
Ismail O. Ishola
Department of Pharmacology, Faculty of Basic Medical Sciences, College of Medicine, University of Lagos, PMB 12003, Idi-Araba, Lagos, Nigeria
Sunday O. Olayemi
Department of Pharmacology, Faculty of Basic Medical Sciences, College of Medicine, University of Lagos, PMB 12003, Idi-Araba, Lagos, Nigeria
Abidemi R. Idowu
Department of Pharmacology, Faculty of Basic Medical Sciences, College of Medicine, University of Lagos, PMB 12003, Idi-Araba, Lagos, Nigeria
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 15 | Page No.: 701-710







ABSTRACT
Crinum glaucum A. Chev (Amaryllidaceae) (CG) is a bulbous plant widely used in folk medicine in the treatment of cough, asthma and convulsions. This study was carried out to investigate the anticonvulsant, anxiolytic and hypnotic effects of the aqueous bulb extract of C. glaucum and its possible mechanism (s) of action. The anticonvulsant activity of C. glaucum extract (400-1200 mg kg-1 p.o.) was investigated using picrotoxin, strychnine, isoniazid, pentylenetetrazol and N-methyl-D-aspartate (NMDA)-induced seizures in mice while the elevated plus maze test (EPM) and hexobarbitone-induced sleeping time (HIST) were used to evaluate the anxiolytic and hypnotic effects, respectively. Animals were pretreated with flumazenil (3 mg kg-1; i.p. GABAA receptor antagonist), cyproheptadine (4 mg kg-1; i.p. 5-HT2 receptor antagonist), L-arginine (500 mg kg-1; p.o. Nitric Oxide (NO) precursor) and L-Nitroarginine (L-NNA) (10 mg kg-1 i.p. Nitric Oxide Synthase (NOS) inhibitor) were used to investigate the probable mechanism (s) of anticonvulsant activity. Oral administration of CG significantly (p<0.001) delayed the onset of seizures induced by picrotoxin, strychnine, isoniazid and pentylenetetrazol with peak effect at 1200 mg kg-1 in comparison to control groups. CG (800 and 1200 mg kg-1) strongly antagonized NMDA-induced turning behavior. Pretreatment of mice with cyproheptadine could not reverse the anticonvulsant effect of CG. However, pretreatment with flumazenil and L-NNA significantly (p<0.05) reversed the anticonvulsant effect of CG while L-arginine pretreatment significantly (p<0.001) delayed the onset of seizures when compared with control and extract (1200 mg kg-1 only). CG potentiated hexobarbitone-induced sleeping time with peak effect at 400 mg kg-1 and also significantly (p<0.05) increased open arm exploration in EPM and had its peak anxiolytic effect at 100 mg kg-1. The data obtained suggests that aqueous bulb extract of Crinum glaucum possess anticonvulsant, anxiolytic and hypnotic activities which involve an interaction with GABAergic, nitrergic and glutaminergic systems to exert its effects.
PDF Abstract XML References Citation
Received: December 07, 2012;
Accepted: February 19, 2013;
Published: April 04, 2013
How to cite this article
Ismail O. Ishola, Sunday O. Olayemi and Abidemi R. Idowu, 2013. Anticonvulsant, Anxiolytic and Hypnotic effects of Aqueous Bulb Extract of Crinum glaucum A. Chev (Amaryllidaceae): Role of GABAergic and Nitrergic Systems. Pakistan Journal of Biological Sciences, 16: 701-710.
DOI: 10.3923/pjbs.2013.701.710
URL: https://scialert.net/abstract/?doi=pjbs.2013.701.710
DOI: 10.3923/pjbs.2013.701.710
URL: https://scialert.net/abstract/?doi=pjbs.2013.701.710
INTRODUCTION
Crinum glaucum A.Chev (Amaryllidaceae) is a bulbous plant widely used in folk medicine in West Africa. The common English names are river lily, String-lily, swamp-lily, Crinum lily and Spider lily. In Zulu tribe it’s known as “umNduze”. In Nigeria, it’s known as “Isumeri” (Gbile, 1984) in the Yoruba speaking regions, “Ede chukwu” or “Ede mmo” (God’s Cocoyam) in Igbo speaking regions and “Albasar kwa’adi” (Frog’s onion) in Hausa speaking regions. The bulbs of Crinum glaucum is used in Southwestern Nigeria as an effective remedy in the relief of cough, asthma and convulsions by traditional medicine practitioners (Burkill, 1985; Okpo and Adeyemi, 2002). They are also used as anti-helminthic and emetics (Oliver, 1959) and in the treatment of sores (Kokwako, 1976), sexually transmitted diseases and backache (Duri et al., 1994). It is cut into small pieces and soaked in water for three days; the liquid (aqueous extract) is then decanted and drank to cure these ailments. The bulb is also used in Southern Nigeria for memory loss and other mental symptoms associated with ageing (Houghton et al., 2004). The aqueous extract is widely used in folk medicine in West Africa as an antispasmodic. Studies have shown a non-specific relaxant effect of Crinum glaucum on gastrointestinal smooth muscles (Okpo and Adeyemi, 1998), analgesic and anti-inflammatory effects (Okpo et al., 2001) and antiallergic effect (Okpo and Adeyemi, 2002). Based on literature search, no study has been carried out to scientifically validate the folkloric uses of Crinum glaucum in the treatment of convulsive and anxiety disorders. Hence, this study was carried out to investigate the anticonvulsant, anxiolytic and hypnotic effects of the aqueous bulb extract of Crinum glaucum in mice and its probable mechanism(s) of action.
