
J. Hongthongkham
Department of Biology, Faculty of Science, Khon Kaen University, 123/2001 Moo 16, Friendship Highway, Naimuaeng Sub-district, Muaeng District, Khonkaen Province, 40002, Thailand
S. Bunnag
Department of Biology, Faculty of Science, Khon Kaen University, 123/2001 Moo 16, Friendship Highway, Naimuaeng Sub-district, Muaeng District, Khonkaen Province, 40002, Thailand
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 5 | Page No.: 608-618







ABSTRACT
An efficient method for in vitro propagation and cryopreservation of Aerides odorata was established. Leaf segments were cultured on New Dogashima (ND) mediums supplemented with various concentrations of Benzyladenine (BA) (0-5 mg L-1) combined with Naphthaleneacetic Acid (NAA) (0-2 mg L-1). The optimal treatment for inducing Protocorm-like Bodies (PLBs) from leaf segments was obtained from the combination of 1 or 3 mg L-1 BA and 0.5 or 1 mg L-1 NAA; whereas, the addition of BA or NAA alone induced shoot and/or root initiation rather than PLB or callus formation. Shoots rapidly developed on ND mediums containing 5 mg L-1 BA. Cryopreservation of leaf segment-derived PLBs was successful using the encapsulation-dehydration method. The maximum survival percentage of Cryopreserved (Cryp) PLBs was achieved by encapsulating PLBs with 2% Na-alginate combined with 2 M glycerol and 0.4 M sucrose. The encapsulated PLBs were then precultured in 0.75 M sucrose for 24 h and dehydrated for 6 h before plunging into liquid nitrogen. Genetic stability of Cryp PLBs after regrowth was assessed by flow cytometry. The findings showed no different patterns of ploidy levels and morphology between Cryp and non-cryopreserved (Ncryp) control plantlets.
PDF Abstract XML References Citation
Received: June 01, 2013;
Accepted: August 28, 2013;
Published: November 27, 2013
How to cite this article
J. Hongthongkham and S. Bunnag, 2014. In vitro Propagation and Cryopreservation of Aerides odorata
Lour. (Orchidaceae). Pakistan Journal of Biological Sciences, 17: 608-618.
DOI: 10.3923/pjbs.2014.608.618
URL: https://scialert.net/abstract/?doi=pjbs.2014.608.618
DOI: 10.3923/pjbs.2014.608.618
URL: https://scialert.net/abstract/?doi=pjbs.2014.608.618
INTRODUCTION
The species Aerides odorata is a wild orchid of Thailand, an epiphytic orchid with a monopodial growth habit. It is collectively referred to as ‘Fox-tail’ orchid because of its flowered brush-like inflorescences of white, pink or purple flowers. Moreover, its flowers are highly fragrant with sweet citrus scent. In recent year, this orchid species became valuable because of its capacity to cross-breed with other species within the same genus and other genera, e.g., Vanda and Rhyncostylis. Currently, the species A. odorata has been gradually depopulated due to rapid destruction of natural habitats as a result from deforestation and logging and the expansion of agricultural areas all over the country. Wild orchid trade and over exploitation of natural resources by local forest dwellers are another cause of decline of orchid populations. Although A. odorata populations can naturally multiply through seeds or stem cuttings, their multiplication rates are restricted due to the fact that they lack endosperm in their seeds. Furthermore, they are devoid of enzymes to metabolize polysaccharides; as a result, they are considered as rare species. Therefore, we believe that, there is an urgent need to plan to obtain a method of conservation for germplasm of this orchid species.
Many conservation procedures, e.g., in vitro propagation and cryopreservation, have been introduced as an effective tool to conserve parts of endangered plant species. In vitro propagation is a technique that is used not only for a rapid increase in the number of individuals of plant species, particularly those with reproductive problems and extremely reduced populations (Gonzalez-Benito and Martin, 2011) but also for the supply of virus-free plants. Due to a simple method for producing a large number of plant populations within a short period, several parts of orchids, e.g., seeds (Avila-Diaz et al., 2009), protocorm-like bodies (Luo et al., 2008), protocorms and leaves (Diaz and Alvarez, 2009), shoot-tips (Roy et al., 2007) and axillary buds (Huang and Chung, 2011), have been widely used. Studies on in vitro propagation in the genus Aerides are limited. Many studies report only on protocorm-like body induction from leaf and protocorm segments and seedling growth (Murthy and Pyati, 2001; Sheelavanthmath et al., 2005; Prasertsongskun and Awaesuemae, 2009).
Cryopreservation is an alternative means to conserve plant genetic resources, often performed at the temperature of liquid nitrogen (-196°C). This technique is among the most efficient and cost-effective ones for long-term storage of plant materials and is also important for ex situ conservation of genetic resources in the threatened 1 plant species (Gonzalez-Arnao et al., 2009). Cryopreservation is based on the reduction and subsequent interruption of metabolic functions of biological materials by the decreasing of the temperature to the level of liquid nitrogen, while viability is maintained. During storage at 196°C, cellular divisions and metabolic processes are totally arrested. This technique allows to preserve materials for an unlimited time period, without risk of loss of regeneration capacity, genetic alteration or modifications after thawing (Engelmann, 2004).
New cryopreservation techniques, such as encapsulation-dehydration, vitrification and encapsulation-vitrification, have been widely developed to store various types of plant parts. To avoid ice crystal formation, the previously mentioned techniques principally involves sample dehydration by physical or chemical desiccation prior to freezing by rapid immersion into liquid nitrogen. Different parts from many orchid species, e.g., seeds (Hirano et al., 2005; Nikishina et al., 2001, 2007), cell suspension (Tsukazaki et al., 2000), shoot tips and shoot primordia (Lurswijidjarus and Thammasiri, 2004; Na and Kondo, 1996), protocorms and protocorm-like bodies (Maneerattanarungroj et al., 2007; Pouzi et al., 2011; Zainuddin et al., 2011), have been successfully cryopreserved using new cryopreservation techniques.
