afifi and M.F. El-Kady, 2012. Emergence of a novel genotype of avian infectious Bronchitis virus in Egypt. Arch. Virol., 157: 2453-2457.
CrossRefDirect Link - Abdel-Moneim, A.S., M.F. El-Kady, B.S. Ladman and J. Gelb, 2006. S1 gene sequence analysis of a nephropathogenic strain of avian infectious Bronchitis virus in Egypt. Virol. J., Vol. 3.
CrossRef - Abro, S.H., L.H.M. Renstrom, K. Ullman, M. Isaksson and S. Zohari et al., 2012. Emergence of novel strains of avian infectious bronchitis virus in Sweden. Vet. Microbiol., 155: 237-246.
CrossRef - Acevedo, A.M., H. Diaz de Arce, P.E. Brandao, M. Colas, S. Oliveira and L.J. Perez, 2012. First evidence of the emergence of novel putative infectious bronchitis virus genotypes in Cuba. Res. Vet. Sci., 93: 1046-1049.
CrossRef - Ahmed, Z., K. Naeem and A. Hameed, 2007. Detection and seroprevalence of infectious bronchitis virus strains in commercial poultry in Pakistan. Poult. Sci., 86: 1329-1335.
CrossRefDirect Link - Almeida, D.O., R. Tortelly, E.R. Nascimento, M.A. Chagas, M.I. Khan and V.L. Pereira, 2012. Avian infectious bronchitis and deep pectoral myopathy-a case control study. Poult Sci., 91: 3052-3056.
CrossRef - Alvarado, I.R., P. Villegas, N. Mossos and M.W. Jackwood, 2005. Molecular characterization of avian infectious bronchitis virus strains isolated in Colombia during 2003. Avian Dis., 49: 494-499.
PubMedDirect Link - Bayry, J., M.S. Goudar, P.K. Nighot, S.G. Kshirsagar and B.S. Ladman et al., 2005. Emergence of a nephropathogenic avian infectious bronchitis virus with a novel Genotype in India. J. Clin. Microbiol., 43: 916-918.
CrossRefDirect Link - Benyeda, Z., T. Mato, T. Suveges, E. Szabo and V. Kardi et al., 2009. Comparison of the pathogenicity of QX-like, M41 and 793/B infectious bronchitis strains from different pathological conditions. Avian Pathol., 38: 449-456.
CrossRef - Bijlenga, G., J.K.A. Cook, J. Gelb and J.J. de Wit, 2004. Development and use of the H strain of avian infectious bronchitis virus from the Netherlands as a vaccine: A review. Avian Pathol., 33: 550-557.
CrossRef - Bochkov, Y.A., G.V. Batchenko, L.O. Shcherbakova, A.V. Borisov and V.V. Drygin, 2006. Molecular epizootiology of avian infectious Bronchitis in Russia. Avian Pathol., 35: 379-393.
CrossRefDirect Link - Boots, A.M.H., B.J. Benaissa-Trouw, W. Hesselink, E. Rijke, C. Schrier and E.J. Hensen, 1992. Induction of anti-viral immune responses by immunization with recombinant-DNA encoded avian coronavirus nucleocapsid protein. Vaccine, 10: 119-124.
CrossRef - Boroomand, Z., K. Asasi and A. Mohammadi, 2012. Pathogenesis and tissue distribution of avian infectious bronchitis isolate IRFIBV32 (793/B serotype) in experimentally infected broiler chickens. Sci. World J.
CrossRef - Bourogaa, H., I. Hellal, J. Hassen, I. Fathallah and A. Ghram, 2012. S1 gene sequence analysis of new variant isolates of avian infectious bronchitis virus in Tunisia. Vet. Med.: Res. Rep., 3: 41-48.
CrossRef - Bourogaa, H., K. Miled, L. Gribaa, I.E. Behi and A. Ghram, 2009. Characterization of new variants of avian infectious bronchitis virus in Tunisia. Avian Dis., 53: 426-433.
PubMed - Brandao, P.E., 2010. Avian infectious bronchitis virus in Brazil: A highly complex virus meets a highly susceptible host population. Rev. Brasil. Cienc. Avic., 12: 121-124.
CrossRef - Brown, T.P., J.R. Glisson, G. Rosales, P. Villegas and R.B. Davis, 1987. Studies of avian urolithiasis associated with an infectious bronchitis virus. Avian Dis., 31: 629-636.
Direct Link - Butcher, G.D., R.W. Winterfield and D.P. Shapiro, 1989. An outbreak of nephropathogenic H13 infectious bronchitis in commercial broilers. Avian Dis., 33: 823-826.
Direct Link - Callison, S.A., D.A. Hilt, T.O. Boynton, B.F. Sample, R. Robison, D.E. Swayne and M.W. Jackwood, 2006. Development and evaluation of a real-time Taqman RT-PCR assay for the detection of infectious bronchitis virus from infected chickens. J. Virol. Methods, 138: 60-65.
