
Nwe Nwe Htwe
Institute of BioScience, Universiti Putra Malaysia, 43400, UPM Serdang, Selangor, Malaysia
Ho Chai Ling
Department of Cell and Molecular Biology, Faculty of Biotechnology and Biomolecular, Science Universiti Putra Malaysia, 43400, UPM Serdang, Selangor, Malaysia
Faridah Qamaruz Zaman
Department of Biology, Faculty of Science, Universiti Putra Malaysia, 43400, UPM Serdang, Selangor, Malaysia
Mahmood Maziah
Department of Biochemistry, Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400, UPM Serdang Selangor, Malaysia
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 4 | Page No.: 472-481







ABSTRACT
Rice is one of the most important cereal crops with great potential for biotechnology progress. In transformation method, antibiotic resistance genes are routinely used as powerful markers for selecting transformed cells from surrounding non-transformed cells. In this study, the toxicity level of hygromycin was optimized for two selected mutant rice lines, MR219 line 4 and line 9. The mature embryos were isolated and cultured on an MS medium with different hygromycin concentrations (0, 20, 40, 60, 80 and 100 mg L-1). Evidently, above 60 mg L-1 was effective for callus formation and observed completely dead. Further there were tested for specific concentration (0-60). Although, 21.28% calli survived on the medium containing 45 mg L-1 hygromycin, it seemed suitable for the identification of putative transformants. These findings indicated that a system for rice transformation in a relatively high frequency and the transgenes are stably expressed in the transgenic plants. Green shoots were regenerated from the explant under hygromycin stress. RT-PCR using hptII and gus sequence specific primer and Southern blot analysis were used to confirm the presence of the transgene and to determine the transformation efficiency for their stable integration in regenerated plants. This study demonstrated that the hygromycin resistance can be used as an effective marker for rice transformation.
PDF Abstract XML References Citation
Received: March 12, 2013;
Accepted: May 08, 2013;
Published: November 26, 2013
How to cite this article
Nwe Nwe Htwe, Ho Chai Ling, Faridah Qamaruz Zaman and Mahmood Maziah, 2014. Plant Genetic Transformation Efficiency of Selected Malaysian Rice Based on Selectable Marker Gene (hptII). Pakistan Journal of Biological Sciences, 17: 472-481.
DOI: 10.3923/pjbs.2014.472.481
URL: https://scialert.net/abstract/?doi=pjbs.2014.472.481
DOI: 10.3923/pjbs.2014.472.481
URL: https://scialert.net/abstract/?doi=pjbs.2014.472.481
INTRODUCTION
Rice is the most important cereal crop and also rice genetic transformation is a foremost object for cereal biotechnology. Transgenic research and technology have been progressing steadily and the introduction of multiple genes into rice cultivars via transformation has become more important for desired trait improvement. However, transformations of multiple traits into rice varieties usually require the use of selectable marker systems for the selection of putative transformants. The efficiency of stable gene transfer is not high even in the most successful transfer systems (Datta et al., 1998) and only a fraction of the cells is exposed to integrate the DNA construction into their genomes. As well successful genetic transformation method does not promise expression level by the regulation of transgene expression.
The introduction of most foreign genes into plant does not present a phenotype and it can be used to identify the transformed cells. Encoding product of a selectable marker gene allows surviving the transformed cell and it restrict the growth of non-transformed cell. The growth of non-transformed cells was suppressed by the lowest concentration of toxic compound and does not effect to transformed ones (Roa-Rodriguez et al., 2003). Among the most widely used antibiotic resistance genes as the selectable markers are hygromycin phosphotransferase (hptII) (Brodersen et al., 2000), neomycin phosphotransferase (nptII) (Schroeder et al., 1993), gentamycin acetyltransferase resistance (McBride and Summerfelt, 1990), bleomycin (Hille et al., 1986) and phleomycin resistance (Mulsant et al., 1988).