Encapsulation-dehydration, one of cryopreservation techniques, is widely used due to the fact that it is easy to handle, inexpensive and non-toxic. Moreover, it has been successfully applied to a wide range of undifferentiated tissues, e.g., embryogenic cell suspensions; shoot tips and axillary buds (Wang et al., 2002; Shatnawi et al., 2004; Suzuki et al., 2005).
Present study we have tried to make an efficient protocol for orchid micropropagation, using segments of an orchid leaf as explants and we have used an encapsulation-dehydration method for A. odorata PLB, to make its cryopreservation successful.
MATERIALS AND METHODS
Plant materials: Mature capsules (fruits) of A. odorata were collected after 6 months of pollination. They were surface-sterilized in 70% (v/v) ethyl alcohol for 5 min and subsequently soaked in 20% (v/v) sodium hypochlorite for 30 min prior to washing 3 times in sterile distilled water. Then, the capsules were cut and seeds were transferred to the modified New Dogashima (ND) medium (Tokuhara and Mii, 1993) supplemented with 10% (w/v) potato extract, 1% (w/v) sucrose and 1% (w/v) agar, pH 5.4. The cultures were kept at 25±2°C under a photoperiod of 16 h light/8 h dark with a photon dose of 37 μmoL m-2 sec-1. After 3 months, plantlets with 2-3 leaves were used as explants.
Protocorm-like Body (PLB) production induced by Benzyladenine (BA) and Naphthaleneacetic Acid (NAA): Excised leaf segments (approximately 0.5 cm in length) were cultured on ND mediums without plant growth regulator or ND mediums supplemented with various concentrations of BA (0, 1, 3 and 5 mg L-1) combined with different concentrations of NAA (0, 0.5, 1 and 2 mg L-1) for PLB induction. After 4 weeks of culture, the mean number of PLBs per explant was counted under a stereo microscope and the percentage of PLB induction was calculated based on the number of single explants showing differentiation of PLBs divided by the total number of explants in each treatment.
For plantlet regeneration, four-week-old PLBs were transferred to ND mediums supplemented with 10% (w/v) potato extract and 1% (w/v) sucrose and maintained at 25±2°C under a photoperiod of 16 h light/8 h dark with a photon dose of 37 μmoL m-2 sec-1. Plantlets developed from PLBs were subcultured monthly. All experiments were performed in triplicates, each containing ten explants.
Shoot and root formation from leaf segments induced by BA and NAA: Excised leaf segments were cultured on ND mediums supplemented with different concentrations of BA (0, 1, 3 and 5 mg L-1) or NAA (0, 0.5, 1 and 2 mg L-1) for shoot or root formation. The percentage of shoot formation, mean number of shoots per explant, percentage of root initiation and mean number of roots per shoot were recorded after 4 weeks of culture. The percentage of shoot or root formation was calculated based on the number of leaf segments showing shoot or root formation divided by the total number of leaf segments in each treatment. All experiments were performed in triplicates, each containing ten explants.
Cryopreservation of PLBs using an encapsulation-dehydration method: Encapsulation-dehydration based cryopreservation consisted of six steps: Encapsulation, pretreatment, dehydration, freezing, thawing and recovery. Briefly, PLBs, approximately 1-2 mm in diameter, were immersed in liquid ND mediums supplemented with 2% Na-alginate and a loading solution (a mixture of 2 M glycerol and 0.4 M sucrose). Each PLB was then dropped into a 100 mM CaCl2 solution prepared in liquid ND mediums containing 2 M glycerol and 0.4 M sucrose for bead forming. After 30 min, the encapsulated PLBs (one single PLB per bead) were pretreated in liquid ND mediums containing different concentrations of sucrose (0.5, 0.75 and 1 M) and kept at 25°C for 48 h. Later on, PLBs were dehydrated on a sterile filter paper laid down on an open Petri dish (40 beads/Petri dish) and exposed to sterile air-flow in a laminar air-flow cabinet for 0-8 h. Afterward, ten dehydrated beads were transferred to a 2 mL cryotube and directly plunged into liquid nitrogen (LN) for at least 1 day. After that, cryopreserved (Cryp) PLBs were withdrawn from LN and rapidly thawed in a water bath at 38±2°C for 2 min and transferred to liquid ND mediums supplemented with 1.2 M sucrose for 20 min. after recovery, the resulted PLBs were regrown on ND mediums modified with 10% (w/v) potato extract and 1% (w/v) sucrose.
Measurement of water content was performed after preculturing encapsulated PLBs in liquid ND mediums containing various concentrations of sucrose (0.5, 0.75 and 1 m), followed by dehydrating them for 0-8 h. Dry weight of the resulted PLBs was measured by air-drying at 70°C for 24 h. The percentage of water content in the resulted PLBs was calculated on the basis of fresh weight using the formula:
where, WC is water content, FW is fresh weight and DW is dry weight. Survival percentage of cryopreserved PLBs was determined by regrowing the recovered PLBs on ND mediums for 4 weeks.
Flow cytometry analysis: Leaves taken from 6-month-old Cryp plantlets and Non-Cryopreserved (Ncryp) control plantlets were used for nucleic acid extraction. Isolation of nucleic acids was carried on by extracting chopped leaves in 200 μL of nucleic acids extraction buffer. Nucleic acids were then stained with 1 mL of 4,6-diamidino-2-phenylindole (DAPI) solution [10 mM Tris, 50 mM sodium citrate, 2 mM MgCl2, 0.1% (w/v) Polyvinyl Pyrrolidone (PVP), 0.1% (w/v) Triton X-100 and 2 mg L-1 DAPI] and analyzed by Flow Cytometry (FCM) (Partec PAS).
Statistical analysis: All data were statistically analyzed using one way Analysis of Variance (ANOVA). Differences between means of each two treatments were compared using Duncan’s Multiple Range Test (DMRT) at p = 0.05.