CrossRef - Callison, S.A., M.W. Jackwood and D.A. Hilt, 2001. Molecular characterization of infectious bronchitis virus isolates foreign to the United States and comparison with United States isolates. Avian Dis., 45: 492-499.
Direct Link - Capua, I., Z. Minta, E. Karpinska, K. Mawditt, P. Britton, D. Cavanagh and R.E. Gough, 1999. Co-circulation of four types of infectious bronchitis virus (793/B, 624/I, B1648 and Massachusetts). Avian Pathol., 28: 587-592.
CrossRefDirect Link - Casais, R., B. Dove, D. Cavanagh and P. Britton, 2003. Recombinant avian infectious bronchitis virus expressing a heterologous spike gene demonstrates that the spike protein is a determinant of cell tropism. J. Virol., 77: 9084-9089.
CrossRef - Chen, H.W., Y.P. Huang and C.H. Wang, 2009. Identification of Taiwan and China-like recombinant avian infectious bronchitis viruses in Taiwan. Virus Res., 140: 121-129.
CrossRef - Cavanagh, D., 2005. Coronaviruses in poultry and other birds. Avian Pathol., 34: 439-448.
CrossRefDirect Link - Cavanagh, D., 2007. Coronavirus avian infectious bronchitis virus. Vet. Res., 38: 281-297.
CrossRefDirect Link - Chen, B.Y., S. Hosi, T. Nunoya and C. Itakura, 1996. Histopathology and immunohistochemistry of renal lesions due to infectious bronchitis virus in chicks. Avian Pathol., 25: 269-283.
CrossRef - Chen, H.T., J. Zhang, Y.P. Ma, L.N. Ma and Y.Z. Ding et al., 2010. Reverse transcription loop-mediated isothermal amplification for the rapid detection of infectious bronchitis virus in infected chicken tissues. Mol. Cell. Probes, 24: 104-106.
CrossRef - Chen, H.W. and C.H. Wang, 2010. A multiplex reverse transcriptase-PCR assay for the genotyping of avian infectious bronchitis viruses. Avian Dis., 54: 104-108.
PubMed - Collisson, E.W., J. Li, L.W. Sneed, M.L. Peters and L. Wang, 1990. Detection of avian infectious bronchitis viral infection using in situ hybridization and recombinant DNA. Vet. Microbiol., 24: 261-271.
CrossRef - Cook, J.K.A., M. Jackwood and R.C. Jones, 2012. The long view: 40 years of infectious bronchitis research. Avian Pathol., 41: 239-250.
CrossRefDirect Link - Cook, J.K.A., S.J. Orbell, M.A. Woods and M.B. Huggins, 1996. A survey of the presence of a new infectious bronchitis virus designated 4/91 (793B). Vet. Rec., 138: 178-180.
Direct Link - Cook, J.K.A., S.J. Orbell, M.A. Woods and M.B. Huggins, 1999. Breadth of protection of the respiratory tract provided by different live-attenuated infectious bronchitis vaccines against challenge with infectious bronchitis viruses of heterologous serotypes. Avian Pathol., 28: 477-485.
CrossRefDirect Link - Cook, J.K., J.H. Darbyshire and R.W. Peters, 1976. The use of chicken tracheal organ cultures for the isolation and assay of avian infectious bronchitis virus. Arch. Virol., 50: 109-118.
PubMed - Cumming, R.B., 1963. Infectious avian nephrosis (uraemia) in Australia. Aust. Vet. J., 39: 145-147.
CrossRef - Deb, R., S. Chakraborty, B. Veeregowda, A.K. Verma, R. Tiwari and K. Dhama, 2013. Monoclonal antibody and its use in the diagnosis of livestock diseases. Adv. Biosci. Biotechnol., 4: 50-62.
CrossRefDirect Link - Montassier, M.F.S., L. Brentano, H.J. Montassier and L.J. Richtzenhain, 2008. Genetic grouping of avian infectious bronchitis virus isolated in Brazil based on RT-PCR/RFLP analysis of the S1 gene1. Pesquisa Veterinaria Brasileira, 28: 190-194.
Direct Link - De Wit, J.J. and H. van de Sande, 2009. Efficacy of combined vaccines at day of hatch against a D388 challenge in SPF and commercial chickens. Proceedings of the 6th International Symposium on Corona and Pneumoviruses and Complicating Pathogens, June 14-17, 2009, Rauischholzhausen, Germany, pp: 177-182.
- De Wit, J.J., 2000. Detection of infectious bronchitis virus. Avian Pathol., 29: 71-93.
CrossRefDirect Link - De Wit, J.J.S., J.K.A. Cook and H.M.J.F. van der Heijden, 2010. Infectious bronchitis virus in Asia, Africa, Australia and Latin America-history, current situation and control measures. Rev. Bras. Cienc. Avic., 12: 97-106.