Encoding plasmid hygromycin resistance gene in Escherichia coli was isolated and sequenced by Gritz and Davies (1983). Hygromycin resistance is an amino glycoside antibiotic produced by Streptomyces hygroscopicus and suitable marker system for both plant and animal. It is 1026 base pairs long, 39000 mol wt and encoding for hygromycin phosphotransferase (Zheng et al., 1991). The hptII gene from E. coli confers resistance to hygromycin and the resistance gene codes for a kinase that inactivates hygromycin through phosphorylation and more toxic than kanamycin. It kills sensitive cells more rapidly and has been used for plant transformation in different species (Akutsu et al., 2004). At the present time, it is one of the preferred antibiotic resistance marker systems for monocot plants transformation, mostly gramineae. The explant type, the developmental stage, the tissue culture conditions and the genotype were important for sensitivity of selective agent (Eustice and Wilhelm, 1984). The successful hygromycin selection system can be used in developing an efficient system of embryogenic suspension cultures for sweet potato genotypes (Liu et al., 2001). Also, Janna et al. (2000) reported that hygromycin had been found to be an effective selective agent used in Dendrobium plant transformation. It was capable to cause complete fatality of the tissues at very low concentrations, i.e., 10, 20 and 25 mg L-1. Thus, hygromycin resistance may be an interesting and useful marker system in rice transformation for different purposes. Keeping in view all these factors, the sensitivity level of the potential target tissue to hygromycin and determination of the minimum inhibitory concentration of the selective agents was vital prior to developing an efficient transformation protocol. Therefore the suitability of hygromycin resistance studied as a selectable marker system in selected Malaysian rice. The hptII gene coding for hygromycin resistance is an effective and reliable alternative selectable marker for rice transformation. The marker system developed here is a progressive step towards multiple genetic transformations of rice and supply an important alternative selection.
MATERIALS AND METHODS
Hygromycin sensitivity study: The callus was induced as the target tissue for genetic transformation and was investigated for its hygromycin sensitivity. Two genotypes MR219 line 4 and 9 were two potential rice mutants that were generated from MR219 and used in this study. Hygromycin was prepared at 50 mg L-1 in a phosphate buffer saline solution. In the preliminary study, the experiment was conducted with a large range of different concentrations, i.e., 0, 20, 40, 60, 80 and 100 mg L-1. Hygromycin was added onto the callus induction medium containing MS+10 μM 2,4-D. The data were recorded at one-week interval. Notably, changes in the appearance of callus from creamish/yellowish to brownish would be described as the indicator for the death of callus tissues. The growth of the untreated control callus was considered as 100%, assuming that there was no inhibitory of growth effect. Therefore, the percentage of the treated tissues was related to the untreated callus tissues. In order to determine the minimal inhibitory concentration of hygromycin, repeated experiments of different concentrations were conducted with lower and specific concentrations, i.e., 0, 5, 10, 15, 20, 25, 30, 35, 40, 45 and 50 mg L-1. The growth rate of the untreated tissues was used as the standard and calculated following the formula below:
For the transformation study, the mature embryos were isolated and cultured as described above. The plasmid pCAMBIA 1304 (PCAMBIA plasmid carrying a mgfp 5 and gus A by CaMV 35S promoter CAMBIA-mgfp-nos and hptII gene) was used for this study (Fig. 1). The rice callus clumps were placed in the centre of a moist filter paper in sterile petri plates (approximately 100-150 mg of somatic embryos per plate) and partly dried in a laminar flow hood for 15 min prior to bombardment. Plasmid DNA was precipitated into gold particles and bombarded according to the protocols supplied for the Biolistic PDS-1000/He particle delivery system (BioRad, USA) with minor modifications. DNA was coated on the gold particles and bombarded at 1100 psi helium gas pressure under 27 in Hg vacuum at a shooting distance of 6 cm from the rupture disk to the target tissue.
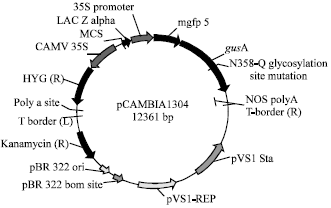 | |
| Fig. 1: | A Schematic diagram of the plasmid pCAMBIA 1304.The binary vector pCAMBIA 1304 (CSIRO, Australia) harboring the reporter gusA and mgfp5 genes driven by the CaMV 35S promoter |
Immediately after bombardment, the embryogenic callus were cultured on a medium without any selection agent for three days. Then, the explants were cultured on an MS medium containing hygromycin according to the above result and sub-cultured on the same medium.
