
M.O. Odeyemi
Department of Zoology, University of Ilorin, Ilorin, P.M.B. 1515, Ilorin, Kwara State, Nigeria
J.O. Fasoranti
Department of Zoology, University of Ilorin, Ilorin, P.M.B. 1515, Ilorin, Kwara State, Nigeria
A.T. Ande
Department of Zoology, University of Ilorin, Ilorin, P.M.B. 1515, Ilorin, Kwara State, Nigeria
I.K. Olayemi
Department of Biological Sciences, Federal University of Technology, Minna, P.M.B. 65, Minna, Niger State, Nigeria
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 15 | Page No.: 735-738







ABSTRACT
This study was aimed at generating baseline information for sustainable genetic improvement of Cirana forda larvae for entomophagy, through the use of gamma irradiation. Eggs of C. forda were irradiated with increasing doses of gamma rays from 0 to 200 Gy and raised through larval instal stages under laboratory conditions. The Body Weight (BW) and Head Capsule Width (HCW) of the larval instar stages were monitored as indices of productivity. Successful larval emergence was recorded for all irradiation doses tested and BW of the 1st and 2nd instar larvae were not significantly (p>0.05) different between the control and treated groups (range = 0.021±0.003 g/larva in the 200 Gy treatment to 0.028±0.003 g/larva in the control group and 0.105±0.003 g/larva in 20 Gy treatment to 0.172±0.009 g/larva in the control group, respectively). On the other hand, BW during the 3rd and 4th larval instars were significantly (p<0.05) lower among the irradiated treatments than control. Pattern of distribution of HCW was different from that of BW; as HCW increased with irradiation dose from 10-50 Gy during the 3rd and 4th larval instars. Also, HCW during the 5th instar larvae among the irradiated treatments (range = 5.256±0.012 to 5.662±0.026 mm) were not higher than that of the 6th instar in the control group (6.065±0.010 mm). These results suggest promising potentials of the use of gamma irradiation in sustainably improving the productivity of C. forda larvae for entomophagy.
PDF Abstract XML References Citation
Received: January 10, 2013;
Accepted: March 04, 2013;
Published: April 04, 2013
How to cite this article
M.O. Odeyemi, J.O. Fasoranti, A.T. Ande and I.K. Olayemi, 2013. Influence of Gamma Irradiation on Productivity Indices of the Edible Emperor Moth Caterpillar, Cirina forda (Lepidoptera: Saturniidae). Pakistan Journal of Biological Sciences, 16: 735-738.
DOI: 10.3923/pjbs.2013.735.738
URL: https://scialert.net/abstract/?doi=pjbs.2013.735.738
DOI: 10.3923/pjbs.2013.735.738
URL: https://scialert.net/abstract/?doi=pjbs.2013.735.738
INTRODUCTION
The larvae of the Emperor moth Cirina forda is the most economically important life stage in the insect’s life cycle; popularly consumed as a rich source of animal protein supplement among native populations in Africa in general and Nigeria in particular (Fasoranti and Ajiboye, 1993; Agbidye et al., 2009). According to Ande (1991), the larvae of C. forda enjoys a high level of acceptability and commercial value due to its appreciable size and availability in large quantities thus, satisfying a major criterion for selecting food insects (Ene, 1963). However, because the larvae are pests of an equally economically important tree species, Vitellaria paradoxa, the insect species development and production are often interrupted by the natives, especially, due to the prolonged larval stage whose gregarious feeding normally coincides with the reproductive season of the host plant. Therefore, there is a need to strike a balance between sustainable production of both C. forda larvae and the sole host plant, V. paradoxa. This may be achieved by shortening the duration of the larval stage, such that the species is able to complete its development within a short period, particularly outside the reproductive season of its host plant. Though, the ecology and biology of C. forda have been well elucidated (Ande, 1991); Odeyemi and Fasoranti, 2000; Odebiyi et al., 2009), little have been reported on sustainable strategies for improving the productivity of the species. Over the years, however, different techniques have been employed in inducing mutational changes that may result in improved performance of insect species generally. One of such techniques is the use of gamma irradiation in achieving desirable genetic changes including, shortening of life stage duration (Elias, 1989; Gonzalez, 1994). Therefore, in order to preserve natives’ culture of entomophagy in the areas of distribution of C. forda, this study was carried out to elucidate the potential of the use of gamma rays in enhancing growth rate and biomass of C. forda larvae, as a prelude to eventual reduction and, perhaps seasonal changes in the life cycle of the species.