MATERIAL AND METHODS
Plant material: Fresh bulbs of Crinum glaucum were purchased from herb sellers in Mushin market, Mushin, Lagos, Nigeria and botanical authentication was carried out by Mr. T.K. Odewo of the Herbarium in the Department of Botany, Faculty of Science, University of Lagos, Akoka, Nigeria, where the herbarium voucher was deposited with the voucher specimen number LUH 5082.
Drugs: Pentylenetetrazol (PTZ), picrotoxin, L-arginine, L-NG-Nitroarginine, strychnine, N-methyl-D-aspartic acid (NMDA), hexobarbitone and cyproheptadine (Sigma Aldrich, St. Louis, MO, USA), phenobarbitone (May and Baker, Lagos, Nigeria), flumazenil (Hikma Farmaceutical, Portugal, S.A.), diazepam (Swipha pharmaceuticals, Nigeria), isoniazid (Mancleods Pharmaceuticals Ltd., India) and normal saline (Unique pharmaceutical limited, Lagos, Nigeria).
Animals: Experimental animals Albino mice of both sexes weighing between 20-25 g were obtained and kept at the Laboratory Animal Centre of the College of Medicine, University of Lagos. The animals maintained under standard environmental conditions had free access to standard diet (Pfizer Feeds, PLC, Lagos, Nigeria) and water ad libitum. The experimental procedures were carried out in accordance with the United States National Institute of Health Guidelines for Care and Use of Laboratory Animals in Biomedical Research (NIH, 1985).
Preparation of plant extract: The fresh bulbs of Crinum glaucum (1.78 kg) were cut into small pieces and soaked in 5 L of distilled water. On day 4, the liquid was decanted and filtered. The filtrate was evaporated to dryness in an oven set at 40°C. A dark brown sticky extract was obtained and stored in airtight bottles in a refrigerator until they were ready for use. The dried extract obtained was weighed. The percentage yield was 3.26%.
Acute toxicity testing: Toxicity of plant extract was determined using the method described by Adeyemi et al. (2007). Mice were fasted for 12 h, divided into five groups: Group I-IV were given CG (1, 2, 4 and 8 g kg-1; p.o) and Group V-normal saline (10 mL kg-1; p.o). Mice were observed for behavioral changes and toxic symptoms for the first 2 h after administration and mortality for a period of 14 days.
Anticonvulsant test
Picrotoxin-or isoniazid-induced convulsions: For each chemoconvulsant 60 mice were used (n = 10). Normal saline (10 mL kg-1, p.o) was given to control group, phenobarbitone (40 mg kg-1, i.p.) was given as a reference standard and CG (200-1200 mg kg-1, p.o.). Thirty minutes (i.p.) or 1 h (p.o.) after treatment, the mice were given picrotoxin (7 mg kg-1, i.p.) or Isoniazid (250 mg kg-1, i.p., seizure consisted of clonic tonic convulsion and wild running around and death). The presence or absence of clonic convulsions was noted for 60 min following administration of convulsants and percentage protections were recorded (Perazzo et al., 2003; Bernasconi et al., 1988).
Strychnine-pentylenetetrazol-induced seizures: For each chemoconvulsant 35 mice were used (n = 7). Normal saline (10 mL kg-1, p.o) was given to control group, phenobarbitone (40 mg kg-1, i.p.) was given as a reference standard and CG (400-1200 mg kg-1, p.o.). Thirty minutes (i.p.) or 1 h (p.o.) after treatment, the mice were given strychnine (5 mg kg-1, i.p.) or pentylenetetrazol (90 mg kg-1, i.p.). The latency to convulsion and percentage protection were recorded (Adeyemi et al., 2007; Gupta et al., 2012).
NMDA induced seizures in mice: Thirty five mice of either sex were randomly allotted to 5 groups (n = 7). Treatment was carried out as follows: Group 1: Normal Saline (NS) (10 mL kg-1 p.o.), Group 2: Phenobarbitone (40 mg kg-1 i.p.) and Group 3-5: CG (400, 800 and 1200 mg kg-1, p.o.). Mice were injected (i.p) with NMDA, 100 mg kg-1, 1 h after intraperitoneal or oral administration of the extract and were observed for 30 min. Turning behaviour was characterized by two consecutive 360° cycles fulfilled by the same animal (Velisek, 2006). Animals that did not exhibit turning behaviour within the 30 min observation period were declared protected. The time of onset of this behaviour in non-protected animals were recorded.
GABAergic pathway involvement: Fourty albino mice of either sex were randomly allotted to 4 groups (n = 10). They were pretreated with flumazenil (3 mg kg-1, i.p.). 15 min after pretreatment, Group I-Mice were given normal saline (10 mL kg-1, p.o.)., Group II-Phenobarbitone (40 mg kg-1, i.p.)., Group III and IV-CG (800 and 1200 mg kg-1). Thirty minutes (i.p.) or 1 hour (p.o.) after treatment, the mice were given picrotoxin (7 mg kg-1, i.p.). The latency to convulsion and percentage protection were recorded (Nogueira and Vassilieff, 2000).