Histochemical gus staining and fluorescence microscopy: The gus assay was carried out according to the method described by Jefferson (1989). Concerning gus, the bombarded and non-bombarded tissues were placed in 1.5 mL individual Eppendorf tubes and stained overnight at 37°C with 0.5 mL of filter sterilized gus assay buffer {100 mM Na3PO4 (pH 7.0), 10 mM EDTA, 1 mM K3Fe(CN)6, 1 mM K4Fe(CN)6, 2 mM 5-bromo-4-chloro-3-indolyl--D-GlcUA (X-Gluc) (50 mg mL-1) and 0.1% Triton X-100}. The stained tissues were then transformed into 95% ethanol for 24 h to remove chlorophylls. No transformed rice callus was used as control. The transient gus activity was recorded as blue spots using a light microscope.
Selection and regeneration of transformants: In the above result, the minimal hygromycin concentration, it was suggested to be able to eliminate the non-transformed cells and used to select the putative transformed plant. After transformation with plasmid PCAMBIA 1304, the bombarded calli were transferred onto the hygromycin containing medium to select the putative transformants. After two weeks of post bombardment, the calli were transferred to the selection medium and the data were recorded. The putative transformants were monitored weekly and the surviving calli were used for further molecular analysis. These putative transformants were maintained and sub-cultured on a fresh medium for continuous selection and molecular analysis in order to confirm the presence of transgene for further study.
Total plant RNA extraction and reverse transcription-polymerase chain reaction (RT-PCR): The total RNA used for RT-PCR analysis was extracted from the stable transformed plants. Sixteen independently derived transgenic plants, confirmed to contain the hptII and gus, were analyzed by RT-PCR to characterize the expression at the RNA level. The total plant of RNA was extracted using the RNeasy mini kit according to the manufacturer’s instructions (QIAGEN). One-step RT-PCR was carried out using a thermal cycler (TECHNE- TC 521) and it was preheated to 50°C with a heated lid before placing the samples. The first strand of the cDNA synthesis by reverse transcription was performed in one cycle at 50°C for 30 min and followed by initial PCR activation at 95°C for 15 min. Then, the PCR amplification was undertaken for 35 cycles by initial denaturation at 94°C for 1 min, annealing at 55°C for 1 min and elongation at 72°C for 1 min 30 sec. A final elongation step was done at 72°C for 10 min for the gus genes. For hptII one cycle at 50°C for 30 min, initial PCR activation at 95°C for 15 min and followed by the PCR amplification was undertaken for 30 cycles by denaturation at 94°C for 1 min, annealing at 55°C for 1 min and elongation at 72°C for 1 min. A final elongation step was done at 72°C for 10 min and holding at 4°C. The primers used for hptII amplification were as follows: 5’ATG CGG AGC ATA TAC GCC 3’ for the forward primer and 5’ATG AAA AAG CCT GAA CTC AC3’ for the reverse primer and 5’ATG CTC TAC ACC ACG CCG AAC AC3’(Forward) and 5’TCA AGA AGG ACC ATG TGG T3’(Reverse) for gus. gene. Southern blot analysis was performed according to a standard protocol. A non radioactive method of southern blotting hybridization analysis was performed to identify the presence of transgene in the genomic of the transformed tissue. DIG DNA labelling and Detection kit (Roche) was used for this study. The kit uses dig oxigenin (DIG), a steroid hapten, to label DNA probes for hybridization and subsequent colour detection by enzyme immunoassay.
Statistical analysis: All experiments were conducted in three replicates. The data were analysed in a completely randomized design using the SAS and MSTATC computer programmes. In addition, the comparisons of means were tested for significance using the LSD test at 0.05 level of probability.
RESULTS
The mature embryos were isolated and cultured on the best callus induction medium, i.e. MS+10 μM 2,4-D containing different concentrations of hygromycin. As a preliminary study, higher amounts (i.e., 0, 20, 40, 60, 80 and 100 mg L-1) were used in two genotypes, i.e., MR219 line 4 and 9. The callus tissues were found completely dead at low concentrations of 40 and 60 mg L-1 (Table 1, Fig. 2).