Direct Link - Dhama, K., M. Mahendran, R. Somvanshi and M.M. Chawak, 2008. Chicken infectious anaemia virus: An immunosuppressive pathogen of poultry-A review. Indian J. Vet. Pathol., 32: 158-167.
Direct Link - Dhama, K., M. Mahendran, P.K. Gupta and A. Rai, 2008. DNA vaccines and their applications in veterinary practice: Current perspectives. Vet. Res. Commun., 32: 341-356.
CrossRefDirect Link - Dhama, K., V. Verma, P.M. Sawant, R. Tiwari, R.K. Vaid and R.S. Chauhan, 2011. Applications of probiotics in poultry: Enhancing immunity and beneficial effects on production performances and health: A review. J. Immunol. Immunopathol., 13: 1-19.
Direct Link - Dhama, K., S. Chakraborty, R. Tiwari, A. Kumar and A. Rahal et al., 2013. Avian/bird flu virus: Poultry pathogen having zoonotic and pandemic threats: A review. J. Med. Sci., 13: 301-315.
CrossRefDirect Link - Dhama, K., A.K. Verma, R. Tiwari, S. Chakraborty and K. Vora et al., 2013. A perspective on applications of Geographical Information System (GIS): An advanced tracking tool for disease surveillance and monitoring in veterinary epidemiology. Adv. Anim. Vet. Sci., 1: 14-24.
Direct Link - Dhama, K., K. Karthik, S. Chakraborty, R. Tiwari, S. Kapoor, A. Kumar and P. Thomas, 2014. Loop-mediated isothermal amplification of DNA (LAMP): A new diagnostic tool lights the world of diagnosis of animal and human pathogens: A review. Pak. J. Biol. Sci., 17: 151-166.
CrossRefDirect Link - Dhama, K., S. Chakraborty, Mahima, M.Y. Wani and A.K. Verma et al., 2013. Novel and emerging therapies safeguarding health of humans and their companion animals: A review. Pak. J. Biol. Sci., 16: 101-111.
CrossRefDirect Link - Dhama, K., S. Chakraborty and R. Tiwari, 2013. Panchgavya therapy (Cowpathy) in safeguarding health of animals and humans: A review. Res. Opin. Anim. Vet. Sci., 3: 170-178.
Direct Link - Dhama, K., S. Chakraborty, M.Y. Wani, R. Tiwari and R. Barathidasan, 2013. Cytokine therapy for combating animal and human diseases-A review. Res. Opin. Anim. Vet. Sci., 3: 195-208.
Direct Link - Dhama, K., S. Chakraborty, S. Kapoor, R. Tiwari and A. Kumar et al., 2013. One world, one health-veterinary perspectives. Adv. Anim. Vet. Sci., 1: 5-13.
Direct Link - Di Fabio, J., L.I. Rossini, S.J. Orbell, G. Paul and M.B. Huggins et al., 2000. Characterization of infectious bronchitis viruses isolated from outbreaks of disease in commercial flocks in Brazil. Avian Dis., 44: 582-589.
Direct Link - Owoade, A.A., D.O. Oluwayelu, O.A. Fagbohun, W. Ammerlaan, M.N. Mulders and C.P. Muller, 2004. Serologic evidence of chicken infectious anemia in commercial chicken flocks in Southwest Nigeria. Avian Dis., 48: 202-205.
CrossRefPubMedDirect Link - Ducatez, M.F., A.M. Martin, A.A. Owoade, I.O. Olatoye and B.R. Alkali et al., 2009. Characterization of a new genotype and serotype of infectious bronchitis virus in Western Africa. J. Gen. Virol., 90: 2679-2685.
Direct Link - El Bouqdaoui, M., R.A. Mhand, H. Bouayoune and M.M. Ennaji, 2005. Genetic grouping of nephropathogenic avian infectious bronchitis virus isolated in Morocco. Int. J. Poult. Sci., 4: 721-727.
Direct Link - El Houadfi, M. and R.C. Jones, 1985. Isolation of avian infectious bronchitis viruses in Morocco including an enterotropic variant. Vet. Rec., Vol. 116.
CrossRefDirect Link - Elankumaran, S., C. Balachandran, N.D. Chandran, P. Roy, A. Albert and R. Manickam, 1999. Serological evidence for a 793/B related avian infectious bronchitis virus in India. Vet. Rec., 144: 299-300.
PubMedDirect Link - Susan, S.E.M., M.M. El-Hady and Y.A. Soliman, 2010. Isolation and characterization of nephropathogenic strain of infectious bronchitis virus in EGYPT. J. Am. Sci., 6: 669-675.