Serotonergic pathway involvement: Thirty albino mice of either sex were randomly allotted to 3 groups (n = 10). They were pretreated with cyproheptadine (4 mg kg-1, p.o). 15 minutes after pretreatment, Group I-Mice were given normal saline (10 mL kg-1, p.o.)., Group II-Phenobarbitone (40 mg kg-1, i.p.)., Group III-CG (1200 mg kg-1). Thirty minutes (i.p.) or 1 hour (p.o.) after treatment, the mice were given picrotoxin (7 mg kg-1, i.p.). The latency to convulsion and percentage protection were recorded (Michael, 2006).
Nitric oxide pathway involvement: Fourty albino mice of either sex were randomly allotted to 4 groups (n = 10). They were pretreated with nitric oxide synthase inhibitor-L-NNA (10 mg kg-1, p.o) 15 minutes after pretreatment, the animals were given normal saline or CG 1200 mg kg-1. The effect of CG on nitric oxide synthesis was investigated through oral pretreatment with L-arginine (500 mg kg-1, p.o.). 15 minutes after pretreatment, mice were given normal saline (10 mL kg-1, p.o.) or CG (1200 mg kg-1). 1 hour (p.o.) after treatments, the mice were given picrotoxin (7 mg kg-1, i.p.). The latency to convulsion and percentage protection were recorded (Paul and Subramanian, 2002).
Hexobarbitone sleeping time: Five groups of 7 mice each were given normal saline (10 mL kg-1, p.o.), CG (200, 800 and 1600 mg kg-1, p.o.) and diazepam (3 mg kg-1, p.o.) respectively. One hour later, hexobarbitone (100 mg kg-1, i.p.) was administered to each mouse in turn. The mice were placed on their backs in separate chambers and the duration of loss of righting reflex starting at the time of hexobarbitone administration until they regained their righting reflexes were recorded. When there was any doubt, the animal was placed gently on its back again and if it righted itself within 1 min, this was regarded as the end point (Vogel and Vogel, 1997; Mujumdar et al., 2000).
Elevated plus maze test: The EPM testing procedure was as described by Pellow et al. (1985). The equipment was made of wood and had four arms of equal dimensions (50x12 cm). Two of the arms were enclosed by 40 cm high walls and were arranged perpendicularly to two opposite open arms. The apparatus was elevated 50 cm above the floor. One hour following oral administration of CG (200, 800 and 1600 mg kg-1, p.o.), diazepam (1.5 mg kg-1, p.o.) and normal saline (10 mL kg-1, p.o.), mice were placed in the centre of the maze facing an open arm. The cumulative time spent in the open or closed arm were recorded for 5 min. The maze was cleaned with an alcohol solution after each trial.
Statistical analysis: Results are expressed as Mean±S.E.M., statistical analysis of data was done by a means of One-Way analysis of variance (ANOVA) followed by Tukey’s multiple comparison tests. A level of significance (p<0.05, 0.01 or 0.001) was considered for each test.
RESULTS
Acute toxicity study: Oral administration of the extract (1, 2, 4 and 8 g kg-1) caused 20, 40, 100 and 100% death of animals respectively. These percentage responses were converted to probit values which were plotted against log doses of the aqueous extract. The LD50 obtained (after extrapolation) was 1.479 g kg-1. The aqueous extract was found to cause behavioural toxic effects which include; calmness, hypoactivity, abdominal writhing and stooling.
Anticonvulsant activity
Picrotoxin induced seizures: Picrotoxin (7 mg kg-1 i.p.) elicited seizures in all the seven mice used in normal saline control group with an onset of 240.00 ± 26.83 s and 100% mortality. Aqueous bulb extract of Crinum glaucum (400 and 1200 mg kg-1) significantly (p<0.05; p<0.01) delayed the onset of picrotoxin elicited clonic convulsion (468.00±81.39 and 528.00±81.39 s respectively) and 40% protection in comparison to normal saline treated group in a dose dependent manner. Phenobarbitone (40 mg kg-1) significantly (p<0.01) delayed the onset of seizures (542.20±50.91 s) with 100% protection (Table 1).
Isoniazid induced seizures: Isoniazid (250 mg kg-1 i.p.) elicited clonic tonic convulsion in all the animals used. The normal saline control group produced convulsion and showed an onset of 32.80±2.55 min. The aqueous extract of C. glaucum (400-1200 mg kg-1) significantly (p<0.01; 0.001) delayed the onset of isoniazid induced seizures from 32.80±2.55 min in control to 43.20±0.83, 40.60±0.55 and 53.00±0.97 min respectively and shows dose-dependent increase in the anticonvulsant activity. Similarly, phenobarbitone 40 mg kg-1 pretreatment significantly (p< 0.001) increased the time to the onset of isoniazid induced seizures from 32.80±2.55 min to 47.00±2.10 min (Table 1).