| Table 1: | The number and percentage of the surviving rice callus in the medium combination with different concentrations of hygromycin |
 | |
| All these data were resulted from the three replicates | |
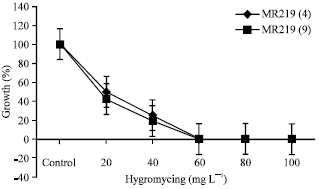 | |
| Fig. 2: | Minimum inhibitory level of hygromycin concentration in the two rice genotypes, i.e., MR219 line 4 and MR219 line 9 |
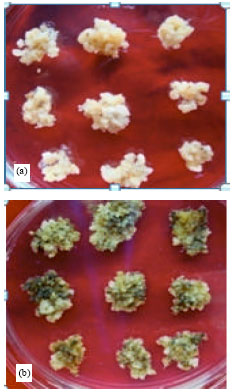
The toxicity effect of hygromycin was clearly found on the reduction of growth at 80 and 100% for the callus tissues treated with 40 mg L-1. It was clearly evident that the physical appearance of the calli cultured on the medium containing hygromycin was distinctly different from the calli cultured on the control medium in all concentration (Fig. 3). As early as the first week of incubation period, the toxicity effect was observed in the callus tissues and it was rapid. Based on these findings, low concentrations of hygromycin were sufficient to kill the untransformed calli and these callus were highly significantly sensitive to hygromycin. They were brown or whitish in color indicating the death of the tissues in contrast to the healthy and actively proliferating calli.
 | |
| Fig. 3(a-f): | The morphology of calli on different concentrations of hygromycin as the selectable agent and the calli cultured on the media containing (a) 0 mg L-1, (b) 20 mg L-1, (c) 40 mg L-1, (d) 60 mg L-1, (e) 80 mg L-1 and (f) 100 mg L-1 of hygromycin concentrations. The bar scale shows 1 cm |
Also, Table 2 shows that, in the analysis of variance, the effects of hygromycin concentration (0, 20, 40, 60, 80, 100) and all genotypes, the interaction between hygromycin concentration and genotypes were significantly different (p<0.001).
Concerning the study of incubation period during the first week of callus formation, the efficiency on the medium without hygromycin was 100% while the callus formation efficiency varied from 91.13, 70.16, 36.88, 25.21 and 11.61% in MR219 line 4 and 83.34, 50.32, 27.76, 23.58 and 10.91% in MR219 line 9 on the media containing 20, 40, 60, 80 and 100 mg L-1 hygromycin, respectively. The callus formation rate was constant on the hygromycin free media during the 2nd, 3rd, 4th and 5th weeks. The values dropped to 68.72, 43.75, 30.78, 19.05 and 4.89% in MR219 line 4 and 52.45, 37.46, 25.21, 21.98 and 9.16 % in MR219 line 9 for 20, 40, 60, 80 and 100 mg L-1 hygromycin, respectively during the 2nd week. This trend continued in the 3rd, 4th and 5th weeks. In addition, the callus survival rates after five weeks were 24.27, 9.8, 0, 0 and 0% for MR219 line 4 and 35.48, 10.61, 0, 0 and 0% for MR219 line 9 on 20, 40, 60, 80 and 100 mg L hygromycin, respectively (Table 3). These results showed that hygromycin might be an effective selective agent for identification of transformants in rice.
Additionally, determination of the minimum specific inhibitory level was also essential while lower concentrations would allow the chance of getting the untransformed cells and higher concentrations would inhibit the transformed cells. Table 4 shows the number and percentage of the surviving rice calli in the medium combination with different concentrations of hygromycin (i.e., 0, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55 and 60 mg L-1), the callus growth weight was totally inhibited at 45 mg L-1 (Fig. 4).
| Table 2: | Analysis of variance of the effects of the different concentrations of hygromycin in the media and genotypes on the callus growth rate |
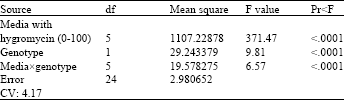 | |
| The result showed that the media and genotypes were significantly different (p<0.001) for all genotypes and media. Also, the analysis revealed a significant effect on the genotypes and media interaction (p<0.001) | |
Clearly, it was evident that the morphological appearance of the calli cultured on the medium added with hygromycin was distinctly different from the tissue culture on the control medium. These calli were brownish in colour, indicating the death of callus tissues which was in contrast to the healthy embryogenic calli. Likewise, a similar observation was discovered by Tee et al. (2001). The toxicity effect of 30 mg L-1 hygromycin concentration on the calli was observed as soon as the first week of incubation period. Notably, the surviving callus percentage decreased in the following weeks. It was recommended that there was a rapid inhibitory effect on the callus tissues tested even at low concentrations. Also, according to the analysis of variance as shown in Table 5, the effects of different concentrations of hygromycin media and all genotypes, as well as the interaction between media and genotypes were significantly different (p<0.001). Thus, based on these results, 45 mg L-1 hygromycin seemed suitable for the identification of transformants. In a separate experiment, the calli bombarded with the hptII gene were placed on the selection media. Seemingly, these calli were bright in colour, larger in size and grew faster as compared to the control calli growing on the media containing hygromycin (Fig. 5).
