
M.M. Rahman
Institute of Biological Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysia
M. Tateyama
Faculty of Agriculture, University of Miyazaki, Miyazaki 889-2192, Japan
M. Niimi
Faculty of Agriculture, University of Miyazaki, Miyazaki 889-2192, Japan
R.B. Abdullah
Institute of Biological Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysia
W.E. Wan Khadijah
Institute of Biological Sciences, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysia
O. Kawamura
Faculty of Agriculture, University of Miyazaki, Miyazaki 889-2192, Japan
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 4 | Page No.: 586-589







ABSTRACT
Oxalate concentration in forage plants is important, because it results mineral deficiency in ruminants. Data on oxalate concentration in forage plants in conjunction with cutting and uncutting conditions throughout the growing period are limited. This study was aimed to investigate the changes in oxalate and some mineral concentrations of setaria (Setaria sphacelata). The plants were harvested at different stages (vegetative, boot, pre-flowering, flowering and seed) of maturity and at about 50 cm in length of regrowth (second to sixth cuttings) for evaluation of soluble oxalate, insoluble oxalate and some mineral concentrations. Soluble oxalate and total oxalate concentrations, as well as mineral concentrations, decreased with advancing maturity. Both oxalate concentrations (soluble or insoluble) were higher in leaf compared to stem. Soluble oxalate and total oxalate concentrations of regrowth were the highest at third cutting and lowest at sixth cutting. Insoluble oxalate concentration of regrowth was almost similar in all cuttings, except for the sixth cutting. The highest concentrations of potassium, sodium and magnesium of regrowth were observed at third cutting, while the highest concentration of calcium was observed at sixth cutting. A relationship between oxalate and mineral concentrations was partially observed. Results suggest that cutting materials of setaria from June to October could achieve oxalate levels that are toxic to ruminants.
PDF Abstract XML References Citation
Received: March 28, 2013;
Accepted: May 06, 2013;
Published: November 26, 2013
How to cite this article
M.M. Rahman, M. Tateyama, M. Niimi, R.B. Abdullah, W.E. Wan Khadijah and O. Kawamura, 2014. Changes in Oxalate and Some Mineral Concentrations of Setaria sphacelata Under Cutting and Uncutting Conditions. Pakistan Journal of Biological Sciences, 17: 586-589.
DOI: 10.3923/pjbs.2014.586.589
URL: https://scialert.net/abstract/?doi=pjbs.2014.586.589
DOI: 10.3923/pjbs.2014.586.589
URL: https://scialert.net/abstract/?doi=pjbs.2014.586.589
INTRODUCTION
Oxalate is an anti-nutritional nutrient in forage plants that affects mineral utilization in ruminants (De Carvalho et al., 2011). It can bind with some minerals and form insoluble oxalate, which are not readily available for ruminants. Evensen and Standal (1984) reported that tropical plants accumulate more oxalate than temperate plants. Among the tropical forage plants, setaria (Setaria sphacelata) contains moderate to high levels of oxalate (Rahman et al., 2013).
Oxalate concentration in forage plants is affected by many internal and external factors (Rahman and Kawamura, 2011). For example, the oxalate concentration was much higher in leaves than in stems and this concentration declined rapidly in both leaf and stem parts with increasing age (Middleton and Barry, 1978; Rahman et al., 2006). Oxalate concentrations were also observed higher in autumn and winter seasons compared to other seasons when plant growth rates were low (Middleton and Barry, 1978). In contrast, plants grown in early summer showed higher oxalate concentration when compared to plants grown later in the season (Rahman et al., 2006). Rahman et al. (2008) reported that oxalate concentration has relationship with some mineral concentrations in Napier grass (Pennisetum purpureum), and it varies not only between forage plants, but also varies according to the soil, water and climatic conditions where the forage is grown (Rahman and Kawamura, 2011). Since oxalate concentration varies with advancing season, it is necessary to investigate the oxalate concentration of setaria throughout the growing period. The aim of this study was to investigate the changes in oxalate and some mineral concentrations of setaria (cultivar: Splenda) under cutting and uncutting conditions in Kyushu, Japan.