Strychnine induced seizures: Strychnine (5 mg kg-1) produced tonic convulsion in 100% of the mice used. CG (400, 800 and 1200 mg kg-1) pretreatment significantly (p<0.05, 0.01, 0.001) increase the time of onset of tonic seizure from 29.20±0.40 sec in normal saline treated control to 67.60±8.69, 78.20±3.54 and 107.40±8.38 s, respectively (Table 2).
| Table 1: | Effect of aqueous bulb extract of Crinum glaucum (CG) in picrotoxin (PCT) and isoniazid (INH)-induced seizures in mice |
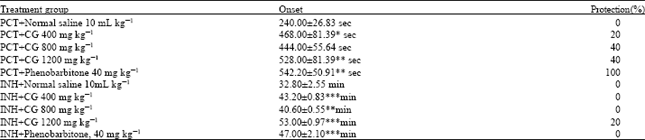 | |
| The data were expressed as Mean±SEM (n = 10) *p<0.05. **p< 0.01; ***p<0.001 versus normal saline treated control group, the data was analyzed using One Way ANOVA followed by Tukey’s multiple comparison post hoc tests | |
| Table 2: | Effect of aqueous bulb extract of Crinum glaucum (CG) in Strychnine (STR)-, Pentylenetetrazole (PTZ)-and N-methyl-D-aspartic acid (NMDA) induced seizures in mice |
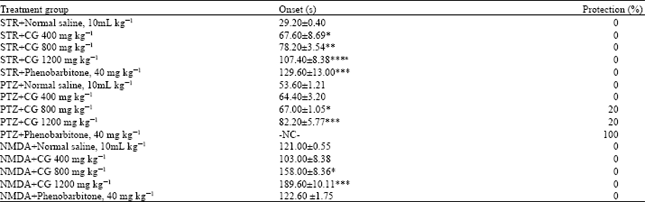 | |
| NC: No convulsion, the data were expressed as mean±SEM (n = 7). *p<0.05, **p<0.01, ***p<0.001 versus normal saline treated control group, ap<0.05 versus CG 400 mg kg-1 treated group, the data was analyzed using One Way ANOVA followed by Tukey’s multiple comparison Post hoc tests | |
Phenobarbitone (40 mg kg-1) also significantly (p<0.001) increased the onset of tonic from 29.20±0.40 s in saline treated control to 129.60±13.00 s.
Pentylenetetrazole induced seizures: PTZ (90 mg kg-1, i.p.) elicited tonic convulsion in 100% of the animals used in normal saline treated control. CG (800 and 1200 mg kg-1, p.o.) dose dependently delayed the onset of PTZ (90 mg kg-1, i.p.)-induced clonic convulsion significantly (p<0.05, 0.001), respectively (Table 2).
The extract at 800 mg kg-1 increased the time of onset of clonic seizure from 53.60±1.21 s in saline treated control to 67.00±1.05 sec and provided 20% protection while the extract at 1200 mg kg-1 protected 20% of mice against seizure and increased the time of onset of clonic seizure from 53.60±1.21 sec in saline treated control to 82.20±5.77 sec. Phenobarbitone (40 mg kg-1 i.p.) profoundly antagonized the clonic seizures induced by PTZ and also protected all the mice against seizures (Table 2).
NMDA induced seizures: NMDA-induced seizures in rodents have previously been proposed as a model of refractory seizures and are significantly suppressed by NMDA receptor antagonists (Velisek, 2006). NMDA (100 mg kg-1, i.p.) elicited turning behaviour in 100% of normal saline treated control mice with an onset of 121±0.55 sec to tonic seizures. CG (800 and 1200 mg kg-1, p.o.) did not affect the incidence of NMDA (100 mg kg-1, i.p.) induced turning behaviour but significantly (p<0.05) delayed the onset of turning from 121.00±0.55 sec in control to 158.00±8.36 and 189.60±10.11, respectively (Table 2). However, oral administration of phenobarbitone failed to increase the onset of turning behaviour.
Possible involvement of GABAergic, serotonergic and nitric oxide pathways in the anticonvulsant activity of Crinum glaucum: Intraperitoneal pretreatment with flumazenil (3 mg kg-1) reversed the protective effect of Crinum glaucum. Pretreatment with cyproheptadine (3 mg kg-1) followed by CG 1200 mg kg-1 showed significant (p< 0.05) reduction in the latency to seizure in comparison to CG (1200 mg kg-1) only treated group. Nitric oxide synthase inhibitor (L-NNA) significantly (p<0.01) reduced the latency to convulsion when compared with the normal saline treated group. However, nitric oxide precursor co-treatment (L-arginine (500 mg kg-1) +CG 1200 mg kg-1) produced a significant (p< 0.001) increase in the time of onset of clonic seizure in comparison to L-NNA + CG (1200 mg kg-1) co-treated and 40% protection was observed (Table 3).
