
Md. Sarower-E-Mahfuj
Department of Fisheries Biology and Genetics, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh
M. Belal Hossain
Biology Group, Faculty of Science, Universiti of Brunei Darussalam, BE1410, Brunei
M. Mokhlesur Rahman
Department of Fisheries Biology and Genetics, Bangladesh Agricultural University,
Mymensingh-2202, Bangladesh
Md. Imran Hoshan
Department of Fisheries Biology and Genetics, Bangladesh Agricultural University, Mymensingh-2202, Bangladesh
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 4 | Page No.: 490-496







ABSTRACT
Chromosomal and karyotypic studies are required for genetic improvement of any organisms. This study was performed to identify individual chromosomes on morphological basis and to characterize a standard karyotype using the fish, Rohu (Labeo rohita, Hamilton, 1822). Colchicine (0.05%) treated (2, 2.5 and 3 h) tissues of two day-old larvae were used for slide preparations and selected plates were photomicrographed under high resolution research microscope. Slide preparations were done following hydrolysis (10% HCl), mordanting (2% iron alum) and staining with haematoxylin. Colchicine treatment for 2 h gave satisfactory results in respect of degree of contraction of the chromosomes. Chromosome number 2n = 50 was counted at metaphase stage. Measurements of the chromosomes were taken from the selected plates based on morphologically distinct condition. Standard haploid karyotype was formulated following the combined scatter diagram technique. Six individually identifiable and 19 not individually identifiable chromosomes consisted of the haploid complement. The six individually identifiable ones consisted of 2 m+3 sm+1st and not individually identifiable ones consisted of 16 m+3 sm chromosomes. This study report may provide complete report on chromosomal and karyotype knowledge in L. rohita and suggest the genetic purity of L. rohita may contributes to sustainable aquaculture production.
PDF Abstract XML References Citation
Received: April 10, 2013;
Accepted: May 08, 2013;
Published: November 26, 2013
How to cite this article
Md. Sarower-E-Mahfuj, M. Belal Hossain, M. Mokhlesur Rahman and Md. Imran Hoshan, 2014. Chromosomal Studies and Quantitative Karyotypic Analysis of Rohu, Labeo rohita. Pakistan Journal of Biological Sciences, 17: 490-496.
DOI: 10.3923/pjbs.2014.490.496
URL: https://scialert.net/abstract/?doi=pjbs.2014.490.496
DOI: 10.3923/pjbs.2014.490.496
URL: https://scialert.net/abstract/?doi=pjbs.2014.490.496
INTRODUCTION
Rohu, Labeo rohita is one of the important members of the family cyprinidae which is distributed in native river systems of Bangladesh, India, Pakistan and Myanmar (Talwar and Jhingran, 1991). In Bangladesh, this species is mostly found in the Padma-Brahmaputra river systems and their tributaries and in the Halda river of Chittagong region (Alam et al., 2009). Rohu is a pioneer candidate among Indian Major Carps which contribute 24.84% of the total fish production in Bangladesh (DoF, 2011). Recently the stock of Rohu population has been deteriorated through uncontrolled interspecific and intergeneric hybridization and inbreeding (Sarder, 2007; Khan et al., 2006). As a result the chromosomal and genetic constitution is altered resulting in deviation from optimal growth and the purity of Rohu stocks is being gradually lost. It is important to reveal the chromosomal and karyotypic condition in wild and hatchery Rohu populations to identify pure and hybrid Rohu, to prevent loss of genetic purity and protect them from potentially vulnerable to rapid deterioration.
Chromosomes are considered as the physical basis of heredity because they have a special organization and are capable of self reproduction. From the viewpoint of genetic control, the rapid production of inbred lines and cytotaxonomy, karyotypic and chromosomal studies are important for fish breeding (Thorgaard and Disney, 1990; Kirpichnikov, 1981). This type of studies provide main information on the number, size and morphology of chromosomes (Tan et al., 2004) which is needed to undertake chromosome manipulation works (Khan et al., 2000). In the last few decades works have been focused on the field of cytogenetic investigation of fishes, especially in the area of systematics, mutagenesis and aquaculture (Sofy et al., 2008).
In view of the above, L. rohita is a good candidate for genetic investigations such as hybridization and ploidy manipulation. The present investigation aims to provide the detailed information on the chromosome number and quantitative karyotype of this species.