Direct Link - Escorcia, M., T.I. Fortoul, V.M. Petrone, F. Galindo, C. Lopez and G. Tellez, 2002. Gastric gross and microscopic lesions caused by the UNAM-97 variant strain of infectious bronchitis virus after the eighth passage in specific pathogen-free chicken embryos. Poult. Sci., 81: 1647-1652.
Direct Link - Fan, W.Q., H.N. Wang, Y. Zhang, Z.B. Guan and T. Wang et al., 2012. Comparative dynamic distribution of avian infectious bronchitis virus M41, H120 and SAIBK strains by quantitative real-time RT-PCR in SPF chickens. Biosci. Biotechnol. Biochem., 76: 2255-2260.
PubMedDirect Link - Gallardo, R.A., F.J. Hoerr, W.D. Berry, V.L. van Santen and H. Toro, 2011. Infectious bronchitis virus in testicles and venereal transmission. Avian Dis., 55: 255-258.
CrossRefPubMedDirect Link - Gelb, J., J.B. Wolff and C.A. Moran, 1991. Variant serotypes of infectious bronchitis virus isolated from commercial layer and broiler chickens. Avian Dis., 35: 82-87.
PubMed - Gelb, J. Jr., J.K. Rosenberger, P.A. Fries, S.S. Cloud, E.M. Odor, J.E. Dohms and JS Jaeger, 1989. Protection afforded infectious bronchitis virus-vaccinated sentinel chickens raised in a commercial environment. Avian Dis., 33: 764-769.
PubMedDirect Link - Gelb, Jr. J., Y. Weisman, B.S. Ladman and R. Meir, 2005. S1 gene characteristics and efficacy of vaccination against infectious bronchitis virus field isolates from the United States and Israel (1996 to 2000). Avian Pathol., 34: 194-203.
CrossRefPubMedDirect Link - Gowthaman, V., S.D. Singh, K. Dhama, R. Barathidasan and M.A. Kumar et al., 2012. Fowl Adenovirus (FAdV) in India: Evidence for emerging role as primary respiratory pathogen in chickens. Pak. J. Biol. Sci., 15: 900-903.
Direct Link - Hanada, K., Y. Suzuki and T. Gojobori, 2004. A large variation in the rates of synonymous substitution for RNA viruses and its relationship to a diversity of viral infection and transmission modes. Mol. Biol. Evol., 21: 1074-1080.
CrossRefDirect Link - He, K., M. Li, P. Wei, M.L. Mo, T.C. Wei and K.R. Li, 2012. Complete genome sequence of an infectious bronchitis virus chimera between cocirculating heterotypic strains. J. Virol., 86: 13887-13888.
CrossRefDirect Link - Hilton, A., L. Mizzen, G. Macintyre, S. Cheley and R. Anderson, 1986. Translational control in murine hepatitis virus infection. J. Gen. Virol., 67: 923-932.
PubMedDirect Link - Hiscox, J.A. and L.A. Ball, 1997. Cotranslational disassembly of flock house virus in a cell-free system. J. Virol., 71: 7974-7977.
Direct Link - Hiscox, J.A., T. Wurm, L. Wilson, P. Britton, D. Cavanagh and G. Brooks, 2001. The coronavirus infectious bronchitis virus nucleoprotein localizes to the nucleolus. J. Virol., 75: 506-512.
CrossRefDirect Link - Hong, S.M., H.J. Kwon, I.H. Kim, M.L. Mo and J.H. Kim, 2012. Comparative genomics of Korean infectious bronchitis viruses (IBVs) and an animal model to evaluate pathogenicity of IBVs to the reproductive organs. Viruses, 4: 2670-2683.
CrossRefDirect Link - Ignjatovic, J. and P.G. McWaters, 1991. Monoclonal antibodies to three structural proteins of avian infectious bronchitis virus: Characterization of epitopes and antigenic differentiation of Australian strains. J. Gen. Virol., 72: 2915-2922.
CrossRefDirect Link - Ignjatovic, J., D.F. Ashton, R. Reece, P. Scott and P. Hooper, 2002. Pathogenicity of Australian strains of avian infectious bronchitis virus. J. Comp. Pathol., 126: 115-123.
CrossRefDirect Link - Ignjatovic, J., S.I. Sapats and F. Ashton, 1997. A long‐term study of Australian infectious bronchitis viruses indicates a major antigenic change in recently isolated strains. Avian Pathol., 26: 535-552.
CrossRefDirect Link - Ignjatovic, J. and S. Sapats, 2000. Avian infectious bronchitis virus. Revue Scientifique Technique, 19: 493-508.
PubMedDirect Link - Jackwood, M.W., 2012. Review of infectious bronchitis virus around the world. Avian Dis., 56: 634-641.
PubMedDirect Link - Jackwood, M.W., D. Hall and A. Handel, 2012. Molecular evolution and emergence of avian gammacoronaviruses. Infect. Genet. Evol., 12: 1305-1311.
