
Yasser A. Ahmed
Faculty of Veterinary Medicine, South Valley University, Qena 83523, Egypt
Soha A. Soliman
Faculty of Veterinary Medicine, South Valley University, Qena 83523, Egypt
Enas A. Abdel-Hafez
Faculty of Veterinary Medicine, Assiut University, Assiut, Egypt
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 17 | Page No.: 859-864







ABSTRACT
The current study aimed to describe the histological changes of the femur and tibia of the post-hatching quail. Femur and tibia from 1-day- to 6-weeks post-hatching quail were processed for light microscopy. Histological examination revealed that endochondral ossification was a delayed process in the development of femur and tibia preceded by periosteal ossification. Femur and tibia of 1-day-post-hatching quail consisted of growth cartilage enclosed in a tube of periosteal bone collar. The collar extended toward the epiphysis dividing it into articular cartilage proper and lateral articular cartilage. Down to the articular cartilage, there was a physeal growth cartilage, in which the chondrocytes were organized into resting, proliferative and hypertrophic zones. Focal areas of hypertrophic chondrocytes were observed in the epiphysis of the tibia but not of the femur, which acted as a nidus for formation of the secondary ossification centre after in 2-week-posthathcing quail. Primary ossification centre was seen in both femur and tibia after 2 weeks and ossification continued replacing the cartilage until the 6th week when only permanent articular cartilage remained. Cartilage canals were present in both femur and tibia starting from the day 1, but chondrified and completely disappeared after the 6th week. The current study suggests that the periosteal ossification preceded the endochondral ossification and plays an important role in quail long bones development.
PDF Abstract XML References Citation
Received: January 11, 2013;
Accepted: March 04, 2013;
Published: April 15, 2013
How to cite this article
Yasser A. Ahmed, Soha A. Soliman and Enas A. Abdel-Hafez, 2013. Ossification of the Femur and Tibia of the Post-Hatching Japanese Quail. Pakistan Journal of Biological Sciences, 16: 859-864.
DOI: 10.3923/pjbs.2013.859.864
URL: https://scialert.net/abstract/?doi=pjbs.2013.859.864
DOI: 10.3923/pjbs.2013.859.864
URL: https://scialert.net/abstract/?doi=pjbs.2013.859.864
INTRODUCTION
Long bone growth and development occurs through the process of endochondral ossification in which a cartilage template is formed and gradually replaced by bone tissue. During the embryonic life, condensed undifferentiated mesenchymal cells differentiate into chondrocytes which secrete a specific collagen type-II and proteoglycan- rich matrix of a cartilage template. The central chondrocytes of the cartilage template undergo further differentiation into hypertrophic cells and a periosteal bone collar is formed around them. The hypertrophic chondrocytes physiologically die and the cartilage matrix at this area is invaded by bone cells, blood cells and blood vessels forming a Primary Ossification Centre (POC) at the middle of the cartilage template. Similarly, a Secondary Ossification Centre (SOC) is established at each end of the template. Postnatally, remnants of the cartilage template remain only as Physeal Growth Cartilage (PGC) between the POC and SOC and around the SOC as articular cartilage. The ossification spreads through the cartilage and at specific time when the animal matures, the POC and SOC get together leaving only the permanent articular cartilage (reviewed by Mackie et al., 2008, 2011; Cicek et al., 2011). The chondrocytes of the PGC are organized into three distinct zones; resting, proliferative and hypertrophic. The resting chondrocytes are rounded and randomly distributed throughout the cartilage matrix, the proliferative chondrocytes are flattened and arranged in columns parallel to the longitudinal axis of the long bone, while the hypertrophic chondrocytes are enlarged cells important for calcifying the cartilage matrix (Jeffcott and Henson, 1998; Rivas and Shapiro, 2002). Hypertrophic chondrocytes die by modes of physiological non-apoptotic death before replacement of the cartilage remnants with bone tissue (Roach and Clarke, 2000; Ahmed et al., 2007; Chen et al., 2010). Although, the permanent articular cartilage is avascular organ, the developing growth cartilage are normally invaded by many vascular channels contributing to nourishment of the developing long cartilage and thought to help in the formation of the SOC (Burkus et al., 1993; Blumer et al., 2005) but as the animal matures, cartilage canals of the growth cartilage normally chondrify and disappear. The long bone development of mammals has extensively been studied, however, information on the development of the avian long bones is very poor (Pourlis et al., 2006; Simsa and Ornan, 2007). The current study highlighted the process of long bone ossification in quail.