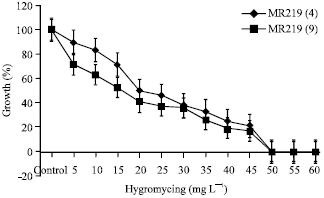 | |
| Fig. 4: | The minimum inhibitory level of hygromycin concentration in the two selected rice genotypes, i.e., MR219 line 4 and MR219 line 9. All the data were resulted from the three replicates |
| Table 3: | Callus induction response of MR219 line 4 and MR219 line 9 on the media containing different concentrations of hygromycin at weekly interval |
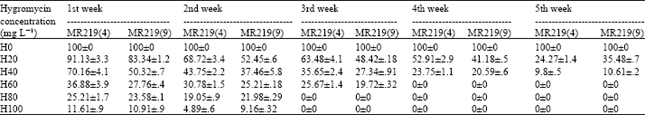 | |
| The value followed by ±shows the mean standard deviation | |
The plants regenerated from these calli proved positive in the molecular analysis which confirmed the effectiveness of the selection system.
The total RNA was extracted from randomly selected transient hptII expressing plant and non-transformed plants. RT-PCR was performed using the specific primers (1317 bp) gene fragment was amplified for all selected plants which were resistant to hygromycin selection. Approximately 20 calli per bombardment and 18 plates were used in the experiment. Plants regenerated on hygromycin selection showed high levels of gus reporter gene expression via., histochemical assay (Fig. 6), resistant and gus-positive transformants were analyzed by RT-PCR using gus gene primers and hptII gene primers.
| Table 4: | Number and percentage of the surviving rice callus in the medium combination with different concentrations of hygromycin |
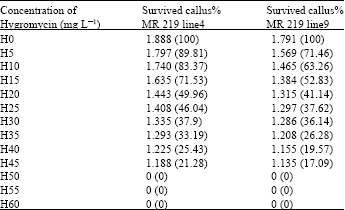 | |
| All these data were resulted from the three replicates | |
 | |
| Fig. 5(a-b): | Bombarded calli were transferred to hygromycin containing the selection medium of (a) Untransformed callus and (b) Transformed callus |
 | |
| Fig. 6(a-d): | Gus expression using histochemical analysis in untransformed callus (a) Transformed callus, (b) Shoot regenerated with gus gene expression, (c) Untransformed shoot and (d) Transformed shoot |
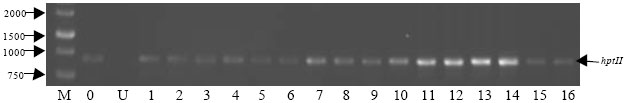 | |
| Fig. 7: | Representative RT-PCR analysis for hptII (815 bp) transcription in transgenic rice plants, lane M: Size markers (1-kbp DNA ladder), Lane C: Positive control from Access RT-PCR system, Lane U: Untransformed plant (negative control) and Lane 1-16: Transgenic plant lines |
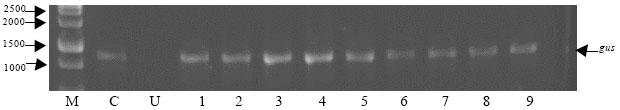 | |
| Fig. 8: | Representative RT-PCR analysis for gus (1317 bp) transcription in transgenic rice plants, Lane M: Size markers (1-kbp DNA ladder), Lane C: Positive control from Access RT-PCR system, Lane U: Untransformed plant (negative control) and Lane 1-9: Transgenic plant lines |
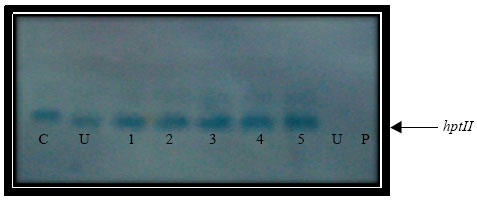 | |
| Fig. 9: | Southern analysis of hptII gene (815 bp) from transgenic rice plants, Lane P: Control plasmid, Lane C: PCR amplified product, Lane U: Untransform plant and Lane 1-6: Selected transgenic plants |
| Table 5: | Analysis of variance of the effects of the different concentrations of hygromycin media and genotypes on the callus growth rate |
 | |
| Results showed that the media, genotypes and the media interaction were significantly different (p<0.001) in all genotypes and media, respectively | |
Predicted 815 bp for hptII (Fig. 7) and 1317 bp (Fig. 8) for gus gene were both amplified from all regenerated plants, indicating the presence of both genes in recovered plants. Apparently, RNA from the negative control (non-transformed calli) did not show any band. This confirmed the expression of hptII and gus genes in the transiently transformed lines. Meanwhile, RT-PCR from the known mRNA provided along with the kit was used as the positive control. Six plants were randomly selected for Southern blot analysis. DNA samples from all the plants showed hybridization with the DIG labeled probe from the coding sequence of the hptII gene, showing that the hygromycin gene has been inserted into the rice genome (Fig. 9). One or more hybridization signals were observed in each of the gus and PCR positive lines. No hybridization signals were detected in non-transformed control plants (Fig. 10).
