
Saliu Joseph Kayode
University of Lagos, Nigeria
Ugorji Ijeoma Chidimma
University of Lagos, Nigeria
Ekpendu Emeka Alwell
University of Lagos, Nigeria
Pakistan Journal of Biological Sciences
Year: 2014 | Volume: 17 | Issue: 12 | Page No.: 1225-1230







ABSTRACT
The adverse effect of crude oil on aquatic ecosystem is a modern day peril. This study investigated the acute toxicities of some Nigerian crude oil (Forcados, Bonnylight and Qua-Iboe) against Clarias gariepinus and the antioxidant enzyme response of the fish to the pollutants. The toxicity assessment was based on LC50 at 96 h. Static renewal bioassays were used to determine the toxicological effects of the 3 crude oils on the antioxidant enzymes of post juveniles Clarias gariepinus using the liver. The 96 h LC50 value for Qua Iboe was 18.966 mL L-1, Forcados, 2.776 mL L-1 and Bonny light, 3.948 mL L-1, indicating that Forcados is the most toxic of the crude oils. Induction of the Glutathione-S-transferase (GST), Glutathione (GSH), Superoxide dismutase (SOD) and Catalase (CAT) in the liver of Clarias gariepinus exposed to Forcados and Bonny light for a period of 28 days was inhibited while it was enhanced in Qua-Iboe crude oil. Malondialdehyde (MDA) levels increased with increasing toxicity and exposure time. SOD, CAT, GSH, GST and MDA are reliable environmental biomarkers for crude oil induced oxidative stress in Clarias gariepinus and therefore useful biological indicators of environmental contamination in the aquatic ecosystem.
PDF Abstract XML References Citation
Received: June 05, 2014;
Accepted: September 24, 2014;
Published: November 11, 2014
How to cite this article
Saliu Joseph Kayode, Ugorji Ijeoma Chidimma and Ekpendu Emeka Alwell, 2014. Response of Some Antioxidant Parameters in Post Juveniles of Clarias
gariepinus after Exposure to Nigerian Crude Oil (Forcados, Bonny Light and Qua-Iboe). Pakistan Journal of Biological Sciences, 17: 1225-1230.
DOI: 10.3923/pjbs.2014.1225.1230
URL: https://scialert.net/abstract/?doi=pjbs.2014.1225.1230
DOI: 10.3923/pjbs.2014.1225.1230
URL: https://scialert.net/abstract/?doi=pjbs.2014.1225.1230
INTRODUCTION
The aquatic ecosystem serves as the final destination for most contaminants (Winston and Di Giulio, 1991). The study and diagnosis of aquatic contamination is therefore of paramount importance.
Crude oil pollutant is among the most important and abundant class of pollutants found in the aquatic environment. The dissolution of the soluble components of crude oil in the surrounding water body causes direct problems for fish (due to its contact with their liver and gills) organs responsible for gaseous exchange (Pacheco and Santos, 2001). This soluble components contain most of the carcinogenic Polycyclic Aromatic Hydrocarbons (PAHs) and have been related to neoplasic and genotoxic effects found in the liver of Benthic fish, (Baumann and Harshbarger, 1998) as well as to toxic effects, such as oxidative stress (Achuba and Osakwe, 2003; Van der Oost et al., 2003; Zhang et al., 2003).
In the last three decades, the major revenue source for Nigeria has been the oil industry. Exploratory activity and exploitation of oil occurs mainly in the southern part of Nigeria, with the Niger delta area accounting for over 90% of crude oil production (Wegwu and Akininwor, 2006). Repeated oil spillage in Nigeria, has led to deleterious effects on both the terrestrial and aquatic ecosystems. According to Wegwu and Akaninwor (2006) the oil producing communities of the Niger delta have become synonymous with oil spills which exert severe disastrous impact on the marine and soil ecosystems. In coastal oil producing areas, the marine and soil ecosystems serve as the main source of livelihood to inhabitants and are determinants of agricultural productivity (Osuji and Adesiyan, 2005). Peculiar to these oil producing communities are the numerous network of crude oil pipelines criss crossing the land mass. One of such network of pipelines, belonging to a major oil producing company is estimated to cover an area of 31000 km2, with 79 flow stations and two export terminals (NDES., 1999). Several factors have been implicated for the spillages that occur repeatedly in these areas which include, hose failure, bursts arising from rupture of flow lines, corrosion of flow lines, sabotage and vandalization of oil installations and production equipments (Wegwu and Akaninwor, 2006).
