
H. Aliyu
Department of Biological Sciences, Ahmadu Bello University, Zaria, Nigeria
M.F. Ishiyaku
Department of Plant Sciences, Ahmadu Bello University, Zaria, Nigeria
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 15 | Page No.: 743-746







ABSTRACT
The development of cowpea aphid larvae was monitored on seven cowpea genotypes (IAR-48, TVu-15866, IT84S-2246-4, SAKA BABBA SATA, IT90K-76, KANANNADO and TVX 3236). The aim of the study was to determine the developmental response of the larvae as an indication of antibiotic resistance of the genotypes. Highly significant differences (p<0.01) were observed with respect to fertility, larval development, adult longevity, life span, multiplication rate and intrinsic rate of increase. KANANNADO and TVX 3236 show minimum antibiotic effects while a landrace SAKA BABBA SATA shows relatively high antibiotic effects. This result further reveals the potential of SAKA BABBA SATA as a resistance source to aphid. The reaction of IT84S-2246-4, a hitherto aphid resistant genotype, which supported higher levels of survival of the larvae relative to other known susceptible genotype IAR-48, may be an indication of the presence of a new biotype of Aphis craccivora endemic to Zaria environs, or that of the ability of insects to overcome hindrances to their survival including various forms of resistance.
PDF Abstract XML References Citation
Received: December 04, 2012;
Accepted: February 15, 2013;
Published: April 04, 2013
How to cite this article
H. Aliyu and M.F. Ishiyaku, 2013. Identification of Novel Resistance Gene Sources to Cowpea Aphid (Aphis craccivora Koch) in Cowpea (Vigna unguiculata L.). Pakistan Journal of Biological Sciences, 16: 743-746.
DOI: 10.3923/pjbs.2013.743.746
URL: https://scialert.net/abstract/?doi=pjbs.2013.743.746
DOI: 10.3923/pjbs.2013.743.746
URL: https://scialert.net/abstract/?doi=pjbs.2013.743.746
INTRODUCTION
Cowpea (Vigna unguiculata L. Walp. Subsp. Unguiculata Fabaceae) is one of the most important food and forage legumes in the tropics. Cowpea is grown on 10.5 million ha, with an annual grain production of about 5.5 million tonnes (http://faostat.fao.org). It is mainly cultivated for the seeds, however other important products from it include the pods (fresh or dried) and leaves (Duke, 1990). By far Nigeria is the biggest producer of cowpea, followed by Niger republic, Burkina Faso and Ghana. Cowpea is however devastated by many biotic and abiotic stresses. Prominent among the biotic constraints are various types of insect pest and one of the key insect pest of cowpea is the cowpea aphid, especially at vegetative stage (also infest at reproductive stage). It primarily infests seedlings and causes damage directly by removal of sap from the plant and indirectly through transmission of Aphid Borne Mosaic Viruses (ABMV). Aphids also cause damage through secretion of honeydew which promotes growth of sooty moulds and other fungal growth on leaves, thereby reducing photosynthetic efficiency of the plant (Annan et al., 1996). Resistance to insect pests in cowpea has been extensively studied. Most cowpea accessions have been screened for resistance to the major insect pest. High to moderate level of resistance has been reported for some of the pests, including cowpea aphids (Singh et al., 1997) and many different levels of these resistances have incorporated into several genotype. Similarly, the genetics of aphid resistance has been studied in some of the resistant cultivars. Two independent and non-allelic genes control the expression of resistance (Pathak, 1988, Myers et al., 1996). However these studies were limited to few of the already identified resistance sources. Also data from genetic crosses showed that more than locus may be involved in the expression of the resistance (Githiri et al., 1996). Therefore, further studies are needed to cover more resistant sources. IT84S-2246 derived its resistance to cowpea aphids from Tvu 1190 (Singh and Singh, 1992). According to B. B. Singh, TVU 1190 and TVu 3000 have genes for aphid’s resistance that do not segregate for aphid reaction (Myers et al., 1996). However, in the routine cowpea program in cowpea section of the IAR, it was observed that IT84S-2246 and lines derived from it show susceptible reaction to aphids. Therefore, it is also necessary to investigate these lines to confirm the presence of resistance or otherwise. This study reports the possibility of the presence of variants of aphids (in Zaria) that are able to attack known resistant lines.
MATERIALS AND METHODS
Genotypes and study area: Seven genotypes (SAKA BABBASATA, TVU 15866, IT90K-76, IT84-2246, IAR-48, TVX 3236 and KANANNADO) were obtained from cowpea section of Institute for Agricultural Research, Ahmadu Bello University and Zaria (11010’ 00’’N 07038’ 00’’E). These genotypes were identified based on the preliminary evaluation were SAKA BABBASATA (a local line), TVU 15866 and IT90K-76 turn out to indicate some level of resistance while IT84S-2246 shows an unprecedented level of aphid support. IAR-48 and TVX 3236 are improved aphids susceptible lines and KANANNADO is a susceptible local line.