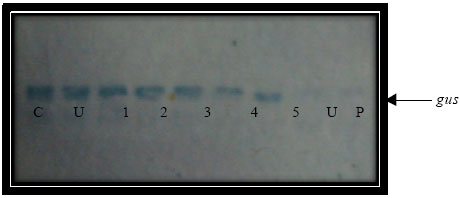 | |
| Fig. 10: | Southern analysis of gus gene (1317 bp) from transgenic rice plants, Lane P: Control plasmid, Lane C: PCR amplified product, Lane U: untransform plant, lane 1-6 selected transgenic plants |
| Table 6: | The transformation efficiency by using the optimized transformation protocol. Transformation efficiency was calculated as the number of surviving tissues on the hygromycin selection over the number of bombarded tissues of each experiment |
 | |
| Transformation efficiency: No. of surviving tissues on the hygromycin selection/No. of bombarded tissuesx100% | |
The difference in the patterns of hybridization signals indicated the independent transformation events and the random integration of the transgene. Also in this study recovered 16 independent transgenic rice lines and determined the overall transformation efficiency by calculating the number of independently derived plants regenerated on the hygromycin-containing medium as a proportion of the total number of calli subjected to particle bombardment. A transformation frequency of 2.5% was achieved for the hptII gene (Table 6).
DISCUSSION
Cellular selection is often necessary in tissue culture and molecular biology. Most systems use a specific dominant selectable marker to enable the recovery of transgenic tissues. Hygromycin is the standard selection agent for rice somatic embryo tissues. However, possibilities exist in using alternative selection agents and markers in place of hygromycin (Rao et al., 2009). A potential group of herbicides, amino acids and their analogs had been tested as possible selection agents to further improve transgenic rice. In this study successfully transformed Mutant MR 219 with exogenous genes using a biolistic procedure. The present transformation and selection system could be applicable to other cultivars. In the previous reports hygromycin at 50 mg L-1 completely prevented untransformed callus growth and effective selective agent for rice transformation also for Dendrobium (Yu et al., 1999; Men et al., 2003). Compared to kanamycin, the use of hygromycin as the selective agent required a much lower concentration that could effectively eliminate the non-transformants. In addition, hygromycin was a superior selective agent for oncidium orchid tissue (Janna et al., 2000). Recently, there is evidence of successful recovery of transgenic orchid tissue using hygromycin as the selectable marker and selective agent. At the same time, the transformants of Oncidium were selected at 5 mg L-1 and this showed high susceptibility of this orchid to hygromycin (Liau et al., 2003). Likewise, this result was also reported for monocots. A successful application of hygromycin as the selective agent to obtain the transgenic crop plant was reported for legume (Aoki et al., 2002) oat (Cho et al., 1999) oil palm callus (Parveez et al., 1996), also ornamentals such as petunia (Shaw et al., 2002), Liliaceous ornamental plant (Suzuki et al., 2001) and Lavender (Mishiba et al., 2001).