RESULTS
PLB production induced by BA and NAA: The effects of BA and NAA on the percentage of surviving explants and PLB induction and mean number of PLBs from leaf segments are represented in Table 1. Based on the present results, high percentages of surviving explants could be obtained in mediums supplemented with BA combined with NAA. The maximum percentage of surviving explants (96) was observed in mediums supplemented with 1 mg L-1 BA combined with 0.5 mg L-1 NAA, followed by 1 mg L-1 BA combined with 1 mg L-1 NAA (94%). High levels of PLB initiation (88-96%) were observed from treatments containing 1 or 3 mg L-1 BA combined with 0.5 or 1 mg L-1 NAA. The maximum frequency of PLB initiation (96%) was observed in treatments with 0.5 mg L-1 BA combined with 1 mg L-1 NAA. The maximum number of PLBs per explant was 13.55. Moreover, PLB formation capacity seemed to be decreased when BA concentrations were raised to 5 mg L-1. In particular, high concentration of BA in combination with 2 mg L-1 NAA resulted in lowering of formation capacity and only 1.85 PLBs per explant were formed. Leaf segments cultured on ND mediums in the absence of Plant Growth Regulators (PGRs) retained green in color after 2 weeks of culture. But when week 4 arrived, the majority of explants turned brown and finally died, whilst some survived with no sign of PLB initiation.
| Table 1: | Effects of BA and NAA on the percentage of surviving explant and PLB initiation, and Mean No. of PLBs per explant from leaf segments of A. odorata after 4 weeks of culture on ND mediums |
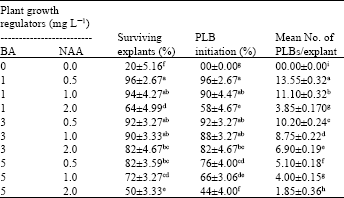 | |
| BA: Benzyladenine, NAA: Naphthaleneacetic acid, Means followed by the same letter within each column are not significantly different using Duncan’s multiple range test at p<0.05 | |
 | |
| Fig. 1(a-f): | PLB initiation and direct shoot formation from leaf segments on ND solid mediums, (a) Clump formation at the base of leaf segments on week three, (b) PLB initiation, (c) PLB development into shoots and roots, (d) direct shoot formation in the presence of 5 mg L-1 BA, (e) shoot induced roots on week six and (f) 3-month-old plantlets. Scale bars: a-b, 5 mm; c, 1 cm; d-f, 5 mm |
On the contrary, PLB initiation was found in all treatments using combinations of PGRs. The results demonstrated that small yellowish clumps initially appeared at the base of leaf segments on week 2 (Fig. 1a), continued expanding, swelling and turning green and eventually developed into PLBs (Fig. 1b). It was evident that there was a similar morphogenetic process of PLB formation among all treatments, while the frequency of PLB initiation was different. PLBs developed from segmented leaf easily developed into shoots and roots within 4 weeks and whole plantlets were obtained on week 8 after transferred to ND mediums supplemented with 10% (w/v) potato extract and 1% (w/v) sucrose without PGRs (Fig. 1c).
Shoot and root formation from leaf segments induced by BA and NAA: Table 2 shows the effects of BA and NAA on the percentage of shoot or root formation and mean number of shoots per explant or roots per shoot. Our findings showed that BA alone could be used to induce high shoot formation. The maximum percentage of shoot formation (96) was obtained in mediums supplemented with 5 mg L-1 BA with the mean number of shoots per explant of 6.05.
| Table 2: | Effects of BA and NAA on the percentage of shoot or root formation and Mean No. of shoots per explant or roots per shoot after 4 weeks of culture |
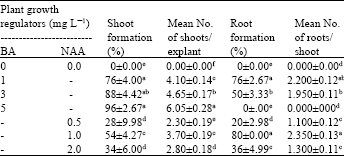 | |
| BA: Benzyladenine, NAA: Naphthaleneacetic acid, Means followed by the same letter within each column are not significantly different using Duncan’s multiple range test at p<0.05 | |
High percentage of shoot formation was also achieved in mediums with lower BA concentrations. Addition of 1 and 3 mg L-1 BA in mediums respectively gave rise to high shoot formation percentages of 76 and 88 and mean numbers of shoots per explant of 4.1 and 4.65. However, either BA or NAA alone could induce high root formation. The present results showed that mediums containing 1 mg L-1 NAA could initiate high percentages of root formation of 80, exhibiting 2.35 roots per explant. In a similar way, BA at the concentration of 1 mg L-1 gave rise to high root formation percentage of 76 with the mean number of shoots per explant of 2.2.
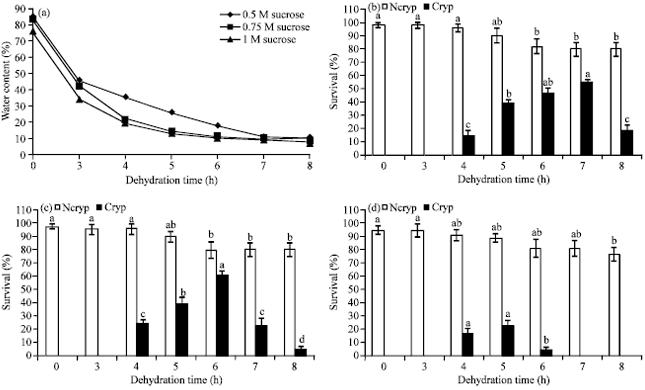 | |
| Fig. 2(a-d): | Water content and survival percentage of Ncryp and Cryp PLBs, (a) Changes in water content of encapsulated beads pretreated with various concentrations of sucrose for 24 h, followed by air-drying for 0-8 h and (b-d), survival percentage of Cryp and Ncryp PLBs when pretreated with 0.5 M, 0.75 M and 1 M sucrose, respectively |
Leaf segments cultured in the absence of PGRs remained green over a period of four weeks. Some appeared light brown in color and finally died on week 6. On the contrary, shoot formation was found on week three in all treatments with PGRs. Small protuberances primarily emerged on the surface of leaf segments and green shoots were formed (Fig. 1d). Root formation was observed two weeks after shoot formation initiated (Fig. 1e-f).