| Table 3: | Effect of co treatment with flumazenil, cyproheptadine, L-arginine and L-NG-Nitroarginine and aqueous bulb extract of Crinum glaucum (CG) in picrotoxin (PCT)-induced seizures in mice |
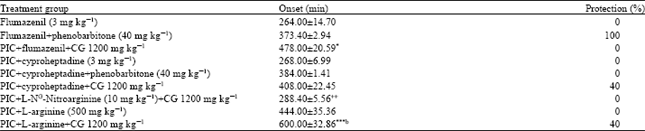 | |
| The data were expressed as mean±SEM (n = 10) *p<0.05; **p<0.01; ***p<0.001 versus normal saline treated control group, +p<0.05; ++p<0.01 versus C. glaucum 1200 mg kg-1 treated group; ap< 0.01, bp<0.001 versus L-NG- Nitroarginine+C. glaucum 1200 mg kg-1 treated group, the data was analyzed using One Way ANOVA followed by Tukey’s multiple comparison post hoc tests | |
| Table 4: | Effect of aqueous bulb extract of Crinum glaucum (CG) on potentiation of hexobarbitone sleeping time |
 | |
| The data were expressed as Mean±S.E.M (n = 7). ***p< 0.001 versus normal saline treated control group, the data was analyzed using One Way ANOVA followed by Tukey’s multiple comparison post hoc tests | |
| Table 5: | Effect of Crinum glaucum on elevated plus maze test |
 | |
| The data were expressed as Mean±SEM (n = 7), **p<0.01, ***p< 0.001 versus normal saline treated control group, #p< 0.001 compared to diazepam 1.5 mg kg-1, ap< 0.001 compared to CG 100 mg kg-1, the data was analyzed using One Way ANOVA followed by Tukey’s multiple comparison post hoc tests | |
Effect of aqueous bulb extract of Crinum glaucum potentiation of hexobarbitone-induced sleeping time: The normal saline treated group (control group) gave an onset of sleep of 3.92±0.10 min and duration of 39.79±0.72 min when hexobarbitone was administered (Table 4). The aqueous extract of C. glaucum (200-1200 mg kg-1) significantly (p< 0.001) reduced the latency to sleep while it also significantly (p<0.001) prolonged the duration of sleep induced by hexobarbitone. The peak hypnotic effect was obtained at 400 mg kg-1 CG treated where the onset of sleep was significantly (p<0.001) reduced from 3.92±0.10 min in saline treated to 1.52±0.01 min while the duration of sleep was significantly (p<0.001) increased from 39.79±0.72 min in saline treated to 77.50±0.81 min. similarly, oral administration of diazepam produced significant (p<0.01) reduction in the onset of sleep (2.40±0.24) and prolonged the duration of sleep (82.00±7.19 min) induced by hexobarbitone in comparison to saline control group. The hypnotic effect of diazepam was comparable to that of CG (400 mg kg-1) (Table 4).
Effect of aqueous bulb extract of Crinum glaucum on elevated plus maze: The normal saline treated animals (control group) showed that 23.40±2.32 s was spent in the open arms (Table 5). The aqueous extract Crinum glaucum produced significant (p<0.01-0.001) increase in the time spent in the open arms compared to control. Diazepam produced significant (p<0.001) increase in the time spent in the open arms. The extract peak anxiolytic effect (highest time spent in open arms) was produced at 100 mg kg-1 (124.00±1.18 s) and it is significantly (p<0.05) higher than that produced by diazepam (84.4±2.84 s) (Table 5).
DISCUSSION
Most of conventional antiepileptic drugs are associated with many side effects such as neurotoxic effects, cognitive deficits and teratogenic effects which decrease their clinical utility (Trimble, 1987; Yerby, 1988; Meador et al., 1990). Recently, the search for novel pharmacotherapy from medicinal plants for neurological and psychiatric diseases has progressed significantly owing to their less side effects and better tolerability (Zhang, 2004). In the present study the anticonvulsant, anxiolytic and hypnotic activities of bulb extract of C. glaucum have been studied. The results obtained in this study demonstrate that the extract dose dependently increased the onset of convulsion induced by picrotoxin, isoniazid, strychnine, pentylenetetrazole and NMDA as well as significant prolongation of sleeping time and time spent in open arms in hexobarbitone induced hypnosis and elevated plus maze tests, respectively.
Picrotoxin exerts its convulsant effect by blocking GABAA receptor-linked chloride ion channel which, normally opens to allow increased chloride ion conductance into the brain cells following activation of GABAA receptors by GABA (Nicoll, 2001; Rang et al., 2003). Phenobarbitone is a standard antiepileptic drug that has been shown to exert its effect by enhancing GABA-mediated inhibition in the brain (Porter and Meldrum, 2001). In picrotoxin-induced seizures, the plant extract at doses 400 and 1200 mg kg-1 significantly delayed the onset of picrotoxin elicited clonic seizures in mice. The extract at 800 and 1200 mg kg-1 protected 40% of the mice while 400 mg kg-1 protected 20% of the mice. Phenobarbitone significantly delayed the onset of clonic seizures while also protecting 100% of the mice. The plant extract at 1200 mg kg-1 produced the highest onset of seizures and also protected 40% of the animals from seizures suggesting that the extract exerts its effect possibly by the enhancement of GABA neurotransmission to attenuate picrotoxin induced convulsion.
Isoniazid exerts its convulsive effect by inhibiting GABA synthesis (Costa et al., 1975). It is a potent monoamine oxidase (MAO) inhibitor and a glutamic acid decarboxylase (GAD) inhibitor (enzyme involved in GABA synthesis) thus increases the brain monoamine content and inhibited GABA synthesis respectively thereby leading to CNS excitation and convulsions (Wood and Peesker, 1973; Marcus and Coulsto, 1985). Isoniazid induced seizures was carried out to further confirm the GABA enhancing activity of the plant extract. The extract (400-1200 mg kg-1) when compared to control treated produced significant increase in the time of onset of clonic seizures, this shows dose-dependent increase in the anticonvulsant activity. Phenobarbitone did not affect the incidence of seizures or death but significantly delayed the onset of seizures.
Strychnine is a selective competitive antagonist that blocks the inhibitory effect of glycine at all glycine receptors (Rajendra et al., 1997; Parmar and Prakash, 2006). In strychnine induced seizures, the plant extract did not protect the animals from seizure or death but increased the seizure onset significantly in a dose dependent manner. Phenobarbitone did not affect the incidence of seizures nor prevent death but delayed the onset of seizures significantly when compared to control (normal saline) group. The peak increase in onset of seizure by CG was obtained at 1200 mg kg-1. This shows that the extract may have minimal glycine receptor antagonism.