Hygromycin-resistant and positive transformants were analyzed by PCR using hptII gene primers. Predicted 815 bp for hptII were amplified from all regenerated plants, indicating the presence of the hpt gene in recovered plants. Sixteen plants were randomly selected for RT PCR analysis. Transformation efficiencies with PCAMBIA 1304 with hygromycin selection were 2.5% in MR219 line 4. Hygromycin selection was very effective and reliable and no escape was observed and Hygromycin mediated selection was sensitive and effective. This indicates that hygromycin mediated transformation may have a wide range of usage and the system is flexible with respect to different needs in rice also in pulm (Tian et al., 2009). The results are important with respect to gene expression technology development. Although various selectable markers have been described in the literature (Miki and McHugh, 2004; Tian et al., 2006), no sole marker can get all purposes and can be simply used across all the species for genetic transformation. A marker which showed effective in selection in one species may not be effective in another species. Evaluation of a selectable marker for its suitability and the establishment of a selection system with the new marker is important and necessary for a plant species. Our study demonstrates that hygromycin is an effective and reliable selectable marker for rice genetic transformation.
In summary, hygromycin caused significant effects on the survival, growth, size and colour of callus which was directly proportional to the concentration of hygromycin. Although, a number of calli survived on low concentrations of hygromycin, the values were significantly different from the control treatment at all concentrations. These effects can even multiply when these genotypes are continuously screened for long periods. Hence, hygromycin may cause damage to the transgenic callus and can produce abnormal genotypes (Nakazawa and Matsui, 2003). In this study the hygromycin concentration of 45 mg L-1 was capable of completely causing for the screening of putative transformants. This was different from the recommended concentration (25 mg L-1) of hygromycin B for the selection of transgenic rice (Datta et al., 1990). In addition, the different genotypes have different levels of hygromycin sensitivity. Therefore, the determination of hygromycin toxicity on plant crops is necessary before applying hygromycin as the selective antibiotic for the regeneration of transgenic plants. Finally, rice callus can be stably transformed and the transgenic characters can be maintained. This result offers a possibility for the application of plant genetic engineering in rice breeding. The hygromycin selection system developed here is useful for gene stacking and provides an important alternative selection system for rice.
CONCLUSION
Results strongly suggest that hygromycin could successfully be used in experiments involving transformation of mutant line MR219 and identification of putative transformants. It is also clear that an efficient and reproducible selection system involves continuous selection for approximately five weeks. According all of result it could be concluded that the hygromycin concentration 45mg L-1 was capable of completely causing the fatality to both of rice callus MR219 line 4 and 9, respectively. This specific hygromycin concentration could be applied for screening of putative transformants when using this callus as the target tissue for plant transformation.
ACKNOWLEDGMENT
The authors are grateful to Universiti Putra Malaysia (UPM) and TWOWS (Third World Organization for Women in Science) for their financial support. Our heartful thanks also go to the Department of Biochemistry, the Faculty of Biotechnology and Biomolecular Science for granting us the access to the laboratory facilities.
REFERENCES
- Aoki, T., A. Kamizawa and S. Ayabe, 2002. Efficient Agrobacterium-mediated transformation of Lotus japonicus with reliable antibiotic selection. Plant Cell Rep., 21: 238-243.
CrossRef - Brodersen, D.E., W.M. Jr, Clemons, A.P. Carter, R.J. Morgan-Warren, B.T. Wimberly and V. Ramakrishnan, 2000. The structural basis for the action of the antibiotics tetracycline, pactamycin and hygromycin on the 30S ribosomal subunit. Cell, 103: 1143-1154.
PubMedDirect Link - Cho, M.J., W. Jiang and P.G. Lemaux, 1999. High-frequency transformation of oat via microprojectile bombardment of seed derived highly regenerative cultures. Plant Sci., 148: 9-17.
CrossRef - Datta, K., A. Vasquez, J. Tu, L. Torrizo and M.F. Alam et al., 1998. Constitutive and tissue-specific differential expression of the cryIA(b) gene in transgenic rice plants conferring resistance to rice insect pests. Theor. Applied Gene, 97: 20-30.
Direct Link - Eustice, D.C. and J.M. Wilhelm, 1984. Mechanisms of action of aminoglycoside antibiotics in eucaryotic protein synthesis. Antimicrob. Agents Chemother, 26: 53-60.
Direct Link - Gritz, L. and J. Davies, 1983. Plasmid-encoded hygromycin B resistance: The sequence of hygromycin B phosphotransferase gene and its expression in Escherichia coli and Saccharomyces cerevisiae. Gene, 25: 179-188.