Cryopreservation of PLBs using an encapsulation-dehydration method: Figure 2 illustrates water content in encapsulated PLBs pretreated with various concentrations of sucrose, followed by different dehydration times. Significant difference in the percentage of water loss was observed among all sucrose treatments. Initial water contents of encapsulated PLBs pretreated with 0.5, 0.75 and 1 M sucrose for 24 h were 84.62, 83.26 and 76.42%, respectively. We observed that water content of sucrose-pretreated PLBs was gradually reduced within the first 3 h of dehydration and was completely decreased after 3 h of dehydration.
Presence of different sucrose concentrations in the pretreatment mediums and dehydration time had a profound effect on survival percentage of both Ncryp and Cryp PLBs. Significant difference in survival rates between Ncryp and Cryp PLBs were observed amongst all treatments. Our findings showed that increasing sucrose concentration could lower the viability of Ncryp control PLBs. Inversely, when sucrose concentration was increased; the viability of Cryp PLBs was enhanced. However, when treated with 1 M sucrose its viability was decreased to less than 23%.
Dehydrated for 4 h after pretreatments with all sucrose concentrations, Cryp PLBs showed markedly noticeable viability. In contrast, pretreated with 1 M sucrose and subsequently dehydrated for 6 h, Cryp PLBs completely died but when pretreated with 0.5 and 0.75 M sucrose, remained viable. The optimal conditions for high survival percentage were those in which encapsulated PLBs were pretreated with 0.5 and 0.75 m sucrose, followed by dehydration for 7 and 6 h, respectively. According to our findings, the maximum survival percentage was found in the pretreatment with 0.75 m sucrose and dehydration for 6 h.
Developmental stages of Cryp PLBs were shown in Fig. 3a-c. No morphological difference between Ncryp plants and Cryp plantlets.
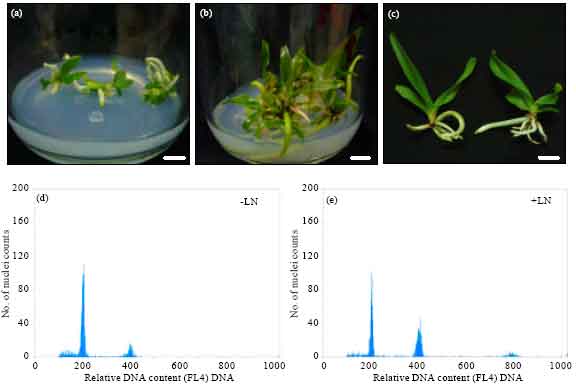 | |
| Fig. 3(a-e): | Cryopreservation of A. odorata PLBs by the encapsulation-dehydration method, ( a-b), Cryp plantlets after regrown for 3 and 12 weeks, respectively, (c) Ncryp plantlets (left) and Cryp plantlets (right) and (d-e), pattern of ploidy levels between Ncryp and Cryp plantlets analyzed by flow cytometry. Scale bar: a-c, 1 cm |
Flow cytometric analysis: Figure 3d-e illustrates ploidy levels between Ncryp plantlets and Cryp plantlets analyzed by flow cytometry. It was evident that nuclei isolated from leaves of those plantlets exhibited the same peak of DNA content, indicating no change of the ploidy levels.
DISCUSSION
The present study demonstrated that BA and NAA could be used in PLB induction and plantlet regeneration in leaf segments of A. odorata. Leaf segments showed different responses to wide range concentrations of BA or NAA. The results presented here also suggested that shoot formation could be achieved in the treatment with BA alone, while a combination of BA and NAA was more likely to promote PLB initiation. Kim and Kim (2003) and Puchooa (2004) supported that a combination of auxin and cytokinin effectively enhanced the number of PLBs and the frequency of responsive explants with PLB formation. Application of BA combined with NAA in attempts to induce PLB formation was successful in many orchid species, e.g., Phalaenopsis amabilis, Cypripedium macranthos, Dendrobium densiflorum and Cymbidium giganteum (Arditti and Ernst, 1993; Shimura and Koda, 2004; Luo et al., 2008; Hossain et al., 2010). Moreover, seedling growth of Rhynchostylis retusa was enhanced when a combination of BA and NAA was applied (Thomas and Michael, 2007). However, BA combined with NAA resulted in callus induction in shoot tips of Dendrobium chrysotoxum, whilst BA alone was optimal for PLB formation (Roy et al., 2007). In addition, some reports claimed that a combination of BA and 2,4-dichlorophynoxyacetic acid (2,4-D) was more effective than NAA for callus and PLB induction. Janarthanam and Seshadri (2008) reported that callus formation from leaf explants of Vanilla planifilia was stimulated when explants received BA combined with 2,4-D. BA alone plays an efficient role in the induction of callus or PLB formation from several orchid parts such as protocorms, leaf segments and shoot tips (Kuo et al., 2005; Sheelavanthmath et al., 2005; Roy et al., 2007). Mok et al. (2000) reported that 6-benzylaminopurine or 6-benzyladenine (BAP, BA) is adenine type of cytokinin which promotes cell divisions and cell expansions in plant culture. It has been widely used in orchids for several purposes such as somatic embryo development, PLBs proliferation and stimulation of shoot formation. Sheelavanthmath et al. (2005) observed the effect of BA on PLB formation in Aerides crispum and found that among Thidiazuron (TDZ), BA and kinetin tested, BA promoted the number of PLBs induced from protocorm and leaf explants. These results were similar to the findings of Lee and Lee (2003) who reported that 4.44 μm BA yielded the highest percentage of PLB formation and maximum number of PLBs per explant in protocorm segments of Cypripedium formosanum. However, the present study suggested that BA alone actively induced shoot formation from leaf segments. The percentage of shoot formation and mean number of shoots per explant increased when high BA concentrations were applied. The present study was in agreement with the findings of Nayak et al. (1997) who reported that addition of cytokinin to mediums was essential to induce shoot bud regeneration in foliar explants of Acampe praemorsa. In Cymbidium giganteum, the maximum number of shoot buds produced on pseudo-stem segments was obtained in the presence of 3 mg L-1 BA but high concentrations of BA restrained shoot bud production (Hossain et al., 2010). Apart from that, Santos Diaz and Alvarez (2009) found that shoot formation from leaf explants without any callus or PLB formation could be induced by a combination of BA and NAA. In particular, 22.21 μm BA combined with 10.7 μm NAA showed the maximum number of shoots (25 shoots per explant) and the longest shoot (48 mm).