MATERIALS AND METHODS
Sample collection and processing: Two day-old alive, larvae were collected from “Brahmaputra Fish Seed Complex”, Shambugonj, Mymensingh, Bangladesh. The larvae were transported to the Cytogenetics laboratory, Department of Genetics and Plant Breeding, Bangladesh Agricultural University, Mymensingh. For chromosomal studies and quantitative karyotypic analysis the method of Ahmad et al. (1983) was followed with some modifications.
The stock solution of colchicine was made by dissolving 0.05 g colchicine in 100 mL distilled water. Healthy and vigorously growing larvae were allowed to swim in 0.05% colchicine solution. Then the larvae were kept in a glass petri dish containing 0.7% NaCl for 15 min at 28°C room temperature. Then the larvae were transferred to another petri dish containing distilled water kept for 15 min. The larvae were transferred into a vial containing freshly prepared fixative solution (acetic acid: ethanol, 1:3).
Slide preparation: Three batches of fixation, 2, 2.5 h, were processed separately. Fixed larvae tissues were hydrolyzed in small vials with 10% HCl at 60°C in an incubator for 2-3 min. Before actual hydrolysis of the tissues 10% HCl was kept in the incubator at 60°C for one hour in a tightly stoppered vial. Fixed tissues were then kept in the same preheated HCl solution in the incubator. Then hydrolized tissues were washed in fresh water for 2-3 times. The tissues were then treated with 2% aqueous solution of iron alum (Ferric ammonium sulphate) for about 7 min for mordanting. Iron alum treated tissues were then washed for 2-3 times with fresh water and stained in 0.5% haematoxylin for 8 min. The stained tissues were kept in water. Slides were prepared within 2-3 h of staining and microscopic observation. Observation: Then mitotic dividing cells were observed among the spreading tissues by using (Olympus microscope, CHD-64M061, Japan). Photomicrographs of the selected chromosome plates were taken with the aid of Olympus research microscope (Olympus-BX40, Japan) using plan 100 X oil immersion objective. Chromosome measurements in millimeters (mm) were made from photo prints and chromosome length were converted to micron (μ) based on calculated final print magnification.
Statistical analyses: Arm ratios of the chromosomes were calculated by dividing the length of the long arm by that of the short arm (L/S). The conventions proposed by Levan et al. (1964) were used to refer to the different types of chromosomes. Scatter diagram using the arm ratio and total length of the individual chromosomes at each of the selected plates were required. Homologous pairs of chromosomes were determined based on the proximity of two points and haploid values of the chromosomes were calculated. Thus a standard karyotype of L. rohita was developed following three steps of analysis: Firstly, a scatter diagram was produced for all chromosomes in each plate separately, by use of which the diploid number of chromosomes were reduced to the haploid number and haploid values of the chromosomes were determined. Secondly, a combined scatter diagram of the haploid complements of all plates was constructed to establish a standard morphology of those chromosomes which could be identified. Thirdly, those chromosomes which could not be identified individually were characterized through probabilistic inferences.
In a combined scatter diagram six groups of points could be delineated, representing six individually identifiable chromosomes. Individually identified groups were encircled by boundaries around those chromosomes. Each group of points included four haploid values, one point of each plate. The mean and standard deviation were measured in terms of total length and arm ratio of four haploid values of each plate of a group boundary. Thus a group boundary represented a haploid identified chromosome. Unidentified chromosomes were classified according to length and arm ratio. These unidentified chromosomes were distributed to the various morphological classes. These were determined by probabilistic inferences based on the frequency in a given class which were occurred in the combined scatter diagram. Finally, all chromosomes in the haploid complement were numbered in decreasing order of length and increasing order of arm ratio.