PubMedDirect Link - Hille, J., F. Verheggen, P. Roelvink, H. Franssen, A. Van Kammen and P. Zabel, 1986. Bleomycin resistance: A new dominant selectable marker for plant cell transformation. Plant Mol. Biol., 7: 171-176.
CrossRefDirect Link - Janna, O.A., M. Maziah and G.K.A. Parvezz, 2000. Research note: Potential selective agent fororchid transformation. Asia Pacific J. Mol. Biol. Biotech., 8: 85-93.
Direct Link - Liau, C.H., S.J. You, V. Prasad, H.H. Hsiao, J.C. Lu, N.S. Yang and M.T. Chan, 2003. Agrobacterium tumefaciens-mediated transformation of an Oncidium orchid. Plant Cell Rep., 21: 993-998.
PubMedDirect Link - Liu, Q.C., H. Zhai, Y. Wang and D.P. Zhang, 2001. Efficient plant regeneration from embryogenic suspension cultures of sweet potato. In vitro Cell Develop. Bio. Plant, 37: 564-567.
CrossRef - McBride, K.E. and K.R. Summerfelt, 1990. Improved binary vectors for Agrobacterium-mediated plant transformation. Plant Mol. Biol., 14: 269-276.
CrossRefDirect Link - Men, S., X. Ming, Y. Wang, R. Liu, C. Wei and Y. Li, 2003. Genetic transformation of two species of orchid by biolistic bombardment. Plant Cell Rep., 21: 592-598.
CrossRefPubMedDirect Link - Miki, B. and S. McHugh, 2004. Selectable marker genes in transgenic plants: applications, alternatives and biosafety. J. Biotechnol., 107: 193-232.
CrossRef - Mishiba, K.I., K. Ishikawai, O. Tsujii and M. MII, 2001. Efficient transformation of lavender (Lavandula latifolia medicus) mediated by Agrobacterium. J. Hortic. Sci. Biotech., 75: 287-292.
Direct Link - Mulsant, P., A. Gatignol, M. Dalens and G. Tiraby, 1988. Phleomycin resistance as a dominant selectable marker in CHO cells. Somatic Cell Mol. Genet., 14: 243-252.
CrossRefDirect Link - Parveez, G.K.A., M.K.U. Chowdhury and N.M. Saleh, 1996. Determination of minimal inhibitory concentration of selection agents for oil palm Elaeis guineensis Jacq. transformation. Asia-Pac. J. Mol. Biol. Biotechnol., 4: 219-228.
Direct Link - Rao, S.S., L. Mamadou, M. McConnell, R. Polisetty, P. Kwanyuen and D. Hildebrand, 2009. Non-antibiotic selection systems for soybean somatic embryos: The lysine analog aminoethyl-cysteine as a selection agent. BMC Biotechnol., Vol. 9.
CrossRefDirect Link - Schroeder, H.E., A.H. Schotz, T. Wardley-Richardson, D. Spencer and T.J.V. Higgins, 1993. Transformation and regeneration of two cultivars of pea (Pisum sativum L.). Plant Physiol., 101: 751-757.
Direct Link - Shaw, J.F., H.H. Chen, M.F. Tsai, C.I. Kuo and L.C. Huang, 2002. Extended flower longevity of petunia hybrida plants transformed with boers, a mutated ERS gene of Brassica oleracea. Mol. Breed., 9: 211-261.
CrossRef - Suzuki, S., K. Supaibulwatana, M. Mii and M. Nakano, 2001. Production of transgenic plants of the Liliaceous ornamental plant Agapanthus praecox ssp. orientalis (Leighton) Leighton via Agrobacterium-mediated transformation of embryogenic calli. Plant Sci., 161: 89-97.
Direct Link - Tian, L., F.A. Canli, X. Wang and S. Sibbald, 2009. Genetic transformation of Prunus domestica L. using the hpt gene coding for hygromycin resistance as the selectable marker. L. Sci. Hortic., 119: 339-343.
CrossRef - Yu, Z., M. Chen, L. Nie, H. Lu and X. Ming et al., 1999. Recovery of transgenic orchid plants with hygromycin selection by particle bombardment to protocorms. Plant Cell Tissue Organ Cult., 58: 87-92.
Direct Link - Zheng, Z., A. Hayashimoto, L. Zhijian and N. Murai, 1991. Hygromycin resistance genecassettes for vector construction and selection of transformed rice protoplasts. Plant. Physiol., 97: 832-835.
CrossRefDirect Link