The local fisheries of these communities have also been affected as the fishes constantly migrate to avoid the oil logged areas (Powell, 1987). Ezenwa and Ayinla (1994) reported that the quota of fish contributed from the Niger delta to the national output was drastically reduced. This has also subsequently led to a reduction in the artisanal and pelagic sectors of the local fishery.
Clarias gariepinus is a model fish largely used in the field of ecotoxicology. Several biochemical responses have been well characterized in this fish in response to exposure to toxic compounds (Rodriguez-Fuentes and Gold-Bouchot, 2004).
It is one of the most common commercially grown fresh water fish and is of great importance locally, serving as a delicacy among many tribes.
This study aims to determine the level of oxidative enzyme response in post juveniles of Clarias gariepinus exposed to sublethal concentrations of 3 Nigerian crude oils (Forcados, Bonny light and Qua-iboe).
MATERIALS AND METHODS
Test animals, source and acclimatization: Clarias gariepinus post juveniles (6-8 weeks old) were purchased from a local fish farm in Surulere, Lagos and transported in open gallons, half filled with pond water to holding tanks (length: 45.00 cm, height 34.00 cm, bottom diameter: 25.00 cm and top diameter: 35.00 cm) in the laboratory. The post juveniles were kept in the plastic holding tanks containing dechlorinated water to acclimatize to laboratory condition (28±2°C) for a period of seven days before they were used in the bioassays. The post juveniles were fed with fish food (Coppens-2 mm) at 3% of body weight twice daily and the water changed once every 48 h aerating it continuously with Bzagdon air pump (double type 1200).
Test compounds: Three crude oils (Forcados, Bonny light and Qua-iboe) used in this study were obtained from Exxon Mobil and Shell Nigeria through the Department of Petroleum Resources (DPR).
General bioassay techniques
Bioassay containers: Rectangular plastic tanks (5 L capacity, with dimensions 22.00x15.00x18.00 cm) were used as bioassay container
Preparation of test media: To prepare test media for bioassays, computed volumes of stock solutions were made up to 4 L. Preliminary studies showed that 5 post juveniles survived well in 4 L of test media for 7 days without aeration.
Selection of animals for bioassay: Active post juveniles of similar age and size (age: 6-8 week old, mean snout to tail length: 16.00-23.00 cm, mean weight 33.00-47.00 g) were taken from holding tanks and randomly assigned to experimental tanks.
Biochemical studies of Clarias gariepinus exposed to sub lethal concentrations of forcados, bonny light and Qua-iboe crude oil acting singly in semi static bioassays: Seven post-juveniles of similar age and size were exposed to sub-lethal concentration (1/10th of the LC50 value) and untreated control in 2 replicates. These series of bioassays went on for 28 days and the semi static bioassay procedure was adopted in order to avoid drastic changes in concentration of test media via evaporation and excessive reduction in dissolved oxygen level. In the semi static procedure, each test media was changed into a fresh solution of exactly the same concentration of crude oil or untreated control, respectively once every three days, transferring the same exposed test animals into the freshly prepared test media over the 28 day period of the experiment.
At time intervals of 7, 14 and 28 days, one live C. gariepinus per replicate (making two per treatment and two from untreated control) were randomly selected, dissected and the liver was kept refrigerated pending further analysis. Clarias gariepinus was exposed to sub lethal concentrations of crude oil in separate experiment as follows:
| • | Forcados at 0.3 mL L-1 (0.1 of 96 h LC50) |
| • | Bonny light at 0.4 mL L-1 (0.1 of 96 h LC50) |
| • | Qua-Iboe at 2 mL L-1 (0.1 of 96 h LC50) |
Biochemical studies of liver of Clarias gariepinus exposed to sub lethal concentrations of Forcados, Bonny light and Qua-Iboe crude oil: The following substrate and enzyme activity was analyzed in the liver of Clarias gariepinus exposed to sub lethal concentrations of Forcados, Bonny light and Qua-Iboe.