CrossRef - Jackwood, M.W., D.A. Hilt and S.A. Callison, 2003. Detection of infectious bronchitis virus by real-time reverse transcriptase-polymerase chain reaction and identification of a quasispecies in the Beaudette strain. Avian Dis., 47: 718-724.
Direct Link - Jackwood, M.W., D.A. Hilt, A.W. McCall, C.N. Polizzi, E.T. McKinley and S.M. Williams, 2009. Infectious bronchitis virus field vaccination coverage and persistence of Arkansas-type viruses in commercial broilers. Avian Dis., 53: 175-183.
PubMed - Jackwood, M.W., D.A. Hilt, C.W. Lee, H.M. Kwon and S.A. Callison et al., 2005. Data from 11 years of molecular typing infectious bronchitis virus field isolates. Avian Dis., 49: 614-618.
PubMedDirect Link - Jackwood, M.W., D.A. Hilt, S.M. Williams, P. Woolcock, C. Cardona and R. O'Connor, 2007. Molecular and serologic characterization, pathogenicity, and protection studies with infectious bronchitis virus field isolates from California. Avian Dis., 5: 527-533.
PubMed - Jones, R.C., 2010. Viral respiratory diseases (ILT, aMPV infections, IB): Are they ever under control? Br. Poult. Sci., 51: 1-11.
CrossRef - Kataria, J.M., C.M. Mohan, S. Dey, B.B. Dash and K. Dhama, 2005. Diagnosis and immunoprophylaxis of economically important poultry diseases: A review. Indian J. Anim. Sci., 75: 555-567.
Direct Link - Kelly, P.J., D. Chitauro, C. Rohde, J. Rukwava, A. Majok, F. Davelaar and P.R. Mason, 1994. Diseases and management of backyard chicken flocks in Chitungwiza, Zimbabwe. Avian Dis., 38: 626-629.
Direct Link - Kinde, H., B.M. Daft, A.E. Castro, A.A. Bickford, J. Gelb Jr. and B. Reynolds, 1991. Viral pathogenesis of a nephrotropic infectious bronchitis virus isolated from commercial pullets. Avian Dis., 35: 415-421.
Direct Link - Kingham, B.F., C.L. Keeler Jr., W.A. Nix, B.S. Ladman and J. Gelb Jr., 2000. Identification of avian infectious bronchitis virus by direct automated cycle sequencing of the S-1 gene. Avian Dis., 44: 325-335.
Direct Link - Dhama, K., M.Y. Wani, R. Deb, K. Karthik and R. Tiwari et al., 2013. Plant based oral vaccines for human and animal pathogens-a new era of prophylaxis: Current and future prospective. J. Exp. Biol. Agric. Sci., Vol. 1.
Direct Link - Kuo, S.M., C.H. Wang, M.H. Hou, Y.P. Huang, H.W. Kao and H.L. Su, 2010. Evolution of infectious bronchitis virus in Taiwan: Characterisation of RNA recombination in the Nucleocapsid gene. Vet. Microbiol., 144: 293-302.
CrossRef - Kuo, S.M., H.W. Kao, M.H. Hou, C.H. Wang, S.H. Lin and H.L. Su, 2013. Evolution of infectious bronchitis virus in Taiwan: Positively selected sites in the nucleocapsid protein and their effects on RNA-binding activity. Vet. Microbiol., 162: 408-418.
CrossRef - Kusters, J.G., H.G.M. Niesters, N.M.C. Bleumynk-Pluym, F.G. Davelaar, M.C. Horzinek and B.A.M. van der Zeist, 1987. Molecular epidemiology of infectious bronchitis in the Netherlands. J. Gen. Virol., 68: 343-352.
PubMed - Ladman, B.S., C.R. Pope, A.F. Ziegler, T. Swieczkowski, C.J. Callahan, S. Davison and J. Gelb Jr., 2002. Protection of chickens after live and inactivated virus vaccination against challenge with nephropathogenic infectious bronchitis virus PA/Wolgemuth/98. Avian Dis., 46: 938-944.
PubMed - Lai, M.M.C., 1996. Recombination in large RNA viruses: Coronaviruses. Semin. Virol., 7: 381-388.
CrossRefDirect Link - Lee, C.W. and M.W. Jackwood, 2001. Origin and evolution of Georgia 98 (GA98), a new serotype of avian infectious bronchitis virus. Virus Res., 80: 33-39.
CrossRefDirect Link - Li, H. and H.C. Yang, 2001. Sequence analysis of nephropathogenic infectious bronchitis virus strains of the Massachusetts genotype in Beijing. Avian Pathol., 30: 535-541.