In general, auxins are important to stimulate callus production and cell growth, to initiate roots and to induce somatic embryogenesis. Types of auxins which are mostly used in plant tissue culture mediums include Indole-3-butyric acid (IBA), 2,4-D and NAA. IBA and 2,4-D are considered to induce rapid cell proliferation but exposed to high concentrations with extended time, especially 2,4-D, morphogenesis process is suppressed (Kuo et al., 2005). These observations were also reported in Dendrobium, Oncidium and Phalaenopsis (Chung et al., 2007; Chen and Chang, 2001, 2006). Moreover, Zhao et al. (2008) suggested that NAA was more effective than IBA for plantlet regeneration from callus in Dendrobium candidum.
Effectiveness of NAA in callus or PLB induction was reported in different orchid species such as Aerides, Cymbidium and Paphiopedilum (Murthy and Pyati, 2001; Huan et al., 2004; Hong et al., 2008). According to the present study, NAA alone could induce shoot formation from leaf segments without interference of PLB formation. At low and moderate concentrations, NAA improved shoot formation, whilst high concentrations limited shoot formation. Likewise, Nayak et al. (1997) reported that 1 mg L-1 NAA was optimal for inducing shoot bud regeneration from foliar explants, while shoot regeneration was suppressed when NAA concentrations increased to 5 mg L-1. It could be inferred that a success in PLB production or plantlet regeneration induced by exogenous PGRs depends upon plant species, types of PGRs and concentrations.
According to the present study, leaf segment-derived PLBs showed high regeneration capacity when transferred to modified ND mediums supplemented with 10% (w/v) potato extract without PGRs. Shoots rapidly formed and development of roots was found on week four. Addition of complex organic extracts such as coconut water, banana homogenate and potato extract, to culture mediums are considered beneficial for plantlet regeneration in orchids (Arditti and Ernst, 1993). Being rich in energy, vitamins and plant hormones, complex organic extracts were widely used for promotion of shoot and root formation in Cypripedium and Doritaenopsis (Lee and Lee, 2003; Park et al., 2003), induction of seed germination in Rhynchostylis (Thomas and Michael, 2007) also in callus and PLB growth of Doritaenopsis (Islam et al., 2003; Rahman et al., 2004).
Leaf segment-derived PLBs of A. odorata were successfully cryopreserved by the encapsulation-dehydration method. In this technique, alginate beads principally play a role as seed coats to protect the embryo and as endosperm to provide nutrients. Alginate beads can protect the explants from injury which may occur during dehydration and freezing (Suzuki et al., 2005). Moreover, pretreatment conditions and optimal dehydration times are key factors for successful cryostorage. Many orchid parts such as seeds, shoot tips, cell suspension, pollen, protocorms and PLBs have been successfully cryopreserved using the encapsulation-dehydration method (Na and Kondo, 1996; Ishikawa et al., 1997; Tsukazaki et al., 2000; Hirano et al., 2005, 2009; Maneerattanarungroj et al., 2007; Thammasiri and Soamkul, 2007; Yin and Hong, 2009; Antony et al., 2010). In general, PLBs are considered as a suitable source of material for germplasm conservation because of their high regeneration capacity to develop into a normal plant, this is why we have used it in our study.
Scottez et al. (1992) suggested that freezing tolerance of plant materials could be induced by pretreatment with high levels of osmotic substances such as sucrose, sorbitol and mannitol. Sucrose has been widely used in pretreatments rather than sorbitol and mannitol because it is inexpensive. Moreover, sorbitol and mannitol were reported to inhibit shoot growth in African violet and Begoniaxerythrophylla (Burritt, 2008; Moges et al., 2004). Paul et al. (2000) reported that different concentrations of sucrose used as a pretreatment, could be used as factor of improving survival after freezing. Moges et al. (2004) used 0.1-0.75 M sucrose to improve viability and regrowth capacity of Cryp shoot tips of African violet and found that the highest survival and regrowth rates were enhanced by the pretreatment with 0.3 M sucrose for 2 days. Halmagyi and Deliu (2007) reported that high shoot regeneration from Cryp shoot tips of carnation was obtained from the pretreatment with 0.5 M sucrose for 48 h. In Dendrobium Bobby Messina, high viability of Cryp PLBs estimated by TTC staining test was improved when PLBs were pretreated with 0.6 M sucrose for 24 h (Antony et al., 2010); whereas, viability of Cryp PLBs (1-2 mm in size) of Dendrobium sonia-28 and Dendrobium Bobby Messina was enhanced when PLBs were pretreated in semi-solid half-strength MS mediums containing 1 M sucrose for 24 h (Pouzi et al., 2011; Zainuddin et al., 2011).
Shibli (2000) suggested that resistance capacity of explants to dehydration and freezing could be enhanced using sucrose-enriched mediums for 1-7 day. Sucrose in pretreatments is involved in osmotic desiccation during the treatment, leading to reduction of water content. It is absorbed into cells and replaces water molecules, resulting in accumulation of soluble sugars. Moreover, high sugar concentrations in the cell cytoplasm help to increase stabilizing membranes and proteins during desiccation and freezing condition and also help to establish a vitrified state during cooling (Shibli et al., 2001), thus enabling cells to tolerate dehydration. However, high levels of sucrose concentrations (>1 M) became toxic and its toxicity increases with a prolonged duration. Wang et al. (2005) found that Cryp shoot tips of raspberry did not survive when encapsulated shoot tips were treated with 1 M sucrose for 3 day, while using stepwise pretreatment of explants by gradually increasing sucrose concentrations from 0.25-1.25 M reduced viability of explants when final concentration reached 1.25 M. Antony et al. (2010) studied the effects of sugar types (sucrose and sorbitol) and concentrations on viability of Cryp PLBs and found that addition of 0.8-1.2 M sucrose trended to decrease viability while viability was obviously reduced in 1.4 M sorbitol. In addition, some reports indicated that the use of 1 M sucrose affected survival and regrowth rates of Cryp shoot tips, PLBs and embryogenic calli of carnation, Dendrobium candidum and Dioscorea bulbifera, respectively (Halmagyi and Deliu, 2007; Ming-Hua and Sen-Rong, 2010; Yin and Hong, 2009).