RESULTS
Clearly, three plates were found at late prophase stage, seven plates were found in highly contracted condition of the chromosome and four plates were found at prometaphase stage which were used in karyotypic studies. Colchicine (0.05%) treatment for 2 h gave satisfactory results in respect of degree of contraction of the chromosomes and frequency of prometaphase plate. The fixative acetic acid and ethyl alcohol (1:3) was found suitable for the present materials. Hydrolysis of the body tissues in 10% HCl for 2-3 min at 60°C was found adequate to soften the specimen. Mordanting of the hydrolysed tissues for 7 min in 2% iron alum solution and staining for 8 min in 0.5% haematoxylin produced adequate degree of staining of chromosomes. Present experiment revealed that the somatic chromosome number of L. rohita was 2n = 50. Late prophase stage was observed; the nuclear envelope breaks up and the chromosomes were condensed. This is called super coiling (Fig. 1a). Exclusively contracted metaphase chromosomes were also observed in the body tissues which were unsuitable for measurement (Fig. 1b). A representative plate of the four plates used in the karyotype analysis is shown in Fig. 1c.
Based on satisfactory staining and spreading four metaphase plates were selected for karyotype analysis. From each of the single plate scatter diagram 25 homologous chromosome pairs were determined by encircling the two proximal points (Fig. 2).
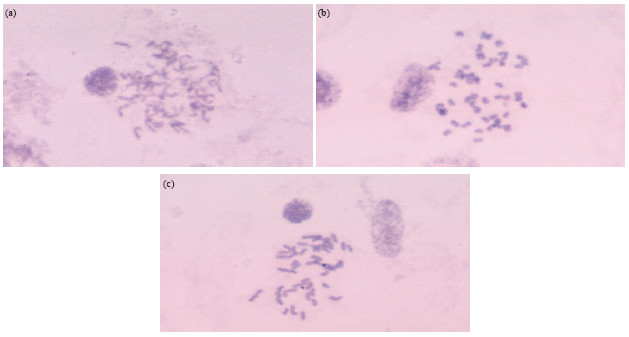 | |
| Fig. 1: | (a) Complement of two day-old larvae of L. rohita at late prophase stage, showing thickly stained chromosome segments, (b) Chromosomes highly contracted. Chromosomes at stages have shown 1 and 2 not used in karyotypic analysis and (c) Representative complement of larvae tissue included for karyotype analysis. All X 100 |
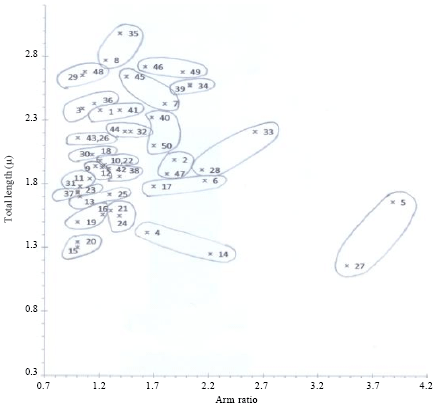 | |
| Fig. 2: | Scatter diagram of a representative plate of L. rohita. Each pair of points encircled was considered to represent a homologous pair |
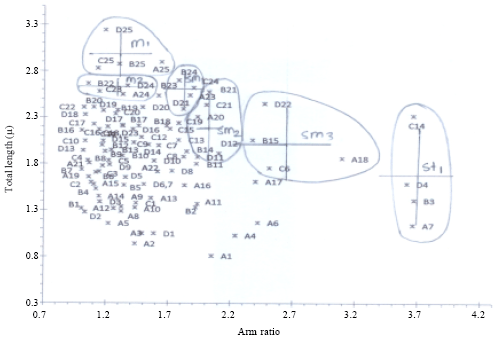 | |
| Fig. 3: | Scatter diagram of the 25 haploid chromosomes values, lengths and arm ratios, from each four plates of L. rohita; points A1-A25 for plate A; points B1-B25 for plate B; points C1-C25 for plate C and points D1-D25 for plate D. Group boundaries were drawn around those chromosomes which were recognizably homologous from each of the four plates. Intersecting lines within the groups represent the group mean of total length and arm ratio±one standard deviation for either variable of that group. Epithets m1, sm1, st1 etc. within the groups are the specific names of the chromosomes concerned |
| Table 1: | Mean lengths (μ) and mean arm ratios of the identified chromosomes of L. rohita |
 | |
The comparability of measurements of chromosomes in the four plates were determined by examining the variation in haploid total lengths among the plates included. The haploid total length ranged from 38.91-52.94 μ with a mean of 48.01±2.69 μ and a coefficient of variation of 5.60%. Thus, there was a similar degree of contraction of the chromosomes in the four plates.