Superoxide dismutase (SOD) activity: Superoxide dismutase activity was determined by its ability to inhibit the auto-oxidation of epinephrine which was estimated by the increase in absorbance at 480 nm as described by Sun and Zigman (1978). The reaction mixture (3 mL) contained 2.95 mL 0.05 M sodium carbonate buffer pH 10.2, 0.02 mL of liver homogenate and 0.03 mL of epinephrine in 0.005 N HCl was used to initiate the reaction. The reference cuvette contained 2.95 mL buffer, 0.03 mL of substrate (epinephrine) and 0.02 mL of water. Enzyme activity was calculated by measuring the change in absorbance at 480 nm for 5 min:
Catalase activity: Serum catalase activity was determined by measuring the decrease in absorbance at 240 nm due to the decomposition of H2O2 in a UV recording spectrophotometer. The reaction mixture (3 mL) contained 0.1 mL of serum in phosphate buffer (50 mM, pH 7.0) and 2.9 mL of 30 mM H2O2 in phosphate buffer pH 7.0. An extinction coefficient for H2O2 at 240 nm of 40.0 M-1 cm-1 (Aebi, 1984) was used for the calculation. The specific activity of catalase was expressed as moles of H2O2 reduced per minute per milligram protein.
Glutathione-s-transferase activity: Glutathione-S-transferase activity was determined by the method of Habig et al. (1974).
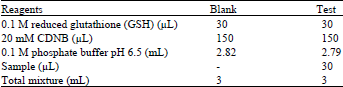
Principle: The assay is based on the fact that all known glutathione-S-transferase demonstrate a relatively high activity with 1-Chloro-2, 4-dintrobenzene (CDNB) as the second substrate. Consequently, the conventional assay for glutathione-S-transferase activity utilizes 1-Chloro-2, 4-dintrobenzene as substrate. When this substrate is conjugated with reduced glutathione (GSH), its absorption maximum shifts to a longer wavelength. The absorption increase at the new wavelength of 340 nm provides a direct measurement of the enzymatic reaction.
Reagents
20 mM 1-Chloro-2,4-dinitrobenzene (CDNB): The 3.37 mg of CDNB (Sigma Chemicals Co, London) was dissolved in 1 mL of ethanol.
Reduced glutathione (0.1 M): The 30.73 mg (0.0307 g) of reduced glutathione powder (Sigma Chemical Co, London) was dissolved in 1 mL of 0.1 M phosphate buffer (pH 6).
Procedure: The medium for the estimation of GST activity was prepared as shown in Table 1 and the reaction was allowed to run for 60 sec each time before the absorbance was read against the blank at 340 nm. The temperature was maintained at approximately 31°C. The absorbance was measured using spectrophotometer.
Calculation:
| SOD/CAT/GST | = | ΔA/minxVT/ΣxVs |
| MDA/GSH | = | AxVT / ΣxVs |
| ΔA | = | Change in absorbance |
| VT | = | Total volume |
| Vs | = | Sample volume |
| Σ | = | Molar extinction |
Glutathione (GSH) determination: The reduced glutathione (GSH) content of liver tissue as non-protein sulphydryls was estimated according to the method described by Sedlak and Lindsay (1968). To the tissue homogenate, 10% TCA was added, centrifuged. Then 1.0 mL of supernatant was treated with 0.5 mL of Ellmans reagent (19.8 mg of 5, 5-dithiobisnitro benzoic acid (DTNB) in 100 mL of 0.1% sodium nitrate) and 3.0 mL of phosphate buffer (0.2 M, pH 8.0). The absorbance was read at 412 nm.
Lipid peroxidation: Malondialdehyde (MDA) an index of lipid peroxidation was determined using the method of Buege and Aust (1978). Briefly 1.0 mL of the supernatant was added to 2 mL of (1:1:1 ratio) TCA-TBA-HCl reagent (thiobarbituric acid 0.37%, 0.24N HCl and 15% TCA) tricarboxylic acid and thiobarbituric acid-hydrochloric acid reagent boiled at 100°C for 15 min and allowed to cool. Flocculent materials were removed by centrifuging at 3000 rpm for 10 min. The supernatant was removed and the absorbance read at 532 nm against a blank. MDA was calculated using the molar extinction coefficient for MDATBA-complex of 1.56x105 M-1 cm-1.