Another factor involved in viability and regrowth capacity of plant materials subject to cryopreservation is dehydration. Owing to plant cells contain high water content which can cause damage as a result from formation of ice crystals during freezing; therefore, reduction of water content is necessary to avoid injury. Wang et al. (2002) compared dehydration capacity between air-drying and silica gel in terms of survival percentage of Cryp shoot tips of grapevine and found out that the remaining water content in the explants after desiccation was a key parameter to consider rather than dehydration methods. In the encapsulation-dehydration technique, many studies indicated that the optimal water content ranged from 10-25% on a fresh weight basis enhanced viability and regrowth capacity of explants after thawing (Shatnawi et al., 2004; Suzuki et al., 2005; Wang et al., 2005) . In the present study, water content in visible Cryp PLBs ranged from 10 to 35%. The present study was in agreement with Wang et al. (2002) who reported that water content in Cryp embryogenic cell suspension viable was 20.6%. Increasing sucrose concentrations and extending dehydration times were also found to completely suppress the survival rate of Cryp PLBs.
The results presented here also indicated that the encapsulation-dehydration method did not affect plantlet regeneration. PLBs could develop into a whole plant within 7 weeks after recovery. According to flow cytometry, ploidy levels between Ncryp plantlets and Cryp plants were not different, indicating cytogenetic stability. The present study was in agreement with Hirano et al. (2005) who reported similar patterns of DNA content and ploidy levels between Ncryp and Cryp plantlets. In potatoes, comparative studies on the polyploidy levels of Ncryp and Cryp plantlets indicated that cryopreservation technique did not cause any changes in polyploidy levels (Zarghami et al., 2008).
CONCLUSION
In summary, the present study suggested a simple and efficient procedure for in vitro propagation and cryopreservation of A. odorata. According to the results, shoot formation could be induced by either BA or NAA alone, while PLBs could be induced by a combination of BA and NAA. The present study also suggested that sucrose concentrations in pretreatments and dehydration times both affected the viability and regrowth capacity of Cryp explants. To assess whether or not cryopreservation caused genetic variation, flow cytometry was considered as an important tool. However, further studies relying on other methods such as Random Amplified Polymorphic DNA (RAPD) and Restriction Fragment Length Polymorphism (RFLP), to detect genetic stability and to expand the applicability of this procedure to other orchid species remains necessary.
ACKNOWLEDGMENTS
This study was financially supported by the Ph.D. scholarship from Office of the Higher Education Commission, Ministry of Education, Thailand.
REFERENCES
- Antony, J.J.J., C.L. Keng, X. Rathinam, U.R. Sinniah and S. Subramaniam, 2010. Preliminary study on cryopreservation of Dendrobium bobby messina protocorm-like bodies by vitrification Afr. J. Biotechnol., 9: 7063-7070.
Direct Link - Avila-Diaz, I., K. Oyama, C. Gomez-Alonso and R. Salgado-Garciglia, 2009. In vitro propagation of the endangered orchid Laelia speciosa. Plant Cell Tiss.ue Organ. Cult., 99: 335-343.
CrossRefDirect Link - Burritt, D.J., 2008. Efficient cryopreservation of adventitious shoots of Begonia x erythrophylla using encapsulation-dehydration requires pretreatment with both ABA and proline. Plant Cell Tissue Organ Cult., 95: 209-215.
CrossRefDirect Link - Chen, J.T. and W.C. Chang, 2001. Effects of auxins and cytokinins on direct somatic embryogenesison leaf explants of Oncidium `Gower Ramsey`. Plant Growth Regul., 34: 229-232.
CrossRef - Chen, J.T. and W.C. Chang, 2006. Direct somatic embryogenesis and plant regeneration from leaf explants of Phalaenopsis amabilis. Biol. Plantarum., 50: 169-173.
CrossRef - Chung, H.H., J.T. Chen and W.C. Chang, 2007. Plant regeneration through direct somatic embryogenesis from leaf explants of Dendrobium. Biol. Plant., 51: 346-350.
CrossRefDirect Link - Diaz, M.D.S.S. and C.C. Alvarez, 2009. Plant regeneration through direct shoot formation from leaf cultures and from protocorm-like bodies derived from callus of Encyclia mariae (Orchidaceae), a threatened Mexican orchid. In vitro Cell. Develop. Biol. Plant, 45: 162-170.
CrossRefDirect Link - Engelmann, F., 2004. Plant cryopreservation: Progress and prospects. In Vitro Cell. Dev. Biol. - Plant, 40: 427-433.
CrossRefDirect Link - Gonzalez-Arnao, M.T., C.E. Lazaro-Vallejo, F. Engelmann, R. Gamez-Pastrana, Y.M. Martinez-Ocampo, M.C. Pastelin-Solano and C. Diaz-Ramos, 2009. Multiplication and cryopreservation of vanilla (Vanilla planifolia Andrews). In vitro Cell. Dev. Biol. Plant, 45: 574-582.
CrossRefDirect Link - Gonzalez-Benito, M.E. and C. Martin, 2011. In vitro preservation of Spanish biodiversity. In Vitro Cell. Dev. Biol. Plant, 47: 46-54.
CrossRefDirect Link - Halmagyi, A. and C. Deliu, 2007. Cryopreservation of carnation (Dianthus caryophyllus L.) shoot tips by encapsulation-vitrification. Sci. Hortic., 113: 300-306.