PTZ mostly exerts its action by inhibiting the t-butyl-bicyclo-phosphorothionate / picrotoxin site of the GABAA receptor (Velisek, 2006) and thereby preventing chloride ion conductance into the brain cells. There is also the involvement of Na+/Ca2+ exchanger (NCX) in PTZ induced convulsion (Salto et al., 2009). In PTZ induced seizures, phenobarbitone completely inhibited the incidence of seizures. The plant extract however did not protect the animals from seizures, but increased the seizure onset significantly with 20% protection at 800 and 1200 mg kg-1. This result shows that the effect of phenobarbitone and the extract are not comparable, but the effect of the plant extract in this model cannot be totally ignored as they still lowered onset of convulsion and protected 20% of the animals. It can then be said that the extract is not very effective in preventing PTZ induced seizures and this could be due to the additional mechanisms by which PTZ acts to induce convulsion other than inhibiting GABAA receptors.
NMDA is an agonist that produces effects similar to glutamic acid at the NMDA receptors and exerts its convulsant effect by activating the receptors to enhance glutaminergic neurotransmission (Watkins and Evans, 1981; Chapman and Meldrum, 1993). NMDA-induced seizures in rodents have previously been proposed as a model of refractory seizures and are significantly suppressed by NMDA receptor antagonists (Velisek, 2006). In NMDA induced seizures, the plant extract did not protect the animals from seizures and death but increased the seizure onset significantly (p<0.05; 0.001) in a dose dependent manner (at doses 800 and 1200 mg kg-1), respectively. The plant extract at 1200 mg kg-1 produced a delayed onset and shorter duration of seizures when compared to phenobarbitone and so had a better anticonvulsant effect in this model, showing that this is the most effective dose. This shows that phenobarbitone does not act by blocking glutamate transmission and that the extract also possesses minimal anti-glutaminergic activity compared to phenobarbitone.
The probable mechanism of anticonvulsant effect of bulb extract of C. glaucum was investigated in picrotoxin induced seizure using the most effective dose of the extract (1200 mg kg-1). To confirm GABA or effect at allosteric benzodiazepine receptor on GABA. Animals were pretreated with flumazenil (a benzodiazepine receptor antagonist), Flumazenil had no significant anticonvulsant activity when it was administered alone. Phenobarbitone and flumazenil co-treated group gave no significant reduction in its onset of convulsion when compared to phenobarbitone treated group only and also gave 100% protection, implying that flumazenil has relatively no effect on the action of phenobarbitone. However, flumazenil pretreatment reversed the anticonvulsant effect of the bulb extract of C. glaucum. This further confirm that C. glaucum may be acting as an agonist of the GABAA-benzodiazepine receptor complex.
Despite the similarity in the seizure phenotype induced by PTZ and picrotoxin, marked difference between their pharmacological profiles have been observed, like carbamazepine, a clinically used anticonvulsant is effective against seizures induced by MES and picrotoxin but not in seizures induced by PTZ (White, 1998). Carbamazepine has also been reported to increase serotonergic neurotransmission as one of its proposed anticonvulsant mechanism (Dailey et al., 1995; Bagdy et al., 2007). In our studies cyproheptadine (5-HT2 receptor antagonist) pretreatment failed to reversed the anticonvulsant activity of C. glaucum extract ruling out the involvement of serotonergic neurotransmission in its anticonvulsant activity.
The possible involvement of NO in the anticonvulsant activity of C. glaucum was investigated by administration of NO precursor (L-arginine 500 mg kg-1) and NOS inhibitor (L-NNA 10 mg kg-1).
It has been shown that 30 and 60 min pretreatment of 1000 mg kg-1 and not 500 mg kg-1 of L-arginine inhibited convulsions induced by picrotoxin, while a Nitric Oxide Synthase (NOS) inhibitor, N-nitro-L-arginine methyl ester (L-NAME) increased convulsant action of picrotoxin (Paul and Subramanian, 2002). The concentrations of nitric oxide (NO) and Gamma Aminobutyric Acid (GABA) were increased in the brain 30 and 60 min after administration of 1000 mg kg-1 and not 500 mg kg-1 of L-arginine. However, L-NAME decreased brain GABA concentration and increased the convulsant action of picrotoxin. Further, L-NAME pretreatment prevented L-arginine (1000 mg kg-1) from producing anticonvulsant and NO and GABA increasing effects (Paul and Subramanian, 2002). In this study, L-arginine 500 mg kg-1 alone produced seizures in all mice used, with 100% mortality. However, when L-arginine when co administered with the extract (1200 mg kg-1), it produced a significant increase in onset of seizures and 40% protection when compared with extract only treated group. In addition, L-NG-Nitroarginine pretreatment reduced the antiseizure activity of the extract. These results support the suggestions of previous investigators that NO modulates the concentration of GABA in the brain (Paul and Jayakumar, 2000). NO seems to increase GABA concentration by decreasing GABA transaminase (GABA-T) activity in the brain because a decreased concentration of GABA by an NO decreasing dose of L-NAME coincided with an increased activity of GABA-T in the brain (Paul and Jayakumar, 2000). These observations and the ability of L-NNA in the present study to prevent C.glaucum from inhibiting seizure, provide further support to the suggestion that NO functions as a modulator of GABA concentration in the brain. Further, NO has been found to activate release of GABA from cerebral cortex (Kuriyama and Ohkuma, 1995), striatum (Lonart et al., 1992) and hippocampus (Segovia and Mora, 1998). Conversely, inhibitors of NO synthesis have decreased GABA release in the cerebral cortex (Montague et al., 1994). These results by NO Bie and Zhao (2001), strongly support the suggestion that a functional interaction occurs between NO and GABA in the brain. This shows that the extract probably acts by increasing the concentration of nitric oxide and GABA concentration in the brain of mice.