MATERIALS AND METHODS
Source and collection of C. forda eggs: C. forda eggs were collected from twigs of Vitellaria paradoxa in a rural community, Enagi, about 100 km from Minna (longitude 6°33'E and latitude 9°37'N), the capital of Niger state, Nigeria. The eggs were maintained in humid boxes before transportation to Energy Laboratory for irradiation with Gamma rays.
Gamma irradiation of eggs and laboratory maintenance of larvae: Collected eggs of C. forda were irradiated with gamma rays at the Centre for Energy Research and Development, Obafemi Awolowo University, Ile Ife, Nigeria. Irradiation doses used were 0, 10, 20, 50, 100, 150, 200 and 300 Gy. The irradiated eggs were transported to the laboratory in humid boxes. The eggs were subsequently incubated for hatching and the larvae were raised according to the methods of Ande (1991).
Measurement of body weight and head capsule width: At the beginning of each larval instar stage, 10 larvae were withdrawn randomly from each treatment group and their weight and Head Capsule Width were measured, following standard procedures (Ande, 1991).
Statistical analysis: Data obtained on growth performance indices were statistically analysed following standard procedures. Differences in mean growth performance parameters, among different groups of irradiated insects, were compared using ANOVA.
RESULTS
The influence of increasing gamma irradiation dose on Body Weight (BW) of C. forda larval stage is presented in Table 1. For all irradiation doses tested, the eggs successfully hatched into larvae (i.e., 1st instar larvae) and there was no significant difference (p>0.05) in 1st instar larval BW between control group and irradiated treatments (range = 0.02±0.003 g/larva in the 200 Gy treatment to 0.028±0.003 g/larva in the control group). However, increasing gamma irradiation inhibited subsequent larval instar development to varying degrees; while, the 150 and 200 Gy treatments did not permit larval development beyond the 1st instar stage, the 100 Gy treatment allowed development to the 3rd instar. The remaining treatment groups (i.e., 10, 20 and 50 Gy) supported larval development to the 5th instar and those of the control group attained the final larval development stage (i.e., 6th instar).
Also, during the 2nd larval instar stage, mean Body Weight (BW) of the control group of larvae were not significantly different from those of the irradiated treatments (range = 0.105±0.003 g/larva in 20 Gy treatment to 0.172±0.009 g/larva in the control group). However, during the 3rd and 4th larval instar stages, with the exception of the 50 Gy treatment, mean larval BW were lower in the irradiated treatments than the control group.
Table 2 highlights larval Head Capsule Width (HCW) responses of C. forda to gamma irradiation. The pattern of distribution of HCW in relation to increasing irradiation doses was different from that of Body Weight, though irradiation treatment had no significant effect on HCW during the 1st instar larval stage (range = 0.926±0.014 mm in control, to 0.955±0.012 mm in the 100 Gy treatment). Unlike BW, Head Capsule Width increased with irradiation dose from 10-50 Gy during the 2nd and 3rd larval instar stages but not significantly different in the 4 and 5th instars. Also, HCWs during the 5th larval instar of the irradiated treatments (range = 5.256±0.072 to 5.662±0.026 mm), were not higher than that of the 6th instar in the control group (6.065±0.10 mm).
| Table 1: | Mean body weight (g) of variously irradiated C. forda for all Instars |
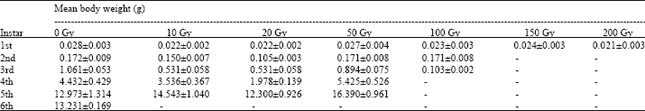 | |
| Table 2: | Mean head capsule width (mm) of variously irradiated C. forda for all Instars |
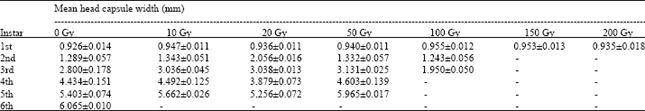 | |
DISCUSSION
Gamma irradiation did not affect egg-hatching and Body Weight (BW) of hatched larvae (1st instar) were not significantly different from those of the control group. Since it was the egg stage of the insect that was exposed to gamma rays, then this result indicates that irradiation had no lethal effect on embryogeny in the species. On the other hand, however, increasing gamma irradiation dose had significant inhibitory effects on subsequent larval instar development. From the economic standpoint, irradiation doses above 50 Gy proved lethal to improved production of C. forda for entomophagy, as such treatments could not support larval development to the 5th instar; a stage considered threshold-acceptability for human consumption (Agbidye et al., 2009).