MATERIALS AND METHODS
Raising of birds: Japanese quail (Coturnix japonica) were raised in the quail research centre belonging to the Histology Department, Faculty of Veterinary Medicine, South Valley University and used for the current experiment. The birds after hatching were feed on poultry starter 28% protein with soybean meal sorghum was added to the ration as an external source of protein. After 14 days, the ration was poultry starter with 21% protein until the end of the experiment. Temperature was 38°C during the first week, but decreased to 24-30°C to the end of the experiment. Regarding the light program, the birds after hatching till the first 3 days were exposed to 24 h light day-1 and decreased gradually to reach 12 h day-1 after 2 weeks to the end of the experiment.
Sample processing for light microscopy: The birds were euthanized using ether inhalation and then the femur and tibia of at least 5 male birds during each stage of post-hatching development (1 day, 1 week, 2 weeks, 3 weeks, 4 weeks and 6 weeks) were carefully dissected without touching the articular surfaces with the scalpel and washed thoroughly in Phosphate-buffered Saline (PBS). Specimens were fixed in either 10% formalin (pH; 7.4) at 4°C for 3 days, then they were decalcified in 0.33 M Ethylene-diamine-tetra-acetic Acid (EDTA). The whole femur and tibia as a whole (before formation of the POC) or the growth cartilage with some adjacent bone (when the POC was found) from the proximal ends of femur and tibia were dissected, dehydrated in ascending grades of ethanol and finally embedded in paraffin using standard methods. Paraffin sections (3-5 μm thickness) were undertaken and stained with hematoxylin and eosin, safranin-O, Crossman’s trichrome and Periodic Acid-Schiff’s reagent (PAS) and examined with light microscopy.
RESULTS
At day one, the femur and tibia were completely formed of cartilage enclosed by a tube of periosteal bone collar. The bone collar was enclosed into the epiphysis dividing articular growth cartilage above the Physeal Growth Cartilage (PGC) into articular cartilage proper and lateral articular cartilage (Fig. 1a). The periosteal bone collar was separated from the PGC by a mixture of haematopoietic cells concentrated at the centre of the femur and tibia, penetrated through the peripherally of the cartilage and run parallel to the periosteal bone collar (Fig. 1a,b). Zonation of PGC into resting, proliferative and hypertrophic areas was present but not clear (Fig. 1c). The chondrocytes of the articular cartilage was overcrowded and randomly distributed throughout the cartilage extracellular matrix. Cellular condensation and focal hypertrophy of chondrocytes were seen in the epiphysis of tibia only but not femur of 1-day- and 1-week-post hatching quail and was also penetrated with cartilage canals (Fig. 1d). The SOC was seen for the first time in the epiphysis of the tibia (Fig. 1e). It was noticed that the proliferative zone of the femur and tibia noticeably longer than of mammals and the proliferative zone of the tibia was much longer than that of the femur. During this stage of development, large areas of the PGC were degraded and replaced by bone trabeculae of the POC (Fig. 1e-f) and a bone marrow cavity filled with a mixture of haematopoietic and fat cells (Fig. 1f). Thus the tibia from 2-week-post-hatching quail had 2 centres of ossification; POC and SOC and the cartilage is divided into PGC between the POC and the SOC and the articular cartilage between the articular surface and the SOC (Fig. 1e). While, the femur at the same stage of development had only POC (Fig. 1f) and the cartilage is still divided into articular cartilage above PGC. In older birds, the ossification gradually replaced the cartilage (Fig. 1g-h).