MATERIALS AND METHODS
Experimental site and planting management: Plot was established from the overwintered setaria (cultivar: Splenda) in an Experimental Field of Faculty of Agriculture, University of Miyazaki, Japan. Climatic conditions during the plant growth period (May to Nov., 2010) were recorded at the Miyazaki Meteorological Station (Latitude 31°56.3'N Longitude 131°24.8'E) about 10 km north from the experimental site. Average temperature and monthly precipitation during the plant growth period were 19.9°C and 253.8 mm, respectively, with maximum precipitation (778.5 mm) in June. The mean maximum and minimum temperatures were 32.1 and 9.1°C, recorded in August and November, respectively. The soil type of the plot was a sandy soil. At the early stage of plant growth (after overwinter), the plot received nitrogen (N), phosphorus (P2O5) and potassium (K2O) (60 kg h-1 of each element) as NPK compound fertilizer in May 2010.
Treatments and experimental design: To follow the maturity, the plot was split into subplot for five stages of cutting with five replicates (1) Vegetative (50 cm in plant length), (2) Boot, (3) Pre-flowering (5-10% flowering), (4) Flowering and (5) Seed. To follow the botanical fractions, plant at pre-flowering stage was harvested with five replicates and separated into leaf (leaf blade) and stem (stem with leaf sheath). To follow the regrowth, plants regrown from first cutting subplot at vegetative stage were sampled with five replicates on five occasions (when the plant length was about 50 cm): 28 June, 16 July, 10 Aug., 13 Sept. and 4 Nov. Summary of sampling date and plant length are presented in Table 1.
Chemical analysis: Approximately 400 g samples for each replication were dried at 70°C for 48 h. The dried samples were ground in a Wiley mill through a 1 mm screen.
| Table 1: | Summary of sampling date and plant length |
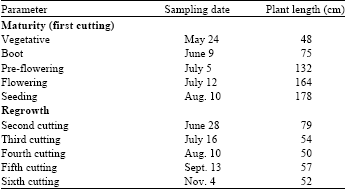 | |
These samples were analyzed for soluble and total oxalates concentration by the method of Rahman et al. (2007). Insoluble oxalate was estimated by subtracting soluble oxalate from total oxalate. Concentrations of Ca, magnesium (Mg), potassium (K) and sodium (Na) in samples were determined by flame atomic spectroscopy after wet digestion with nitric acid and hydrogen peroxide (Laboratory of Agricultural Chemistry the University of Tokyo, 1978).
Statistical analysis: Data of changes in oxalate and some mineral concentrations were analyzed using the General Linear Model (GLM) procedure of SPSS (version 12.0, SPSS Inc., Chicago, IL, USA). When the F-test was significant (p<0.05), Least Significant Difference (LSD) test for comparisons was used to compare means.
RESULTS AND DISCUSSION
Uncutting condition: The effect of uncutting condition on oxalate and mineral concentrations in setaria is presented in Table 2. Soluble oxalate concentration decreased gradually with advancing maturity. It was the highest (36.2 g kg-1) at vegetative stage and the lowest (13.1 g kg-1) at the seed stage. Insoluble oxalate concentration was also affected by maturity, but no trend was observed among the different stages. Similar to soluble oxalate concentration, insoluble oxalate concentration was the highest (9.6 g kg-1) at the vegetative stage and the lowest (1.5 g kg-1) at the seed stage. In most of the cases, the concentrations of K, Na, Ca and Mg decreased gradually with advancing maturity. Maturity is usually considered to be the primary factor affecting the chemical composition and nutritive value of forage. The present findings are in line with the findings of Middleton and Barry (1978) who observed that the oxalate concentration of Setaria splendida declined rapidly with increasing age. Rahman et al. (2009) also observed that the oxalate concentration declined as the harvest interval increased.
| Table 2: | Effect of uncutting on oxalate and mineral concentrations (g kg-1) |
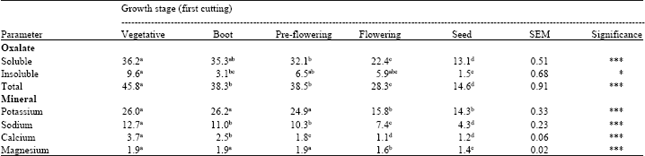 | |
| SEM: Standard error of the mean; ***p<0.001; *p<0.05, Means in the same row with different superscripts differ significantly at p<0.05 | |
Much of this decline could be due to a dilution effect caused by higher yield as the harvest interval increased. In addition, this decline with age would also be enhanced by an increasing proportion of stem as the proportion of leaf, as stems contain less oxalate than leaves (Rahman et al., 2006).