Experimental design: Each genotype was sown in pots filled with soil in insect-proof cages in the screen house. There were three replications in a Randomized Complete Design (RCD).
A single aphid was placed on each plant for each genotype. After the release of two larvae the mother aphid along with any other larvae were removed. The newly hatched larvae were maintained to produce the second-generation larvae. A single aphid of this generation was subsequently observed daily to determine the duration of larval stage, the duration of reproductive life and the total number of larvae produced (fertility). These were used to calculate Multiplication Rate (MR) and the intrinsic rate of natural increase (rm), using the following formulas:
where:
(Asin and Pons, 2001), where, Fe = effective fertility = number of larvae produced during a period equivalent to Tpr and Tpr = pre-reproductive time = number of days from the birth of the aphid until production of the first larva. The data obtained was analyzed to compare the genotypes in terms of larval development. In either case the data analysis was carried out using the SAS 9.0 software.
RESULTS
The analysis of variance result for the reaction of aphids is presented in Table 1. The result of the analysis showed highly significant differences (p<0.01) for fertility, larval development, adult longevity, intrinsic rate of increase and multiplication rate of aphids in the seven genotypes studied.
Aphid fertility was highest on KANANNADO with 62.0 larvae and lowest on IT90K-76 with 26.7 larvae (Table 2) while the mean fertility was 43.6 larvae. Similarly, IT90K-76 showed the maximum hindrance to larval development with 9.0 day for larval development. The shortest duration was 6.0 days in KANANNADO and TVX 3236 while the mean was 7.8 days. The mean adult longevity was 17.3 days with aphids surviving on IT90K-76 for a maximum of 10.0 days and the longest days was 24.3 days in TVX 3236. The average life span of the aphids was 25.14 days. Aphids survived longest on TVX 3236 (30.3 days) and the shortest number of days was 19.0 days in IT90K-76. The multiplication rate was highest on KANANNADO and lowest on IT90K-76. Similarly, highest the intrinsic rate of increase was 0.225 on KANANNADO and the lowest of 0.117 on IT90K-76. The mean of intrinsic rate of increase was 0.17.
| Table 1: | Mean squares of development parameters in seven genotypes of cowpea |
| **Significant p<0.001 | |
| Table 2: | Mean values of development parameters of aphids in seven cowpea genotypes |
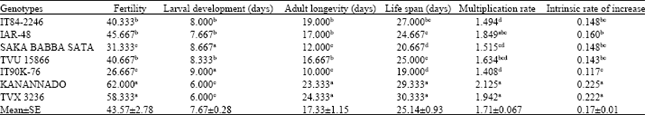 | |
| Means with the same letters are not significantly different | |
DISCUSSION
The result of analysis of variance indicate significant differences (p<0.05) in fertility, larval development, adult longevity, intrinsic rate of increase and multiplication rate of aphids in the seven genotypes studied. Many workers have used this method to determine resistance (especially by antibiosis) particularly when all genotypes support the infestation by aphids as noted in this study. Laamari et al. (2008) reported significantly lower fertility, multiplication rate and duration of reproductive life and longer duration of larval development on some landraces of broad bean to indicate antibiosis-mediated resistance to aphids. Similarly Obopile and Ositile (2010) reported that fecundity; survival and intrinsic rate of natural increase, pre-reproductive period and relative growth rate of cowpea aphid differed significantly among resistant and susceptible varieties of cowpea and further stressed the importance of using these parameters as useful means of characterizing aphid resistance in plants. Similar results were reported by Ofuya (1988); Annan et al. (1997), Le Roux et al. (2004) and Hafiz and Damarany (2006).