The aqueous extract produced a significant (p<0.001) reduction in the onset of sleep induced by hexobarbitone at 200-1200 mg kg-1 and significant (p<0.001) prolongation of sleep at doses 400 and 1200 mg kg-1. The prolongation of sleep observed at 400 mg kg-1 is comparable to that produced by diazepam 3 mg kg-1 and therefore when compared to each other, there was no significant difference in the duration of sleep observed in both treatments. Prolongation of hexobarbitone sleeping time indicates a central nervous system depressant activity of the extract (Mujumdar et al., 2000). It also indicates that the extract is an enzyme inhibitor as possibly increased the action of hexobarbitone by preventing its metabolism.
Furthermore, this work demonstrated that the administration of different doses of CG in mice was able to induce anxiolytic effect. The elevated plus maze is considered to be an etiologically valid animal model of anxiety because it uses natural stimuli (fear of a novel open space and fear of balancing on a relatively narrow, raised platform) that can induce anxiety in humans (Dawson and Tricklebank, 1995). An anxiolytic agent increases the frequency of entries into the open arms and increases the time spent in open arms of the EPM. In the present study, oral administration of C.glaucum induced an anxiolytic-like effect in mice, since it increased the number of entries and the time spent on open arms and decreased the time spent in closed arms in the EPM test. In agreement with previously published reports, diazepam increased the duration of time spent on open arms and the number of entries on open arms (Ishola et al., 2012).
The aqueous extract (100-1200 mg kg-1) significantly increased time spent in open arms of the elevated plus maze but no statistical difference in the number of entries (result not shown). The anxiolytic effect peaked at 100 mg kg-1 which was comparable to diazepam.
We have no adequate explanation for the lack of dose dependence in this assay. Further studies to investigate the effect of Crinum glaucum at different intervals of time might disclose a dose-dependent effect.
The result of the acute oral toxicity showed that the extract possesses wide margin of safety which is in agreement with previous study of Okpo et al. (2001).
The phytochemical constituents of the plant include phenols, flavonoids, glycosides and alkaloids. One or more of these substances may be involved in the anticonvulsant activity of the extract.
CONCLUSION
In conclusion, the present study provides scientific evidence for the use of bulb of Crinum glaucum in treatment of epileptic and anxiety disorders by Southwestern Nigerians. Crinum glaucum exerts its anticonvulsant and anxiolytic effects by increasing the concentration, release and the activity of GABA which is a well-documented inhibitory neurotransmitter in the brain possessing an anticonvulsant and anxiolytic properties.
ACKNOWLEDGMENTS
The authors are grateful to Mr. Micah Chijioke of the Department of Pharmacology, College of Medicine, University of Lagos, Lagos, Nigeria, for technical assistance rendered in the course of this research work.
REFERENCES
- Adeyemi, O.O., O.K. Yemitan and O.A. Olayemi, 2007. Sedative and anticonvulsant activities of the aqueous root extract Sanseviera liberica Gerome and Labroy (Agavaceae). J. Ethnopharmacol., 113: 111-114.
CrossRef - Bagdy, G., V. Kecskemeti, P. Riba and R. Jakus, 2007. Serotonin and epilepsy. J. Neurochem., 100: 857-873.
CrossRef - Bernasconi, R., M. Klein, P. Martin, P. Christen, T. Hafne, C. Portet and M. Schmutz, 1988. Gamma-Vinyl GABA: Comparison of neurochemical and anticonvulsant effects in mice. J. Neural Trans., 72: 213-233.
CrossRefDirect Link - Bie, B.H. and Z.Q. Zhao, 2001. Nitric oxide inhibits GABA-evoked current in dorsal root ganglion neuron via PKG-dependent pathway. Brain Res. Bull., 55: 335-339.
CrossRef - Chapman, A.G. and B.S. Meldrum, 1993. Excitatory amino acid antagonists and epilepsy. Biochem. Soc. Trans., 21: 106-110.
PubMed - Costa, E., A. Guidotti and C.C. Mao, 1975. Evidence for involvement of GABA in the action of benzodiazepines: studies on rat cerebellum. Adv. Biochem. Psychopharmacol., 14: 113-130.
PubMedDirect Link - Dailey, J.W., Q.S. Yan, L.E. Adams, J.R. Ryu, K.H. Ko, P.K. Mishra and P.C. Jobe, 1995. Neurochemical correlates of antiepileptic drugs in the Genetically Epilepsy-Prone Rat (GEPR). Life Sci., 58: 259-266.
CrossRef - Dawson, G.R. and M.D. Tricklebank, 1995. Use of the elevated plus maze in the search for novel anxiolytic agents. Trends Pharmacol. Sci., 16: 33-36.
CrossRef - Duri, Z.J., J.P. Scovill and J.W. Huggins, 1994. Activity of a methanolic extract of Zimbabwean Crinum macowanii against exotic RNA viruses in vitro. Phytother. Res., 8: 121-122.
CrossRef - Gupta, G., I. Kazmi, M. Afzal, M. Rahman and S. Saleem et al., 2012. Sedative, antiepileptic and antipsychotic effects of Viscum album L. (Loranthaceae) in mice and rats. J. Ethnopharmacol., 141: 810-816.