Even though oxalate declined with advancing maturity, setaria still maintained toxic levels of oxalate (more than 30 g kg-1 soluble oxalate) until the pre-flowering stage. Setaria exhibited a rapid decrease in oxalate and mineral concentrations during the flowering and seed stages compare to other growth stages (vegetative, boot or pre-flowering). Setaria also contained low amount of Ca (1.1-3.7 g kg-1), it is suggested that bioavailability of Ca from this plant is low due to interactions between nutrients and antinutrients (especially oxalate) in the feed. Due to the low solubility of Ca oxalate, extremely little soluble Ca can exist in forages in the presence of oxalic acid. Because of its low solubility, the Ca in Ca-oxalate is poorly available to the grazing herbivore, passes through the digestive tract and is excreted in the faeces (Ward et al., 1979).
Botanical fractions: The effect of botanical fractions on oxalate content in setaria at pre-flowering stage is shown in Table 3. Leaf contained higher soluble oxalate (43.1 vs. 17.8 g kg-1), insoluble oxalate (9.9 vs 0.5 g kg-1) and total oxalate (53.0 vs. 18.3 g kg-1) compared to stem. This result is in agreement with the findings of Rahman et al. (2006) who reported that leaf of napiergrass exhibited higher oxalate concentration than stem. Similar result was also found in kikuyugrass (Pennisetum clandestinum) (Marais et al., 1997).
Cutting condition: The soluble and total oxalate concentrations of regrowth were the highest (45.9 and 56.7 g kg-1) at third cutting and the lowest (30.5 and 37.2 g kg-1) at sixth cutting, respectively (Table 4). The insoluble oxalate concentration of regrowth was almost similar in all cuttings, except for the sixth cutting. The effect of regrowth on mineral concentration in setaria is presented in Table 4. The concentrations of K, Na, Ca and Mg in regrowth plant were affected by cutting. The highest concentrations of K (43.7 g kg-1), Na (22.6 g kg-1) and Mg (3.8 g kg-1) of regrowth were observed at third cutting and the highest concentration of Ca (4.5 g kg-1) of regrowth was observed at sixth cutting. There are limited data regarding the influence of regrowth on oxalate concentration, particularly when no significant changes in the leaf: stem ratio occur. Since leaf contains higher oxalate than stem, oxalate concentration declines with age due to increasing proportion of stem compared to leaf. Results of this study showed that oxalate and mineral concentrations of regrowth were not similar among all the cuttings (cut when the plant was 50 cm in length), though proportion of leaf and stem was not measured. This change may be associated with advancing season, because many environmental effects associated with seasonal change which may alter oxalate levels (e.g., temperature, precipitation, day length, hours of sunlight, floral induction etc.,) (Rahman and Kawamura, 2011). Rahman et al. (2006) reported that oxalate concentration affected by the season with the highest value being associated with early summer samples and the lowest value with late autumn samples.
Setaria is widely grown for grazing by dairy and beef cattle. In most of the cases, pastures usually graze when the grass is about 30-50 cm high. In the present study, all regrowth plants contained more than 30 g kg-1 soluble oxalate when the grass was about 50-60 cm in length and these oxalate levels can cause toxic to ruminants. Radostits et al. (2000) reported that acute oxalate poisoning and death have been observed in cattle eating Setaria sphacelata.
Relationship between oxalate and mineral concentration in plants: Table 5 represents the correlation coefficient between the oxalate and mineral concentrations in plants. For maturity (first growth), the soluble and total oxalate concentrations were positively correlated with each type of cation (K, Na, Ca and Mg) content, while the insoluble oxalate concentrations was positively correlated with the Na and Ca concentrations.