Though, irradiation treatments (i.e., 10-50 Gy), that produced entomophagy-threshold larval stage (i.e., 5th instar), could not support total larval development, they nevertheless produced significantly heavier larvae at the 5th instar (as much as 16.390±0.961 in the 50 Gy treatment), than those recorded for the 6th instar larvae in the control group (i.e., 13.231±0.169 g/larva). The significant effects of gamma irradiation on the development of C. forda was extended to the 2nd instar stage; but beyond this stage, the effects became more apparent. However, while the effects on Body Weight were mostly subtractive during the 3rd and 4th larval instars when compared with the control, the reverse was the case during the 5th instar stage. These observations, in addition to confirming the non-lethal effects of gamma rays on embryony and egg-hatching, may mean that gamma irradiation disrupted larval growth metabolic activities during the early larval stage (i.e. 3rd and 4th instars), especially feeding and food processing and/or utilization activities. This finding has great ecologic and economic implications. The 3rd and 4th larval instars are the most voracious and disruptive phase of C. forda against Vitellaria paradoxa. Therefore if gamma irradiation checks feeding activities of the insect pest during its most destructive phase but still produce much heavier larvae during the 5th instar, as indicated by the results of this study, may mean effective pest control with an added advantage of enhanced yield for entomophagy. Again, Head Capsule Width during the 1st instar larval stage were not significantly different between the control and irradiated groups of larvae. This result, perhaps, further confirms that gamma irradiation has no lethal effect on embryonic development of C. forda.
The results of this study showed that larval Body Weight and Head Capsule Width responded differently to increasing gamma irradiation dose. This disparity may be explained by the differential physiological bases of the two variables. While, biomass in wet organic matter is highly influenced by its moisture retentive capacity, Head Capsule in insects is more of a function of the degree of sclerotization during ecdysis. Therefore, it may mean that these two physiological processes respond differently to gamma irradiation. Also, unlike its effect on body weight, gamma irradiation treatment did not produce wider Head Capsule in its terminal larval stage (i.e., 5th instar), than that of the control group (i.e., 6th instar). Fortunately, such an effect may not diminish from the potentials of gamma irradiation in producing increased C. forda biomass for entomophagy. According to Agbidye et al. (2009), the nutritional constituents of C. forda larvae contained mainly in the thorax and abdomen and not the head capsule.
CONCLUSION
The findings of this study showed that gamma irradiation has little or no effects on embryony and, perhaps, early larval development in C. forda. Its effects on older larval instars may, however, be subtractive or otherwise, depending on irradiation dose. On the whole, Gamma irradiation shows promising potentials for improving the biomass of C. forda larvae. However, the dose of irradiation must be carefully selected to balance the subtractive effects observed in the early larval stages against the anti-ecdysis effects of high gamma irradiation doses. The optimum irradiation dose (i.e., that will promote increased biomass accumulation and complete larval development), may lie between 20 and 50 Gy. The findings of this study should provide baseline information for sustainable production of C. forda larvae for entomophagy, though gamma irradiation genetic improvement.
ACKNOWLEDGMENT
We are grateful to the Management of Energy Research and Development Center, Obafemi Awolowo University, Ife, Nigeria, for assistance rendered regarding gamma irradiation of C. forda eggs.
REFERENCES
- Agbidye, F.S., T.I. Ofuya and S.O. Akindele, 2009. Marketibility and nutritional qualities of some edible forest insects in Benue state, Nigeria. Pak. J. Nutr., 8: 917-922.
Direct Link - Fasoranti, J.O. and O.O. Ajiboye, 1993. Some edible insects of Kwara state Nigerria. Am. Entomol., 39: 113-116.
Direct Link - Gonzalez, A.J., 1994. Biological effects of low doses of ionizing radiation: A fuller picture. IAEA Bull., 4: 37-45.
Direct Link