CrossRefDirect Link - Houghton, P.J., J.M. Agbedahunsi and A. Adegbulugbe, 2004. Choline esterase inhibitory properties of alkaloids from two Nigerian Crinum species. Phytochemistry, 65: 2893-2896.
CrossRef - Ishola, I.O., M. Chatterjee, S. Tota, N. Tadigopulla, O.O. Adeyemi, G. Palit and R. Shukla, 2012. Antidepressant and anxiolytic effects of amentoflavone isolated from Cnestis ferruginea in mice. Pharmacol. Biochem. Behav., 103: 322-331.
CrossRefDirect Link - Kuriyama, K. and S. Ohkuma, 1995. Role of nitric oxide in central synaptic transmission: Effects on neurotransmitter release. Jpn. J. Pharmacol., 69: 1-8.
CrossRefPubMedDirect Link - Lonart, G., J. Wang and K.M. Johnson, 1992. Nitric oxide induces neurotransmitter release from hippocampal slices. Eur. J. Pharmacol., 220: 271-272.
CrossRef - Meador, K.J., D.W. Loring, K. Huh, B.B. Gallagher and D.W. King, 1990. Comparative cognitive effects of anticonvulsants. Neurology, 40: 391-394.
CrossRefDirect Link - Montague, P.R., C.D. Gancayco, M.J. Winn, R.B. Marchase and M.J. Friedlander, 1994. Role of NO production in NMDA receptor-mediated neurotransmitter release in cerebral cortex. Science, 263: 973-977.
CrossRefPubMedDirect Link - Mujumdar, A.M., D.G. Naik, R.J. Waghole, D.K. Kulkarni and M.S. Kumbhojkar, 2000. Pharmacological studies on Sterculia foetida leaves. Pharma. Biol., 38: 13-17.
PubMed - Nogueira, E. and V.S. Vassilieff, 2000. Hypnotic, anticonvulsant and muscle relaxant effects of Rubus brasiliensis. Involvement of GABAA-system. J. Ethanopharmacol., 70: 275-280.
CrossRefPubMedDirect Link - Okpo, S.O. and O.O. Adeyemi, 2002. The anti-allergic effects of Crinum glaucum aqueous extract. Phytomedicine, 9: 438-441.
CrossRef - Okpo, S.O. and O.O. Adeyemi, 1998. Effects of Crinum glaucum aqueous extract on intestinal smooth muscle activity. Phytother. Res., 12: 413-416.
CrossRef - Okpo, S.O., F. Fatokun and O.O. Adeyemi, 2001. Analgesic and anti-inflammatory activity of Crinum glaucum aqueous extract. J. Ethnopharmacol., 78: 207-211.
CrossRefPubMedDirect Link - Paul, V. and A.R. Jayakumar, 2000. A role of nitric oxide as an inhibitor of γ-aminobutyric acid transaminase in rat brain. Brain Res. Bull., 51: 43-46.
CrossRef - Paul, V. and E.H. Subramanian, 2002. Evidence for an involvement of nitric oxide and gamma aminobutyric acid in the anticonvulsant action of L-arginine on picrotoxin-induced convulsions in rats. Pharmacol. Bioch. Behav., 72: 515-519.
CrossRefDirect Link - Pellow, S., P. Chopin, S.E. File and M. Briley, 1985. Validation of open: Closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci. Methods, 14: 149-167.
CrossRefDirect Link - Perazzo, F.F., J.C.T. Carvalho, J.E. Carvalho and V.L.G. Rehder, 2003. Central properties of the essential oil and the crude ethanol extract from aerial parts of Artemisia annua. J. Pharmacol. Res., 48: 497-502.
CrossRef - Rajendra, S., J.W. Lynch and P.R. Schofield, 1997. The glycine receptor. Pharmacol. Therapeut., 73: 121-146.
CrossRefDirect Link - Salto, R., E. Kaneko, Y. Tanaka, K. Honda and T. Matsuda et al., 2009. Involvement of Na+/Ca2+ exchanger in pentylenetetrazol-induced convulsion by the use of Na+/Ca2+ exchanger knockout mice. Biol. Pharm. Bull., 32: 1928-1930.
CrossRef - Segovia, G. and F. Mora, 1998. Role of nitric oxide in modulating the release of dopamine, glutamate and GABA in striatum of the freely moving rat. Brain Res. Bull., 45: 275-280.
CrossRef - Trimble, M.R., 1987. Anticonvulsant drugs and cognitive function: A review of the literature. Epilepsia, 28: S37-S45.
CrossRef - Watkins, J.C. and R.H. Evans, 1981. Excitatory amino acid transmitters. Ann. Rev. Pharmacol. Toxicol., 21: 165-204.
CrossRefDirect Link - Wood, J.D. and S.J. Peesker, 1973. The role of gaba metabolism in the convulsant and anticonvulsant actions of aminooxyacetic acid. J. Neurochem., 20: 379-387.
CrossRef - Zhang, Z.J., 2004. Therapeutic effects of herbal extracts and constituents in animal models of psychiatric disorders. Life Sci., 75: 1659-1699.
CrossRefDirect Link - Michael, A.R., 2006. Diverse mechanisms of antiepileptic drugs in the development pipeline. Epilepsy Res., 69: 273-294.
CrossRefDirect Link