The chondrocytes of the PGC were organized into clear three zones, resting, proliferative and hypertrophic after 2 weeks of hatching (Fig. 2a-b) and the zonal organization of the chondrocytes became well defined as the development proceeded. The cartilage canals of 1-day-old quail contained blood capillaries that were surrounded by many undifferentiated mesenchymal cells (Fig. 2c). The cartilage canals of 2-week and older post-hatching quail had well defined wall and contained arterioles surrounded by many mesenchymal cells (Fig. 2d). After four weeks, blood vessels of many of the cartilage canals diapered and the contents of the canals were mostly fibrinoid and chondrified (Fig. 2e). By the 6th week, the cartilage canals were no longer observed within the permanent articular cartilage (Fig. 2f).
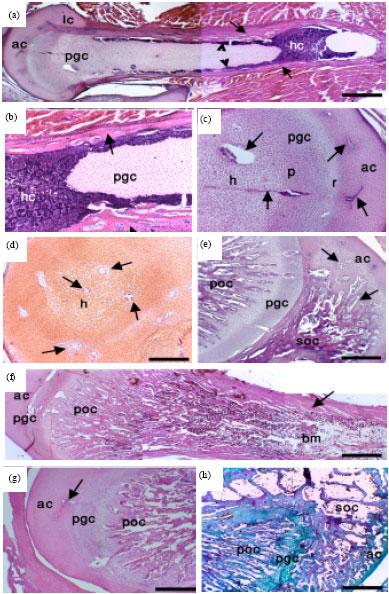 | |
| Fig. 1(a-h): | Periosteal and endochondral ossification of the femur and tibia of the post-hatching Japanese quail. Paraffin sections from the femur (a, b, c, f, g) and tibia (d, e, h) from 1-day (a, b, c), 1-week (d), 2-week (e, f) and 4-week (g-h) -post-hatching quail stained with H and E (a, b, c, f, g), safranin-O (d) and PAS (e). Note articular cartilage proper (ac), lateral articular cartilage (lc), physeal growth cartilage (pgc), haematopoietic cells (hc), resting (r), proliferative (p), hypertrophic (h) chondrocytes, primary ossification centre (poc), secondary ossification centre (soc), physeal growth cartilage (pgc) and bone marrow (bm). Arrows indicate periosteal bone collar (a, b, f) and cartilage canals (a, c, d, e, g) and arrowheads indicate haematopoietic cells (a). Bars = 200 μm in a, f, g, h, 80 μm in b, c, d, e and 140 μm in F |
DISCUSSION
The aim of the current study was to illustrate the main histological changes of the quail long bones during post-hatching development. To achieve this aim, paraffin sections of the femur and tibia from 1 day to 6 weeks was examined with light microscopy. The femur and tibia of 1-day-post-hatching quail was formed of cartilage enclosed by a tube of periosteal bone collar. The bony tube extended from the metaphysis into the epiphysis dividing the articular cartilage into inner and outer parts. This picture is similar to the larval stages of frogs (Mackie et al., 2008).