CrossRefDirect Link - Hirano, T., T. Godo, M. Mii and K. Ishikawa, 2005. Cryopreservation of immature seeds of Bletilla striata by vitrification. Plant Cell Rep., 23: 534-539.
CrossRefDirect Link - Hirano, T., T. Godo, K. Miyoshi, K. Ishikawa, M. Ishikawa and M. Mii, 2009. Cryopreservation and low-temperature storage of seeds of Phaius tankervilleae. Plant Biotechnol. Rep., 3: 103-109.
CrossRef - Hong, P.I., J.T. Chen and W.C. Chang, 2008. Plant regeneration via protocorm-like body formation and shoot multiplication from seed-derived callus of a maudiae type slipper orchid. Acta Physiologiae Plantarum, 30: 755-759.
CrossRefDirect Link - Hossain, M.M., M. Sharma, J.A.T. da Silva and P. Pathak, 2010. Seed germination and tissue culture of Cymbidium giganteum Wall.ex Lindl. Scientia Hortic., 123: 479-487.
CrossRef - Huan, L.T., T. Takamura and M. Tanaka, 2004. Callus formation and plant regeneration from callus through somatic embryo structures in Cymbidium orchid. Plant Sci., 166: 1443-1449.
Direct Link - Huang, C.H. and J.P. Chung, 2011. Efficient indirect induction of protocorm-like bodies and shoot proliferation using field-grown axillary buds of a Lycaste hybrid. Plant Cell Tissue Organ Cult., 106: 31-38.
CrossRefDirect Link - Ishikawa, K., K. Harata, M. Mii, A. Sakai, K. Yoshimatsu and K. Shimomura, 1997. Cryopreservation of zygotic embryos of a Japanese terrestrial orchid Bletilla striata by vitrification. Plant Cell Rep., 16: 754-757.
CrossRefDirect Link - Islam, M.O., A.R.M.M. Rahman, S. Matsui and A.K.M.A. Prodhan, 2003. Effects of complex organic extracts on callus growth and PLB regeneration through embryogenesis in the Doritaenopsis orchid. Japan Agric. Res. Quart., 37: 229-235.
Direct Link - Janarthanam, B. and S. Seshadri, 2008. Plantlet regeneration from leaf derived callus of Vanilla planifolia Andr. In Vitro Cell. Dev. Biol. Plant., 44: 84-89.
CrossRefDirect Link - Kuo, H.L., J.T. Chen and W.C. Chang, 2005. Efficient plant regeneration through direct somatic embryogenesis from leaf explants of phalaenopsis `Little Steve`. In Vitro Cellular Dev. Biol. Plant, 41: 453-456.
CrossRefDirect Link - Lee, Y.I. and N. Lee, 2003. Plant regeneration from protocorm-derived callus of cypripedium formosanum. In vitro Cell. Dev. Biol. Plant, 39: 475-479.
CrossRef - Luo, J.P., Y. Wang, X.Q. Zha and L. Huang, 2008. Micropropagation of Dendrobium densiflorum Lindl. ex Wall. through protocorm-like bodies: Effects of plant growth regulators and lanthanoids. Plant Cell, Tissue Org. Culture, 93: 333-340.
CrossRef - Lurswijidjarus, W. and K. Thammasiri, 2004. Cryopreservation of shoot tips of Dendrobium walter oumae by encapsulation/dehydration. Sci. Asia, 30: 293-299.
Direct Link - Maneerattanarungroj, P., S. Bunnag and M. Monthatong, 2007. In vitro conservation of Cleisostoma areitinum (Rchb.f.) garay, rare thai orchid species by an encapsulation-dehydration method. Asian J. Plant Sci., 6: 1235-1240.
CrossRefDirect Link - Ming-Hua, Y. and H. Sen-Rong, 2010. A simple cryopreservation protocol of Dioscorea bulbifera L. embryogenic calli by encapsulation-vitrification. Plant Cell Tissue Organ Cult., 101: 349-358.
CrossRefDirect Link - Moges, A.D. R.A. Shibli and N.S. Karam, 2004. Cryopreservation of African violet (Saintpaulia ionantha Wendl.) shoot tips. In Vitro Cell. Dev. Biol. Plant, 40: 389-395.
Direct Link - Mok, M.C., R.C. Martin and D.W.S. Mok, 2000. Cytokinins: Biosynthesis metabolism and perception. In Vitro Cell Dev. Biol. Plant., 36: 102-107.
CrossRefDirect Link - Murthy, H.N. and A.N. Pyati, 2001. Micropropagation of Aerides maculosum Lindl. (Orchidaceae). In Vitro Cell Dev. Biol.-Plant, 37: 223-226.
CrossRef - Na, Y.H. and K. Kondo, 1996. Cryopreservation of tissue-cultured shoot primordia from shoot apices of cultured protocorms in Vanda pumila following ABA preculture and desiccation. Plant Sci., 118: 195-201.
CrossRefDirect Link - Nayak, N.R., S. Patnaik and S.P. Rath, 1997. Direct shoot regeneration from foliar explants of an epiphytic orchid,Acampe praemorsa (Roxb.) Blatter and McCann. Plant Cell Rep., 16: 583-586.
CrossRefDirect Link - Nikishina, T.V., A.S. Popov, G.L. Kolomeitseva and B.N. Golovkin, 2001. Cryopreservation of seeds of some tropical orchids. Dokl. Biochem. Biophys., 378: 231-233.
CrossRef - Nikishina, T.V., E.V. Popova, M.G. Vakhrameeva, T.I. Varlygina and G.L. Kolomeitseva et al., 2007. Cryopreservation of seeds and protocorms of rare temperate orchids. Rus. J. Plant Physiol., 54: 121-127.
CrossRef - Park, S.Y., H.N. Murthy and K.Y. Paek, 2003. Protocorm-like body induction and subsequent plant regeneration from root tip cultures of Doritaenopsis. Plant Sci., 164: 919-923.