CrossRefDirect Link - Li, M., X.Y. Wang, P. Wei, Q.Y. Chen, Z.J. Wei and M.L. Mo, 2012. Serotype and genotype diversity of infectious bronchitis viruses isolated during 1985-2008 in Guangxi, China. Arch. Virol., 157: 467-474.
CrossRef - Lim, T.H., M.S. Kim, J.H. Jang, D.H. Lee and J.K. Park et al., 2012. Live attenuated nephropathogenic infectious bronchitis virus vaccine provides broad cross protection against new variant strains. Poult Sci., 91: 89-94.
CrossRef - Lin, K.H., C.F. Lin, S.S. Chiou, A.P. Hsu and M.S. Lee et al., 2012. Application of purified recombinant antigenic spike fragments to the diagnosis of avian infectious bronchitis virus infection. Applied Microbiol. Biotechnol., 95: 233-242.
CrossRefDirect Link - Liu, S. and X. Kong, 2004. A new genotype of nephropathogenic infectious bronchitis virus circulating in vaccinated and non-vaccinated flocks in China. Avian Pathol., 33: 321-327.
CrossRefPubMedDirect Link - Liu, X., H. Ma, Q. Xu, N. Sun and Z. Han et al., 2013. Characterization of a recombinant coronavirus infectious bronchitis virus with distinct S1 subunits of spike and nucleocapsid genes and a 3' untranslated region. Vet. Microbiol., 162: 429-436.
CrossRefDirect Link - Liu, X., Y. Shao, H. Ma, C. Sun and X. Zhang et al., 2013. Comparative analysis of four Massachusetts type infectious bronchitis coronavirus genomes reveals a novel Massachusetts type strain and evidence of natural recombination in the genome. Infect. Genet. Evol., 14: 29-38.
CrossRefDirect Link - Lohr, J.E., 1976. Serologic differences between strains of infectious bronchitis virus from New Zealand, Australia and the United States. Avian Dis., 20: 478-482.
Direct Link - Lopez, J.C. and R. McFarlane, 2006. Environmental factors influence the prevalence of infectious bronchitis virus. Proceedings of the 18th Australian Poultry Science Symposium, February 20-22, 2006, Poultry Research Foundation, University of Sydney, Camden, New South Wales, Australia, pp: 127-130.
Direct Link - Mahima, A. Rahal, R. Deb, S.K. Latheef and H.A. Samad et al., 2012. Immunomodulatory and therapeutic potentials of herbal, traditional/indigenous and ethnoveterinary medicines. Pak. J. Biol. Sci., 15: 754-774.
CrossRefDirect Link - Mahmood, Z.H., R.R. Sleman and A.U. Uthman, 2011. Isolation and molecular characterization of Sul/01/09 avian infectious Bronchitis virus, indicates the emergence of a new genotype in the Middle East. Vet. Microbiol., 150: 21-27.
CrossRef - Mase, M., T. Inoue, S. Yamaguchi and T. Imada, 2008. Existence of avian infectious bronchitis virus with a European-prevalent 4/91 genotype in Japan. J. Vet. Med. Sci., 70: 1341-1344.
Direct Link - Matthijs, M.G., J.H. van Eck, W.J. Landman and J.A. Stegeman, 2003. Ability of Massachusetts-type infectious bronchitis virus to increase colibacillosis susceptibility in commercial broilers: A comparison between vaccine and virulent field virus. Avian Pathol., 32: 473-481.
PubMedDirect Link - McFarlane, R. and R. Verma, 2008. Sequence analysis of the gene coding for the S1 glycoprotein of infectious bronchitis virus (IBV) strains from New Zealand. Virus Genes, 37: 351-357.
PubMed - Meir, R., E. Rosenblut, S. Perl, N. Kass, G. Ayali, S. Perk and E. Hemsani, 2004. Identification of a novel nephropathogenic infectious bronchitis virus in Israel. Avian Dis., 48: 635-641.
PubMed - Meulemans, G. and T.P. van den Berg, 1998. Nephropathogenic avian infectious bronchitis viruses. World Poult. Sci., 54: 145-153.
CrossRefDirect Link - Meulemans, G., M. Boschmans, M. Decaesstecker, T.P. van den Berg, P. Denis and D. Cavanagh, 2001. Epidemiology of infectious bronchitis virus in Belgian broilers: A retrospective study, 1986 to 1995. Avian Pathol., 30: 411-421.
CrossRefDirect Link - Thekisoe, M.M., P.A. Mbati and S.P. Bisschop, 2003. Diseases of free-ranging chickens in the Qwa-Qwa District of the northeastern Free State province of South Africa. J. S. Afr. Vet. Assoc., 74: 14-16.
PubMedDirect Link - Mo, M.L., S.M. Hong, H.J. Kwon, I.H. Kim, C.S. Song and J.H. Kim, 2013. Genetic diversity of spike, 3a, 3b and e genes of infectious bronchitis viruses and emergence of new recombinants in Korea. Viruses, 5: 550-567.