Physico- chemical parameters of Forcados, Bonny light and Qua-Iboe crude oil: The physico-chemical parameters for Forcados, Bonny light and Qua-Iboe crude oil and the fresh water used in analysis are given in Table 2 and 3.
Statistical analysis: The dose response data was analyzed by probit analysis after Finney (1971) using SPSS 10.00 (statistical package).
| Table 1: | Preparation of the medium for the estimation of GST |
 | |
| Table 2: | Physico-chemical parameters of Forcados, Bonnylight and Qua-Iboe crude oil |
 | |
| Table 3: | Physico-chemical parameters of fresh water used in this experiment |
 | |
One way analysis of variance (ANOVA) was used to compare the means of results obtained from biochemical analysis.
RESULTS
LC50 values at 96 h: The 96 h LC50 values of the three crude oil showed that Forcados is the most toxic, followed by Bonny light and Qua-Iboe (2.776, 3.948 and 18.966 mL L-1, respectively).
Biochemical studies of liver of Clarias gariepinus exposed to sub lethal concentrations of Forcados, Bonny light and Qua-Iboe crude oil: The lipid peroxidation assay indicated that the level of malondialdehyde (MDA) in the liver of fish exposed to all the test chemicals (Forcados, Bonny light and Qua-Iboe crude oil) increased significantly (p<0.05), when compared to control animals (Table 4). The level of MDA increased from a range of 0.0040 nM g-1 in the control group to 1.533 nM g-1 in the exposed groups. The highest level of peroxidation was in fish exposed to Forcados, followed by Bonny light and the least in Qua-Iboe.
Conversely, the activity of the enzymes SOD, CAT, GST and GSH were inhibited in fishes exposed to Forcados and Bonny light crude oil but was enhanced when exposed to the Qua Iboe crude oil (Table 4). Measurements of SOD, CAT, GST and GSH activities in liver of fish exposed to sub lethal concentration of the test chemicals (Forcados and Bonny light crude oil) over a 28 day period decreased significantly (p<0.05) when compared to control animals but increased significantly (p<0.05) when exposed to the Qua iboe oil.
The level of SOD decreased from a range of 1.193 U mg-1 in control groups of Forcados and Bonny light to 0.436 U mg-1 (Forcados) and 0.6667 U mg-1 (Bonny light) in the exposed groups.
However, the SOD activity increased in Qua-Iboe from 1.193 U mg-1 in control group to 5.573 U mg-1 in exposed group (p<0.05) (Table 4).
The level of CAT decreased from 7.760 U mg-1 in the control groups of Forcados and Bonny light to 2.937 U mg-1 (Forcados) and 0.6667 U mg-1 (Bonny light) in the exposed groups (Table 4). CAT, however, increased in the control group of Qua-iboe from 7.760-35.99 U mg-1. The levels of GST activities decreased from 1.123 U mg-1 in Forcados and Bonny light control groups to 0.5967 U mg-1 (Forcados) and 1.097 U mg-1 (Bonny light). In Qua iboe, however, the level of GST activities increased from 1.123 U mg-1 in control group to 10.36 U mg-1 in exposed group (Table 4).
| Table 4: | Activities of MDA, SOD, CAT, GST and GSH from post-juvenile of Clarias gariepinus exposed singly to Forcados, Bonny light and Qua Iboe crude oil |
 | |
| S.D: Standard deviation, *Significant different (p<0.05), **More significant (p<0.01), ***Most significant (p<0.001) | |
The GSH decreased from 0.1533 U mg-1 in control group to 0.04667 U mg-1 (Forcados) and 0.08667 U mg-1 (Bonny light). However the level of GSH activity increased from 0.1533 U mg-1 in control groups to 0.5967 U mg-1 (Table 4).
