
B.A. Falana
Department of Anatomy, College of Medicine and Health Sciences, Osun State University, Oshogbo, Nigeria
O.M. Ogundele
Department of Anatomy, College of Medicine and Health Sciences, Afe Babalola University, Ado-Ekiti, Nigeria
F.I. Duru
Department of Anatomy, College of Medicine and Health Sciences, University of Lagos, Lagos Sate, Nigeria
A.A. Oshinubi
Department of Anatomy, College of Medicine and Health Sciences, University of Lagos, Lagos Sate, Nigeria
D.T. Falode
Department of Nursing, College of Medicine and Health Sciences, Afe Babalola University, Ado-Ekiti, Nigeria
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 2 | Page No.: 67-73







ABSTRACT
The germinal epithelium is the delicate epithelial lining of the seminiferous tubule lying on the blood-testes barrier; formed by the sustenacular cells of Sertoli and the adjoining basement epithelium this study addresses the effect of lead (Pb) toxicity on the epithelium and the proliferative effect of Zinc (Zn) and Selenium (Se) administered in trace concentration. Sixty F1 generation adult male Wistar rats were divided into four groups of 15 animals each. Group 1 received normal saline, group 2: 100 mg kg-1 of lead acetate, group 3: 100 mg kg-1 of lead acetate then 2.25 mg kg-1 each of Zinc (Chelated zinc) and Selenium (Sodium Selenium) and group 4: 2.25 mg kg-1of zinc and selenium (Se+Zn). The duration of treatment was 56 days following which the animals were sacrificed on the 57th day and the testes harvested and fixed in Bouin’s fluid. Pb induced toxicity can follow a mitochondria pathway involving Cathepsin D (CAD) or a cytoplasmic pathway involving p53 (protein 53; a 53 KDa nucleolase), the most predominant form of cell death is apoptosis which can result from both pathways. Se+Zn treatment improves proliferation and counters Pb toxicity by substitution, activation of enzymes (radical scavengers and vitamins), growth factors, activation of endothelial factors and activation of oxygen radical scavengers.
PDF Abstract XML References Citation
Received: December 05, 2012;
Accepted: February 09, 2013;
Published: March 14, 2013
How to cite this article
B.A. Falana, O.M. Ogundele, F.I. Duru, A.A. Oshinubi and D.T. Falode, 2013. Role of Se+Zn in Regeneration (Ki-67) Following Pb Toxicity (p53andcad)
in the Germinal Epithelium of Adult Wistar Rats. Pakistan Journal of Biological Sciences, 16: 67-73.
DOI: 10.3923/pjbs.2013.67.73
URL: https://scialert.net/abstract/?doi=pjbs.2013.67.73
DOI: 10.3923/pjbs.2013.67.73
URL: https://scialert.net/abstract/?doi=pjbs.2013.67.73
INTRODUCTION
The germinal epithelium represents the interior of the seminiferous tubule; it houses the cells of the spermatogenic linage (spermatogonia to spermatozoa) and other supporting and secretory cells such as the sustenacular cells of Sertoli and the interstitial cells of Leydig (Xiang et al., 2013). The Sertoli cells are large and possess multiple cellular projections extending from the region of the basement membrane where it forms junctional complexes to the luminal part of the epithelium (Tajaddini et al., 2013). The significance of the junctional complexes formed by the Sertoli cells cannot be over emphasized as it represents the blue print for the blood testes barrier (Su and Liu, 2013). The circulation of nutrients in the germinal epithelium is unique in the sense that the basement membrane and the Sertoli cells are the points of contact to almost every molecule passing on to the delicate spermatogenic cells (Mok et al., 2012). Aside this, adjacent Sertoli cells communicate via gap junctions and synchronize using chemical and ionic signals especially those involving chemokines and chloride ions. Thus, the barrier is the first region to be affected by all toxins and heavy metals including lead (Pb) (Du et al., 2013). Previous studies have shown that heavy metals like Mercury, Cadmium and Lead are capable of inducing wide range toxicity in the germinal epithelium as morphometric studies reveals low cell count, distorted lumen and cell death in general; other physiological changes includes male infertility, hepatotoxicity, bone marrow disorders and tumorgenesis (Xiao et al., 2012).