Direct Link - Mushi, E.Z., M.G. Binta, R.G. Chabo and K. Itebeng, 2006. Diseases of indigenous chickens in Bokaa village, Kgatleng district, Botswana. J. S. Afr. Vet. Assoc., 77: 131-133.
PubMed - Nakamura, K., J.K. Cook, K. Otsuki, M.B. Huggins and J.A. Frazier, 1991. Comparative study of respiratory lesions in two chicken lines of different susceptibility infected with infectious bronchitis virus: Histology, ultrastructure and immunohistochemistry. Avian Pathol., 20: 241-257.
CrossRefPubMedDirect Link - Shi, P., L. Yu, Y. Fu, J.F. Huang, K.Q. Zhang and Y. Zhang, 2006. Evolutionary implications of Avian Infectious Bronchitis Virus (AIBV) analysis. Cell Res., 16: 323-327.
CrossRef - Natesan, S., J.M. Kataria, K. Dhama, S. Rahul and N. Baradhwaj, 2006. Biological and molecular characterization of Chicken anaemia virus isolates of Indian origin. Virus Res., 118: 78-86.
CrossRefPubMedDirect Link - Nelson, G.W., S.A. Stohlman and S.M. Tahara, 2000. High affinity interaction between nucleocapsid protein and leader/intergenic sequence of mouse hepatitis virus RNA. J. Gen. Virol., 81: 181-188.
PubMed - Nguyen, T.T., H.J. Kwon, I.H. Kim, S.M. Hong, W.J. Seong, J.W. Jang and J.H. Kim, 2013. Multiplex nested RT-PCR for detecting avian influenza virus, infectious bronchitis virus and Newcastle disease virus. J. Virol. Methods, 188: 41-46.
CrossRefDirect Link - Parsons, D., M.M. Ellis, D. Cavanagh and J.K. Cook, 1992. Characterisation of an infectious bronchitis virus isolated from vaccinated broiler breeder flocks. Vet. Rec., 131: 408-411.
PubMed - Pennycott, T.W., 2000. Causes of mortality and culling in adult pheasants. Vet. Rec., 146: 273-278.
PubMedDirect Link - Pohuang, T., N. Chansiripornchai, A. Tawatsin and J. Sasipreeyajan, 2009. Detection and molecular characterization of infectious Bronchitis virus isolated from recent outbreaks in broiler flocks in Thailand. J. Vet. Sci., 10: 219-223.
PubMed - Rahul, S., J.M. Kataria, N. Senthilkumar and K. Dhama, S.A. Sylvester and R. Uma, 2005. Association of fowl adenovirus serotype-12 with hydropericardium syndrome of poultry in India. Acta Virologica., 49: 139-143.
PubMed - Ramirez-Gonzalez, S., E.J. Gutierrez-Ruiz, F.J. Aranda-Cirerol, R. Rodriguez-Vivas and M.E. Bolio-Gonzalez, 2012. Isolation and antigenic characterization of infectious bronchitis virus from backyard chickens in Yucatan, Mexico. Int. Res. J. Agricul. Sci. Soil Sci., 2: 063-067.
Direct Link - Rimondi, A., M.I. Craig, A. Vagnozzi, G. Konig, M. Delamer and A. Pereda, 2009. Molecular characterization of avian infectious bronchitis virus strains from outbreaks in Argentina (2001-2008). Avian Pathol., 38: 149-153.
CrossRefPubMedDirect Link - Roussan, D.A., G.Y. Khawaldeh and I.A. Shaheen, 2009. Infectious bronchitis virus in Jordanian chickens: Seroprevalence and detection. Can. Vet. J., 50: 77-80.
PubMed - Roussan, D.A., W.S. Totanji and G.Y. Khawaldeh, 2008. Molecular subtype of infectious bronchitis virus in broiler flocks in Jordan. Poult. Sci., 87: 661-664.
CrossRef - Gharaibeh, S.M., 2007. Infectious bronchitis virus serotypes in poultry flocks in Jordan. Preventive Vet. Med., 78: 317-324.
CrossRefPubMedDirect Link - Sabarinath, A., G.P. Sabarinath, K.P. Tiwari, S.M. Kumthekar, D. Thomas and R.N. Sharma, 2011. Seroprevalence of infectious bronchitis virus in birds of grenada. Int. J. Poult. Sci., 10: 266-268.
CrossRefDirect Link - Sapats, S.I., F. Ashton, P.J. Wright and J. Ignjatovic, 1996. Sequence analysis of the S1 glycoprotein of infectious bronchitis viruses: Identification of a novel genotypic group in Australia. J. Gen. Virol., 77: 413-418.
PubMedDirect Link - Siddell, S., H. Wege, A. Barthel and V. ter Meulen, 1981. Intracellular protein synthesis and the In vitro translation of coronavirus JHM mRNA. Adv. Exp. Med. Biol., 142: 193-207.