| Table 3: | Effect of botanical fractions on oxalate concentrations (g kg-1) at pre-flowering stage |
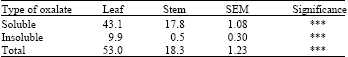 | |
| SEM: Standard error of the mean, ***p<0.001 | |
| Table 4: | Effect of cutting on oxalate and mineral concentrations (g kg-1) |
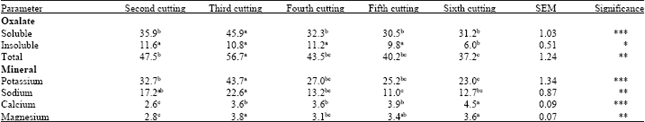 | |
| SEM: Standard error of the mean, *p<0.05; **p<0.01; ***p<0.001; Means in the same row with different superscripts differ significantly (p<0.05) | |
| Table 5: | Correlation coefficients between oxalate and mineral concentrations under uncutting and cutting conditions |
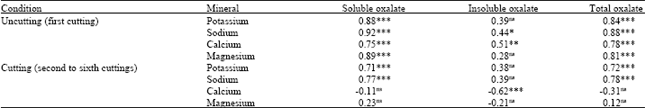 | |
| ns: Not significant (p>0.05), *p<0.05, **p<0.01; ***p<0.001 | |
For regrowth (second to sixth cuttings), the soluble and total oxalate concentrations were positively correlated with the K and Na concentrations, while the insoluble oxalate concentration was negatively correlated with the Ca concentration. This result agreed partially with Rahman et al. (2008) who observed that the soluble oxalate concentration in Napier grass was positively correlated with K concentration, while the insoluble oxalate concentration was positively correlated with Ca and Mg concentrations. The results of the present study suggest that the relationship between oxalate and mineral concentrations may differ with growth stages of development of plant.
CONCLUSION
It is concluded that oxalate and mineral concentrations declined gradually with advancing maturity. With regard to the botanical fractions, oxalate concentration was higher in leaf compared to stem. Oxalate and mineral concentrations were also affected by the regrowth plant. A relationship between oxalate and mineral concentrations was partially observed. All stages of growth (except for flowering and seed stages) or regrowth of setaria showed more than 30 g kg-1 soluble oxalate, which may be considered harmful for ruminant.
REFERENCES
- De Carvalho, P.R., E.M.C. Villalobos, P.A.F. de Castilho, J.E. Loureiro, P.R. dos Santos Mello and L.C. da Silva, 2011. Screening to prevent to carential and metabolic disease and HPTNS of equids grazing forage grasses with unbalanced levels of minerals, through the mineral profile and creatinine clearance ratio for Ca and P assessment. Pak. J. Nutri., 102: 519-538.
CrossRefDirect Link - Middleton, C.H. and G.A. Barry, 1978. A study of oxalate concentration in five grasses in the wet tropics of Queensland. Trop. Grasslands, 12: 28-35.
Direct Link - Rahman, M.M., M. Niimi, Y. Ishii and O. Kawamura, 2006. Effects of season, variety and botanical fractions on oxalate content of napiergrass (Pennisetum purpureum Schumach). Grassland Sci., 52: 161-166.
CrossRefDirect Link - Rahman, M.M., M. Niimi and O. Kawamura, 2007. Simple method for determination of oxalic acid in forages using high-performance liquid chromatography. Grassl. Sci., 53: 201-204.
CrossRef - Rahman, M.M., Y. Ishii, M. Niimi and O. Kawamura, 2008. Effects of levels of nitrogen fertilizer on oxalate and some mineral contents in napiergrass (Pennisetum purpureum Schumach). Grassl. Sci., 54: 146-150.
CrossRef - Rahman, M.M., Y. Ishii, M. Niimi and O. Kawamura, 2009. Effect of clipping interval and nitrogen fertilisation on oxalate content in pot-grown napier grass (Pennisetum purpureum). Trop. Grassl., 43: 73-78.
Direct Link - Rahman, M.M. and O. Kawamura, 2011. Oxalate accumulation in forage plants: Some agronomic, climatic and genetic aspects. Asian-Aust. J. Anim. Sci., 24: 439-448.
CrossRefDirect Link - Rahman, M.M., M. Ikeue, M. Niimi, R.B. Abdullah, W.E. Wan Khadijah, K. Fukuyama and O. Kawamura, 2013. Case study for oxalate and its related mineral contents in selected fodder plants in subtropical and tropical regions. Asian J. Anim. Vet. Adv., 8: 535-541.
CrossRefDirect Link - Ward, G., L.H. Harbers and J.J. Blaha, 1979. Calcium-containing crystals in Alfalfa: Their fate in cattle. J. Dairy Sci., 62: 715-722.
CrossRef