DISCUSSION
This study revealed differential toxicities in the three crude oils tested against C. gariepinus (based on LC50 values). Otitoloju (2006) observed that differential toxicity between test chemicals can be tied to the differences in the nature of the physical and chemical constituents. Thus the action site and toxicity exerted on target organisms. The greater toxicity of Forcados when compared to the other two (Bonny light and Qua-iboe) can be attributed to a higher composition of low boiling point hydrocarbons, such as aromatics, including benzene, toluene, xylene naphthalene and phenanthrene, all of which generally have a higher solubility in water and are known to be toxic to a wide variety of marine organisms. The bonny light crude oil also had a higher composition of these hydrocarbons than Qua-iboe thus conferring on it a higher toxicity.
The liver is an organ that performs various functions associated with the metabolism of xenobiotics (Jiminez and Stegeman, 1990). The breakdown of aliphatic and aromatic hydrocarbons and other xenobiotics lead to the generation of free radical species in various tissues (Achuba and Osakwe, 2003). Hepatocytes rely on antioxidant enzymes to protect them from the activity of reactive oxygen species which are products of transformation of xenobiotics. Antioxidant enzyme activity can be enhanced or inhibited by chemical stress and this depends on the intensity and duration of stress applied and also susceptibility of the target species.
In this study there was an inhibition of GST, GSH, SOD and CAT in Forcados and Bonny light crude oils while there was an enhancement of these anti oxidant enzymes in Qua iboe. Previous workers have reported the inhibition of antioxidant enzymes in crude oil and refined petroleum oil. Wang et al. (2006) reported the inhibition of GST activity in Sebasticus marmoratus after exposure to Benzene. Otitoloju and Olagoke (2011) noted a significant reduction (p<0.05), in SOD and GST in the liver of Clarias gariepinus exposed to crude oil, benzene, toluene and xylene while Vieira et al. (2008) reported a significant decrease in acetylcholinestrase (ACLE) and GST in Pomatoschistos microps exposed to high concentrations of Anthracene.
Inhibition in anti oxidant enzyme activity is responsible for the significant lipid peroxidation observed in this study. Malondialdehyde (MDA) which is one of the oxidative damage products of lipid peroxidation was found to increase in the 3 crude oils in the study. Such increases have been reported by Farombi et al. (2007).
Oxidative stress due to the toxic effect of pollutants is usually indicated by increased levels of products of oxidative damage (MDA) and subsequent decrease in defense enzymes (GST-GSH, SOD, CAT) due to overwhelming effects of the pollutants (Otitoloju and Olagoke, 2011).
The enhancement of the antioxidants in Qua-Iboe crude which is an indication of lower toxicity and oxidative stress as shown in results observed in this study may be attributed to the fact that Qua-Iboe is the most viscous among the three oils, hence the least soluble in water and less available for uptake. Farombi et al. (2007) and Saliu and Bawa-Allah (2012) similarly reported elevated levels of some antioxidant enzymes in Clarias gareipinus exposed to heavy metals.
Exposure of aquatic organisms to crude and refined oils inflicts severe impact on various aspects of fish physiology sometimes leading to large scale mortalities. This study is therefore significant in indicating that the response of antioxidant enzymes can be used as potential environmental biomarkers for crude oil induced oxidative stress in fish.
REFERENCES
- Achuba, F.I. and S.A. Osakwe, 2003. Petroleum-induced free radical toxicity in African catfish (Clarias gariepinus). Fish Physiol. Biochem., Vol. 29.
CrossRefDirect Link - Baumann, P.C. and J.C. Harshbarger, 1998. Long Term Trends in Liver Neoplasm Epizootics of Brown Bullhead in the Black River, Ohio. In: Trends in Levels and Effects of Persistent Toxic Substances in the Great Lakes, Gilbertson, M., G.A. Fox and W.W. Bowerman (Eds.). Springer, New York, USA., ISBN-13: 9789401062275, pp: 213-223.
- Buege, J.A. and S.D. Aust, 1978. Microsomal Lipid, Peroxidation. In: Methods in Enzymology, Vol. 52, Flesicher, S. and L. Packer (Eds.)., Academic Press, New York, pp: 302-310.
CrossRefDirect Link - Ezenwa, B.I. and O.A. Ayinla, 1994. Conservation strategies for endangered fish breeding and nursery grounds within the coastal wetlands of Nigeria. Aquat. Conserv.: Mar. Fresh Water Ecosyst., 4: 125-133.