CrossRefDirect Link - Paul, H., G. Gaigny and B.S. Sangwan-Norreel, 2000. Cryopreservation of apple (Malus x domestica Borkh.) shoot tips following encapsulation-dehydration or encapsulation-vitrification. Plant Cell Rep., 19: 768-774.
CrossRef - Pouzi, N.Z., X. Rathinam, J.J.J. Antony, R. Poobathy and S. Subramaniam, 2011. Early investigation on cryopreservation of Dendrobium sonia-28 using encapsulation-dehydration with modified Evan blue assay. Afr. J. Biotechnol., 10: 3534-3539.
Direct Link - Prasertsongskun, S. and R. Awaesuemae, 2009. Effect of additive substances and planting substrate on growth development of Aerides houlletiana Rchb.f. seedling by tissue culture. KKU Sci. J., 37: 320-324.
Direct Link - Puchooa, D., 2004. Comparison of different culture media for the in vitro culture of Dendrobium (Orchidaceae). Int. J. Agric. Biol., 6: 884-888.
Direct Link - Rahman, A.R.M., M.O. Islam, A.K.M. Azad-ud-doula and S. Ichihashi, 2004. Effect of complex organic Extracts on Plantlet growth in the Doritaenopsis Orchid. J. Agric. Res. Q., 38: 55-59.
Direct Link - Roy, J., S. Naha, M. Majumdar and N. Banerjee, 2007. Direct and callus-mediated protocorm-like body induction from shoot-tips of Dendrobium chrysotoxum Lindl. (Orchidaceae). Plant Cell Tissue Organ Cult., 90: 31-39.
CrossRefDirect Link - Scottez, C., E. Chevreau, N. Godard, Y. Arnaud, M. Duron and J. Dereuddre, 1992. Cryopreservation of cold-acclimated shoot-tips of pear in vitro cultures after encapsulation-dehydration. Cryobiology, 29: 691-700.
CrossRefDirect Link - Shatnawi, M.A., K.A. Johnson and F.R. Torpy, 2004. In vitro propagation and cryostorage of Syzygium francissi (Myrtaceae) by the encapsulation-dehydration method. In Vitro Cell. Dev. Biol. Plant, 40: 403-407.
CrossRefDirect Link - Sheelavanthmath, S.S., H.N. Murthy, B.P. Hema, E.J. Hahn and K.Y. Paek, 2005. High frequency of protocorm like bodies (PLBs) induction and plant regeneration from protocorm and leaf sections of Aerides crispum. Sci. Hortic., 106: 395-401.
CrossRefDirect Link - Shibli, R.A., 2000. Cryopreservation of black iris (Iris nigricans) somatic embryos by encapsulation-dehydration. Cryo Lett., 21: 39-46.
PubMed - Shibli, R., D. Haagenson, S. Cunningham, W. Berg and J. Volenec, 2001. Cryopreservation of alfalfa (Medicago sativa L.) cells by encapsulation-dehydration. Plant Cell Rep., 20: 445-450.
CrossRefDirect Link - Shimura, H. and Y. Koda, 2004. Micropropagation of Cypripedium macranthos var rebunense through protocorm-like bodies derived from mature seeds. Plant Cell Tissue Organ Cult., 78: 273-276.
CrossRefDirect Link - Suzuki, M., T. Akihama and M. Ishikawa, 2005. Cryopreservation of encapsulated gentian axillary buds following 2 step-preculture with sucrose and desiccation. Plant Cell Tissue Organ Cult., 83: 115-121.
CrossRef - Thammasiri, K. and L. Soamkul, 2007. Cryopreservation of Vanda coerulea Griff. ex Lindl. seeds by vitrification. Sci. Asia, 33: 223-227.
CrossRefDirect Link - Thomas, T.D. and A. Michael, 2007. High-frequency plantlet regeneration and multiple shoot induction from cultured immature seeds of Rhynchostylis retusa Blume., an exquisite orchid. Plant Biotechnol. Rep., 1: 243-249.
CrossRefDirect Link - Tokuhara, K. and M. Mii, 1993. Micropropagation of Phalenopsis and Doritaenopsis by culturing shoot tips of flower stalk buds. Plant Cell Rep., 13: 7-11.
CrossRef - Tsukazaki, H., M. Mii, K. Tokuhara and K. Ishikawa, 2000. Cryopreservation of Doritaenopsis suspension culture by vitrification. Plant Cell Rep., 19: 1160-1164.
CrossRef - Wang, Q., R. Gafny, N. Sahar, L. Sela, M. Mawassi, E. Tanne and A. Perl, 2002. Cryopreservation of grapevine (Vitis vinifera L.) embryogenic cell suspensions by encapsulation-dehydration and subsequent plant regeneration. Plant Sci., 162: 551-558.
CrossRefDirect Link - Wang, Q., J. Laamanen, M. Uosukainen and J.P.T. Valkonen, 2005. Cryopreservation of in vitro-grown shoot tips of raspberry (Rubus idaeus L.) by encapsulation-vitrification and encapsulation-dehydration. Plant Cell Rep., 24: 280-288.
CrossRef - Yin, M. and S. Hong, 2009. Cryopreservation of Dendrobium candidum Wall. ex Lindl. protocorm-like bodies by encapsulation-vitrification. Plant Cell Tissue Organ Cult., 98: 179-185.
CrossRef - Zainuddin, M., A.L. Julkifle, R. Pobathy, U.R. Sinniah and A. Khoddamzadeh et al., 2011. Preliminary analysis of cryopreservation of Dendrobium Bobby Messina orchid using an encapsulation-dehydration technique with Evans blue assay. Afr. J. Biotechnol., 10: 11870-11878.
Direct Link - Zhao, P., F. Wu, F.S. Feng and W.J. Wang, 2008. Protocorm-like body (PLB) formation and plant regeneration from the callus culture of Dendrobium candidum Wall ex Lindl. Vitro Cell. Dev. Biol.-Plant., 44: 178-185.
CrossRef