This result further revealed a new dimension to resistance of cowpea genotypes to cowpea aphids. Genotypes known to be resistant show similar reaction as others known to be susceptible. An established susceptible genotype IAR-48 had similar level of response as IT84S-2246-4. Indeed it supported higher larval development than IAR-48. On the other hand, TVU-15866, IT90K-76 and a landrace SBS had the greatest level of hindrances to aphid development. It is worthy of notice that IT90K-76 derived it resistance gene from IT86-2246-4, TVU-15866 was brought from Kenya and SBS is a local line. These discrepancies observed in the reaction of the genotypes may suggest a gap in our present knowledge of genetics of aphid resistance in cowpea. Probably there is the existence of modifier to gene for aphid resistance as studies by Myers et al. (1996) tend to suggest. In the same vain, there is the possibility of the presence of the presence of different biotypes of A. craccivora in Zaria. There have been reports of the presence different aphid biotypes in Africa (Ombakho et al., 1987; Martyn, 1991) but so far there was not such report in Nigeria. Similarly most aphid resistant cowpea genotypes developed at IITA have been reported to be susceptible in other countries (Messina et al., 1985; Ofuya, 1997). Kusi et al. (2010) recently reported high susceptibility of IITA lines and suggested the existence of cowpea aphid biotype in northern Ghana which is more virulent than the Nigerian biotypes. It is not clear how an anholytic species like the cowpea aphid suddenly produces variant biotypes. Nevertheless, rare mutations, chromosomal rearrangement and mitotic recombination may give rise to new variants (Hales et al., 1997). Another possible source of different biotype could be the unrestricted movement of research materials from one part of the country to another and beyond. Therefore there is the need to conduct further studies to ascertain cowpea aphid types in and around Zaria.
CONCLUSION
The susceptibility of IT84S-2246-4 indicates the possibility that there is a new variant of cowpea aphid that is capable of infesting known resistances sources. Two genotypes, IT90K-76 and the local line SAKA BABBA SATA were identified as a possible resistance sources because the highest level of antibiosis was recorded with these genotypes. The landrace, SAKA BABBA SATA, has a great potential as sources of additional resistant gene against the Cowpea aphids in Zaria and beyond. This further highlight the importance of local line as potential sources of useful genes for the genetic improvement of crop plants.
REFERENCES
- Annan, I.B., G.A. Schaefers and W.M. Tingey, 1996. Impact of density of Aphis craccivora (Aphididae) on growth and yield of susceptible and resistant cowpea cultivars. Ann. Applied Biol., 128: 183-193.
CrossRef - Annan, I.B., W.M. Tingey and G.A. Schaefers, 1997. Population dynamics and clonal comparisons of Cowpea aphid (Homoptera: Aphididae) on resistant and susceptible cowpea cultivars. Environ. Entomol., 26: 250-255.
Direct Link - Asin, L. and X. Pons, 2001. Effect of high temperature on the growth and reproduction of corn aphids (Homoptera: Aphididae) and implications for their population dynamics on the Northeastern Iberian Peninsula. Environ. Entomol., 30: 1127-1134.
CrossRef - Hafiz, N.A. and A.M. Damarany, 2006. Variation in the susceptibility of some cowpea (Vigna unguiculata (L.) Walp) genotypes to infestation with certain pests in Upper Egypt. Assuit Univ. Bull. Environ. Res., 9: 7-15.
Direct Link - Hales, D.F., J. Tomiuk, K. Wohrmann and P. Sunnuck, 1997. Evolutionary and genetic aspects of aphid biology: A review. Eur. J. Entomol., 94: 1-55.
Direct Link - Kusi, F., D. Obeng-Ofori, S.K. Asante and F.K. Padi, 2010. New sources of resistance in cowpea to the cowpea aphid (Aphis craccivora Koch) (Homoptera: Aphididae). J. Ghana Sci. Assoc., 12: 95-104.
Direct Link - Laamari, M., L. Kheffa and A.C. d'Acier, 2008. Resistance source to cowpea aphid (Aphis craccivora Koch) in broad bean (Vicia faba L.) Algerian landrace collection. Afr. J. Biotechnol., 7: 2486-2490.
Direct Link - Le Roux, V., J. Saguez, C. Vincent and P. Giordanengo, 2004. Rapid method to screen resistance of potato plants against Myzus persicae (Homoptera: Aphididae) in the laboratory. J. Econ. Entomol., 97: 2079-2082.
CrossRef - Messina, F.J., J.A.A. Renwick and J.L. Barmore, 1985. Resistance to Aphis craccivora (Homoptera: Aphididae) in selected varieties of cowpea. J. Entomol. Sci., 20: 263-269.
Direct Link - Obopile, M. and B. Ositile, 2010. Life table and population parameters of cowpea aphid, Aphis craccivora Koch (Homoptera: Aphididae) on five cowpea Vigna unguiculata (L. Walp.) varieties. J. Pest Sci., 83: 9-14.
CrossRef - Ofuya, T.I., 1997. Control of the cowpea aphid, Aphis craccivora Koch (Homoptera: Aphididae), in cowpea, Vigna unguiculata (L.) Walp. Integr. Pest Manage. Rev., 2: 199-207.
CrossRef - Ombakho, G.A., A.P. Tyagi and R.S. Pathak, 1987. Inheritance of resistance to the cowpea aphid in cowpea. Theor. Applied Genet., 74: 817-819.
CrossRef