Corresponding chromosomes in different haploid complements were determined through a grouping technique applied to the combined scatter diagram of the four haploid compliments involving 100 haploid chromosomes shown in Fig. 3. Each point in the scatter diagram represented a specific chromosome in a particular haploid complement. Symbols in the diagram refer to specific chromosomes in a particular haploid complement and the adjacent letters A, B, C and D refers to the four plates studied, respectively and numbers 1-25 represent the individual haploid chromosomes. Thus one highest point on the Y axis, D25 represented the haploid chromosome number 25 in plate D. Each cluster must contain one point (chromosome) from each plate studied (Plates A-D).
Six groups of points could be recognized (Fig. 3). Each group must contain four points belonging to four plates. These groups were clearly distinct. The mean, standard error and coefficient of variation were determined for length and arm ratio, using original diploid values. The six identified chromosomes comprised 2 ‘m’, 3 ‘sm’ and 1 ‘st’ types. The chromosomes within each type were numbered in decreasing order of mean length. Chromosome type together with this number constitutes the specific name of the chromosome concerned in Table 1. The six identified chromosomes occupied approximately 28.61% of the total complement length and this was consistent across four plates shown in Table 2.
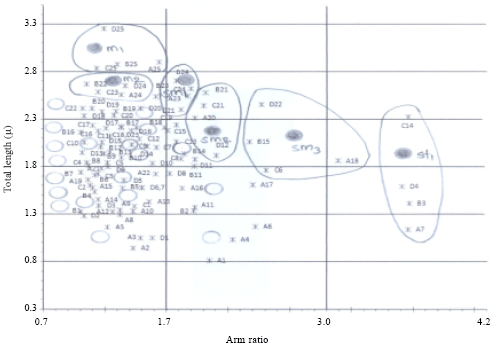 | |
| Fig. 4: | Locations of L. rohita chromosomes in a two-dimensional array, based on total length and arm ratio. Solid circles represent identified chromosomes, open circles represent unidentified chromosomes |
| Table 2: | Proportion of the complement total length occupied by the six identified chromosomes in L. rohita |
 | |
| Table 3: | The allocation of unidentified chromosomes in L. rohita karyotype to different morphological categories |
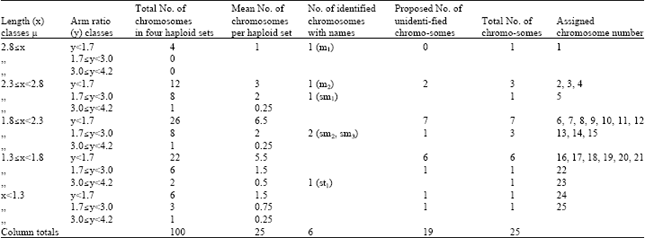 | |
All haploid chromosomes were classified according to length and arm ratio shown in Table 3. Chromosome nomenclature was done based on the categories of class interval of total length and arm ratio. This classification was applied to the scatter diagram of the haploid complements as a grid of length and arm ratio classes were shown in Fig. 4. Length was used in plotting which resulted in vertical displacement of the points.
| Table 4: | Standard haploid karyotype of chromosomes with chromosome type |
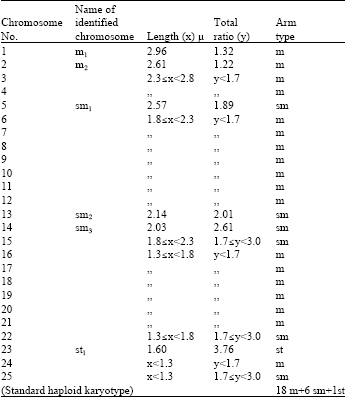 | |
The unidentified chromosomes were distributed to the various morphological classes using probabilistic inferences based on the frequency of chromosomes in a given class per haploid set in column 4 of Table 3, occurrence of points in the combined scatter diagram in (Fig. 4) and the original data and arm ratio of the chromosomes. The number of unidentified chromosomes allocated to the various morphological classes is given in column 6 of Table 3. Finally, all 25 chromosomes in the haploid complement were numbered from 1 to 25 in decreasing order of length and increasing order of arm ratio within a length class shown in column 8 of Table 3. Thus each chromosome was allocated a serial identification number and each of the six chromosomes identified individually also carried a specific name based on its arm ratio. The morphological features of the chromosomes are summarized in Table 4. The proposed standard karyotype consisted of 18 ‘m’, 6 ‘sm’ and 1 ‘st’ chromosome.