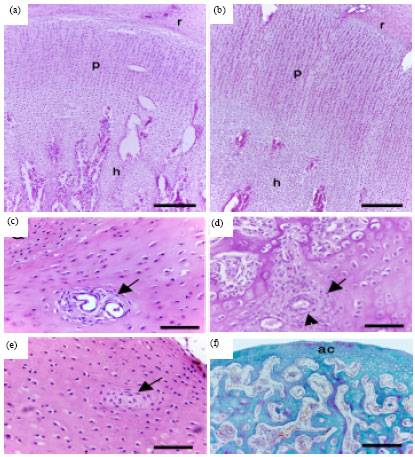 | |
| Fig. 2(a-d): | Histology of the growth cartilage of the post-hatching Japanese quail, Paraffin sections from the 2-week (a, b), 1-day (c), 3-week (d), 4 week (e) and 6-week (f)- post-hatching quail stained with H&E (a, b, e), PAS (c, d) and Crossman’s trichrome (f), Note, resting (r), proliferative (p), hypertrophic (h) zones and articular cartilage (ac). Arrows indicate cartilage canals and arrowhead indicates arterioles within the canals. Bars = 80 μm in a, b and 20 μm in c, d, e, f |
The periosteal bone collar was reported to firstly observed around the long bone at 6-day of the pre-hatching quail and is formed of bone rings surrounding the central hypertrophic zone of the developing cartilaginous long bone, then increases in thickness and expands radially toward the epiphysis with development of the future bone (Pourlis et al., 2006). No sign of endochondral ossification was seen in the developing femur or tibia of the quail before the 2nd week of the post-hatching development, when bone trabeculae was firstly seen forming the POC within the metaphysis of the bone. Thus the development of the periosteal bone in such way is likely to limit the diameter expansion of the developing bone especially until the first 2-post-hatching weeks. Unlike mammals (Rivas and Shapiro, 2002), endochondral ossification in the femur and tibia of quail may be a delayed process and long bone growth and development in the quail may depend more on the periosteal ossification than endochondral ossification at least during the early stages of post-hatching development. Although no endochondral ossification occurred within the PGC, the chondrocytes were undergoing proliferation and hypertrophy as mammals (Miralles-Flores and Delgado-Baeza, 1990; Rivas and Shapiro, 2002). This supports the idea that chondrocyte proliferation and hypertrophy may not related to the matrix calcification, however, it is important for radial expansion associated with bone elongation through periosteal ossification (Felisbino and Carvalho, 1999). A mixture of haematopoietic cells was observed at the middle of the femur and tibia, which extended on both sides of the cartilage under the periosteal bone collar. These cells are likely are important for cartilage erosion and formation of marrow cavity when the development advanced. A striking feature of the bone development in quail was the observation of foci of hypertrophic chondrocytes in the epiphysis of the tibia but not the femur at 1-day and 1-week-post-hatching and later on the SOC was seen in the tibia but not femur at the 2nd week of post-hatching development. These hypertrophic chondrocytes acts as a nucleus for the formation of the SOC and that is similar to what is happening in mammals (Rivas and Shapiro, 2002). It was noticeable that the proliferative zone of the femur and tibia was usually long in comparison to mammals. One of the main histological differences between mammalian and avian long bone PGC is that the avian cartilage has longer proliferative column and larger blood vessels that penetrate further into the hypertrophic zone (Leach and Gay, 1987). Further more, the proliferative zone of the tibia was longer than that of the femur. Taken this with our result that the tibia is longer than the femur and the high weight gain of the quail after the second week of post-hatching development (not published), that may indicate the presence of the SOC in the tibia during this time.
Although, it is known that the cartilage is avascular tissue, the growth cartilage was invaded with a vascular cartilage canals. We described the development of the cartilage canals during embryonic development in another paper (submitted for publication). In this study we found the cartilage canals were found within the cartilage starting from one-day-post-hatching and within the SOC. Cartilage canals provide not only the vasculature and nutrition to the chondrocytes but also the osteoblast precursors, the undifferentiated mesenchymal cells that may contribute to formation of the SOC (Burkus et al., 1993; Rivas and Shapiro, 2002; Blumer et al., 2004; 2005; 2006). The cartilage canals were completely disappeared from the cartilage of mature bird after 6 weeks of post-natal development. The cartilage canals undergo a normal regression, chondrification (Haines, 1974) and retention of the cartilage canals may lead to dyschondroplasia in many species (Ytrehus et al., 2007).
CONCLUSION
In conclusion, the current study showed that, endochondral ossification is a late process in the post-hatching quail, however the bone elongation occur mostly due to the periosteal ossification. The femur, unlike the tibia, has no SOC and the cartilage canals normally disappeared at maturity.