CrossRefDirect Link - Farombi, E.O., O.A. Adelowo and Y.R. Ajimoko, 2007. Biomarkers of oxidative stress and heavy metal levels as indicators of environmental pollution in African cat fish (Clarias gariepinus) from Nigeria Ogun River. Int. J. Environ. Res. Public Health, 4: 158-165.
CrossRefPubMedDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Jiminez, B.D. and J.J. Stegeman, 1990. Detoxification enzymes as indicator of environmental stress on fishes. Am. Fish Soc. Symp., 8: 69-79.
Direct Link - Osuji, L.C. and S.O. Adesiyan, 2005. The Isiokpo oil-pipeline leakage: Total organic carbon/organic matter contents of affected soils. Chem. Biodivers., 2: 1079-1085.
CrossRefPubMedDirect Link - Otitoloju, A.A., 2006. Joint action toxicity of spent lubrication oil and laundry detergent against Poecilia reticulata (Teleostei: Poeciliidae). Afr. J. Aquat. Sci., 31: 125-129.
CrossRefDirect Link - Otitoloju, A. and O. Olagoke, 2011. Lipid peroxidation and antioxidant defense enzymes in Clarias gariepinus as useful biomarkers for monitoring exposure to polycyclic aromatic hydrocarbons. Environ. Monit. Assess., 182: 205-213.
CrossRefDirect Link - Pacheco, M. and M.A. Santos, 2001. Biotransformation, endocrine and genetic responses of Anguilla anguilla L. to petroleum distillate products and environmentally contaminated waters. Ecotoxicol. Environ. Saf., 49: 64-75.
CrossRefDirect Link - Rodriguez-Fuentes, G. and G. Gold-Bouchot, 2004. Characterization of cholinesterase activity from different tissues of Nile tilapia (Oreochromis niloticus). Mar. Environ. Res., 58: 505-509.
CrossRefDirect Link - Saliu, J.K. and K.A. Bawa-Allah, 2012. Toxicological effects of lead and zinc on the antioxidant enzyme activities of post juvenile Clarias gariepinus. Resour. Environ., 2: 21-26.
CrossRef - Sedlak, J. and R.H. Lindsay, 1968. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman's reagent. Anal. Biochem., 25: 192-205.
CrossRefPubMedDirect Link - Sun, M. and S. Zigman, 1978. An improved spectrophotometric assay for superoxide dismutase based on epinephrine autoxidation. Anal. Biochem., 90: 81-89.
CrossRefPubMedDirect Link - van der Oost, R., J. Beyer and N.P.E. Vermeulen, 2003. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol., 13: 57-149.
CrossRefPubMedDirect Link - Vieira, L.R., A. Sousa, M.F. Frasco, I. Lirna, F. Morgado and L. Guilhermino, 2008. Acute effects of Benzo[a]pyrene, anthracene and a fuel oil on biomarkers of the common goby Pomatoschistus microps (Teleostei, Gobiidae). Sci. Total Environ., 395: 87-100.
Direct Link - Wang, C., Y. Zhao, R. Zheng, X. Ding, W. Wei, Z. Zuo and Y. Chen, 2006. Effects of tributyltin, benzo[a]pyrene and their mixture on antioxidant defense systems in Sebastiscus marmoratus. Ecotoxicol. Environ. Saf., 65: 381-387.
CrossRefPubMedDirect Link - Wegwu, M.O. and J.O. Akiniwor, 2006. Assessment of heavy-metal profile of the new Calabar river and its impact on Juvenile Clarias gariepinus. Chem. Biodivers., 3: 79-87.
CrossRefDirect Link - Winston, G.W. and R.T. Di Giulio, 1991. Prooxidant and antioxidant mechanisms in aquatic organisms. Aquat. Toxicol., 19: 137-161.
CrossRef - Zhang, J.F., H. Shen, T.L. Xu, X.R. Wang, W.M. Li and Y.F. Gu, 2003. Effects of long-term exposure of low-level diesel oil on the antioxidant defense system of fish. Bull. Environ. Contam. Toxicol., 71: 234-239.
CrossRefDirect Link