DISCUSSION
The chromosomes are considerably shorter, 0.75 μ to 3.48 μ in total length. The present findings of 2n = 50 chromosomes in L. rohita is in agreement with the earlier reports (Tripathy et al., 2010; Nagpure et al., 2001; Khuda-Bukhsh and Tiwary, 1994). During slide preparation bringing the chromosomes in the same focal plane of the objective lens as well as spreading of the chromosomes posed a major problem. Cells tend to suffer breakage easily causing unavailability of intact chromosome complements. Because the small size and higher number of chromosome of the complement is further disadvantageous for cytological studies in the present material. To attain such condition one or more chromosomes would tend to break and overlap apart in the centromere region. Therefore, utmost care had to be exercised and practiced during slide preparation and selection of chromosome plates in undistorted condition. However, cell preparations for such observations must be satisfactory enough so as to avoid any misconception.
Standard haploid karyotype is formulated in the present research consisted of 18m+6sm+1st. The karyotype study of L. rohita has been described previously by some authors. Tripathy et al. (2010) described the karyotype composition as 8m+12sm+22st+8t/a; Nagpure et al. (2001) depicted the karyotype composition as 10m+14sm+8st+18t and FN = 74; Khuda-Bukhsh and Tiwary (1994) observed as 18m+8sm+24t and FN = 76; all of these studies reported the diploid number to be 50.
The variation of result is dependent on several causes such as the methods of chromosome preparation, staining procedure, tissue source of the body where the dividing cells are found. In the present study the methodology of chromosome preparation was different from previous studies. The result varied due to the degree of contraction and spreading of chromosomes. Probably the difference was due to variation in the degree of contraction of the chromosomes and methodological constraints arising from the small size of chromosomes. Fish karyotypes are not identical, as in human being and other plant and animal species, so a researcher cannot have a standard karyotype for fish because not only there are differences between species, polymorphism often occurs within the same fish species (Al-Sabti, 1991). The problem encountered is the morphological variation that exists even between homologous chromosomes in the same nucleus (Levan et al., 1964; Al-Sabti, 1991).
CONCLUSION
Chromosomal and karyotype have become an active area of research in fish genetics. This study report may provide a complete report on chromosomal and karyotype knowledge in L. rohita and suggest the genetic purity of L. rohita which may contributes to sustainable aquaculture production. The quantitative method used in this study may be appropriate for other fish species which may present similar cytological difficulties and may encourage further work on the cytogenetics especially in cytology and karyotype analysis.
REFERENCES
- Alam, M.S., M. Jahan, M.M. Hossain and M.S. Islam, 2009. Population genetic structure of three major river populations of Rohu, Labeo rohita (Cyprinidae: Cypriniformes) using microsatellite DNA markers. Genes Genom., 31: 43-51.
CrossRef - Khan, T.A., M.P. Bhise and W.S. Lakra, 2000. Chromosome manipulation in fish-a review. Indian J. Anim. Sci., 70: 213-221.
Direct Link - Khan, M., M. Rahman, M.S. Alam and M.M.H. Bhuiyan, 2006. Allozyme variation of hatchery and river populations of rohu (Labeo rohita, Hamilton) in Bangladesh. Aquacult. Res., 37: 233-240.
CrossRef - Levan, A., K. Fredga and A.A. Sandberg, 1964. Nomenclature for centromeric position on chromosomes. Hereditas, 52: 201-220.
CrossRefDirect Link - Nagpure, N.S., B. Kushwaha, S.K. Srivastava and A.G. Ponniah, 2001. Comparative cytogenetic studies in Indian major carps Labeo rohita, Catla catla and Cirrhinus mrigala (Cyprinidae, Pisces). Chromosome Sci., 5: 153-158.
Direct Link - Sofy, H.L., A.M. Layla and M.K.A. Iman, 2008. Karyotypic diversity of some tilapia species. Nat. Sci., 6: 19-27.
Direct Link - Tan, X., J. Qin, B.N. Chen, L. Chen and X. Li, 2004. Karyological analyses on redclaw crayfish Cherax quadricarinatus (Decapoda: Parastacidae). Aquaculture, 234: 65-76.
Direct Link