CrossRefPubMedDirect Link - Singh, S.D., R. Barathidasan, A. Kumar, R. Deb, A.K. Verma and K. Dhama, 2012. Recent trends in diagnosis and control of Marek's Disease (MD) in poultry. Pak. J. Biol. Sci., 15: 964-970.
CrossRefDirect Link - De Wit, J.J., J.K.A. Cook and H.M.J.F. van der Heijden, 2011. Infectious bronchitis virus variants: A review of the history, current situation and control measures. Avian Pathol., 40: 223-235.
CrossRefDirect Link - Sumi, V., S.D. Singh, K. Dhama, V. Gowthaman, R. Barathidasan and K. Sukumar, 2012. Isolation and molecular characterization of infectious bronchitis virus from recent outbreaks in broiler flocks reveals emergence of novel strain in India. Trop. Anim. Health Prod., 44: 1791-1795.
CrossRef - Sylvester, S.A., J.M. Kataria, K. Dhama, S. Rahul, N. Bhardwaj and S. Tomar, 2003. Purification of infectious bronchitis virus propagated in embryonated chicken eggs and its confirmation by RT-PCR. Indian J. Comp. Microbiol. Immunol. Infect. Dis., 24: 143-147.
Direct Link - Tahara, S.M., T.A. Dietlin, C.C. Bergmann, G.W. Nelson, S. Kyuwa, R.P. Anthony and S.A. Stohlman, 1994. Coronavirus translational regulation: leader affects mRNA efficiency. Virology, 202: 621-630.
CrossRef - Terregino, C., A. Toffan, M.S. Beato, R. De Nardi and M. Vascellari et al., 2008. Pathogenicity of a QX strain of infectious bronchitis virus in specific pathogen free and commercial broiler chickens and evaluation of protection induced by a vaccination programme based on the Ma5 and 4/91 serotypes. Avian Pathol., 37: 487-493.
CrossRef - Tiwari, R., S. Chakraborty, K. Dhama, M.Y. Wani, A. Kumar and S. Kapoor, 2013. Wonder world of phages: Potential biocontrol agents safeguarding biosphere and health of animals and humans-current scenario and perspectives. Pak. J. Biol. Sci., (In Press).
Direct Link - Toro, H., D. Pennington, R.A. Gallardo, V.L. van Santen, F.W. van Ginkel, J. Zhang and K.S. Joiner, 2012. Infectious bronchitis virus subpopulations in vaccinated chickens after challenge. Avian Dis., 56: 501-508.
Direct Link - Toro, H., V.L. van Santen and M.W. Jackwood, 2012. Genetic diversity and selection regulates evolution of infectious bronchitis virus. Avian Dis., 56: 449-455.
CrossRefPubMedDirect Link - Villarreal, L.Y.B., P.E. Brandao, J.L. Chacon, M.S. Assayag and P.C. Maiorka et al., 2007. Orchitis in roosters with reduced fertility associated with avian infectious bronchitis virus and avian meta pneumovirus infections. Avian Dis., 51: 900-904.
Direct Link - Wang, H.N., Q.Z. Wu, Y. Huang and P. Liu, 1997. Isolation and identification of infectious bronchitis virus from chickens in Sichuan, China. Avian Dis., 41: 279-282.
Direct Link - Welchman, de B., J.M. Bradbury, D. Cavanagh and N.J. Aebischer, 2002. Infectious agents associated with respiratory disease in pheasants. Vet. Record, 150: 658-664.
CrossRefDirect Link - Wu, Z.Q., Q.W. Yang, C. Fu, X.Y. Zhaoand and J. Ignjatovic, 1998. Antigenic and immunogenic characterization of infectious bronchitis virus strains isolated in China between 1986 and 1995. Avian Pathol., 27: 578-585.
CrossRef - Yu, L., Z. Wang, Y. Jiang, S. Low and J. Kwang, 2001. Molecular epidemiology of infectious bronchitis virus isolates from China and Southeast Asia. Avian Dis., 45: 201-209.
PubMed - Zanella, A., A. Lavazza, R. Marchi, A.M. Martin and F. Pagnelli, 2003. Avian infectious bronchitis: Characterization of new isolates from Italy. Avian Dis., 47: 180-185.
PubMed - Ziegler, A.F., B.S. Ladman, P.A. Dunn, A. Schneider and S. Davison et al., 2002. Nephropathogenic infectious bronchitis in pennsylvania chickens 1997-2000. Avian Dis., 46: 847-858.
CrossRefDirect Link - Zwaagstra, K.A., B.A.M. Van der Zeijst and J.G. Kusters, 1992. Rapid detection and identification of avian infectious bronchitis virus. J. Clin. Microbiol., 30: 79-84.
Direct Link