REFERENCES
- Ahmed, Y.A., L. Tatarczuch, C.N. Pagel, H.M. Davies, M. Mirams and E.J. Mackie, 2007. Physiological death of hypertrophic chondrocytes. Osteoarthritis Cart., 15: 575-586.
CrossRefPubMedDirect Link - Blumer, M.J.F., H. Fritsch, K. Pfaller and E. Brenner, 2004. Cartilage canals in the chicken embryo: Ultrastructure and function. Anat. Embryol., 207: 453-462.
CrossRefDirect Link - Blumer, M.J.F., S. Longato, E. Richter, M.T. Perez, K.Z. Konakci and H. Fritsch, 2005. The role of cartilage canals in endochondral and perichondral bone formation: Are there similarities between these two processes? J. Anat., 206: 359-372.
CrossRefDirect Link - Blumer, M.J.F., C. Schwarzer, M.T. Perez, K.Z. Konakci and H. Fritsch, 2006. Identification and location of bone-forming cells within cartilage canals on their course into the secondary ossification centre. J. Anat., 208: 695-707.
CrossRefDirect Link - Burkus, J.K., T.M. Ganey and J.A. Ogden, 1993. Development of the cartilage canals and the secondary center of ossification in the distal chondroepiphysis of the prenatal human femur. Yale J. Biol. Med., 66: 193-202.
Direct Link - Chen, K.S., L. Tatarczuch, M. Mirams, Y.A. Ahmed, N.C. Pagel and E.J. Mackie, 2010. Periostin expression distinguishes between light and dark hypertrophic chondrocytes. Int. J. Biochem. Cell. Biol., 42: 880-889.
CrossRefPubMedDirect Link - Cicek, K., M. Kumas and D. Ayas, 2011. Differentiation of bone tissue and long bone development in the Uludag frog, Rana macrocnemis tadpoles. Biharean Biologist, 5: 123-126.
Direct Link - Mackie, E.J., L. Tatarczuch and M. Mirams, 2011. The skeleton: A multi-functional complex organ. The growth plate chondrocyte and endochondral ossification. J. Endocrinol., 211: 109-121.
CrossRef - Felisbino, S.L. and H.F. Carvalho, 1999. The epiphyseal cartilage and growth of long bones in Rana catesbeiana. Tissue Cell., 31: 301-307.
CrossRef - Haines, R.W., 1974. The pseudoepiphysis of the first metacarpal of man. J. Anat., 117: 145-158.
Direct Link - Mackie, E.J., Y.A. Ahmed, L. Tatarczuch, K.S. Chen and M. Mirams, 2008. Endochondral ossification: How cartilage is converted into bone in the developing skeleton. Int. J. Biochem. Cell. Biol., 40: 46-62.
CrossRefPubMedDirect Link - Miralles-Flores, C.M. and E.D. Delgado-Baeza, 1990. Histomorphometric differences between the lateral region and central region of the growth plate in fifteen-day-old rats. Acta Anat., 139: 209-213.
CrossRef - Pourlis, A.F., J. Antonopoulos and I.N. Magras, 2006. A light and electron microscopic study of the limb long bones perichondral ossification in the quail embryo (Coturnix coturnix Japonica). Ital. J. Anat. Embryol., 111: 159-170.
PubMed - Rivas, R. and F. Shapiro, 2002. Structural stages in the development of the long bones and epiphyses: A study in the New Zealand white rabbit. J. Bone Joint Surg. Am., 84-A: 85-100.
PubMedDirect Link - Roach, H.I. and N.M. Clarke, 2000. Physiological cell death of chondrocytes in vivo is not confined to apoptosis: New observations on the mammalian growth plate. J. Bone Joint Surg. Br., 82: 601-613.
PubMed - Simsa, S. and E.M. Ornan, 2007. Endochondral ossification process of the turkey (Meleagris gallopavo) during embryonic and juvenile development. Poult. Sci., 86: 565-571.
CrossRefDirect Link