The primary mechanism of Pb toxicity has been found to be majorly induction of oxidative stress via inhibition of allosteric sites in metalloenzymes (Alkaline Phosphatase and Cytochrome C Oxidase:: CcOX). Several cell death pathways will be activated following lead toxicity (Kumari et al., 2013). Firstly is the mitochondria pathway which involves inhibition of the Heme a3-Cuα binuclear centre of CcOX; an enzyme concerned with conversion of molecular oxygen into water at the complex V of the Electron Transport Chain (ETC). When such inhibition occurs, accumulation of oxygen and electrons in the mitochondria matrix is resultant (Musatov and Robinson, 2012), thus molecular oxygen reacts with electrons from reduction of food substances to generate Reactive Oxygen Species (ROS); also called superoxide anions or oxygen radicals. The primary role of ROS is peroxidation of lipids (Humphrey et al., 2012). The first response involves production of mtNOS (mitochondria nitric oxide synthase to counter the Nitric Oxide (NO) that are formed from the reaction of ROS with other nitrogen containing groups (Ekici et al., 2012). This NO pathway represents the central apoptotic pathway resulting from oxidative stress as the NO thus formed is a naturally occurring endogenous modulator of cellular activities but when produced in large quantities can trigger apoptosis by inducing a wide range nuclear damage (Dong et al., 2012). The second pathway is a cytoplasmic pathway where the ROS thus formed induces lipid peroxidation and leaks into the cytoplasm. This is vast and rapid as the lipids constitutes 70% of biomembranes including the nuclear, mitochondria, lysosomal and cell membrane. Such peroxidation will inhibit Alkaline Phosphatase and affect the osmotic balance of the cell (Cramer et al., 2011). This is usually characterized by cell enlargement or rupture and presence of secondary autophagic bodies resulting from sites of lysosomal rupture and hydrolytic digestion within the cell.
Since the germinal epithelium contains several dividing cells, the cell cycle is most active around this region. This study evaluates the role of Pb as an agent capable of inducing degeneration via cell cycle proteins as well as the role of Selenium and Zinc (2.25 mg kg-1) as agents capable of reducing such toxicity by evaluation of cell proliferation. Degeneration and toxicity can be accessed through the mitochondria and cytoplasmic pathways by measuring immunohistochemically the level of expression of p53 (a 53KDa tumor suppressor protein) that is usually activated the Gap phases of the cell cycle to proof read the genome for errors such as cleavage and deletions which are induced by Pb toxicity (National Toxicology Program, 2007). The p53 is a nucleolus that will digest the genetic materials in case of such errors (Mateo et al., 1995). The mitochondria pathways can be tracked via the caspase -3 and -9 systems through a p21 shunt to Cathepsin D (CAD) (Zeng et al., 2009). Although the expression of Cathepsin D has been found in most cases to be inversely related to the expression of p53 its exact role in p53 regulated cell cycle is unclear. CAD is a protein that functions to regulate senescence (aging which might occur due to stress); Bax is also present in this inner cycle but shows a much closer relation to p53 than CAD (Saewu et al., 2012). Selenium and zinc are trace elements in the human body and will usually function as co-enzymes and co-factors for several functional groups most of which are enzymes and vitamins. Some of such vitamins include Vitamin C a potent radical scavenger and Alkaline Phosphatase an enzyme which increases viability of membrane transport. It is also possible that apart from radical scavenging selenium and zinc competes for the binding sites of lead but does not induce oxidative stress by retaining the structure of the enzyme allosteric sites. This form of treatment can be described as targeted and competitive as heavy metals will usually adopt a similar pathway for Bioactivation.
MATERIALS AND METHODS
Sixty F1 generation adult Wistar rats were used. The animals were procured and kept in the animal holding facility of the Osun State University and allowed to acclimatize. The animals were then divided into 4 groups of 15 animals each. The animal handling protocol followed the Helsinki Convention on animals use for research. Group 1 (control) received normal saline, group 2 received 100 mg kg-1 b.wt. of lead acetate (Pb), group 3 received 100 mg kg-1 of Pb, 2.25 mg kg-1 of Se and 2.25 mg kg-1 of Zn. The duration of treatment was 56 days for all groups. The animals were sacrificed on the 57th day and the testes were dissected to obtain the seminiferous tubule. The tissues were immediately fixed in Bouin’s fluid for 24 h and a change of Bouin’s fluid for another 24 h. Tissue processing was done to obtain paraffin wax embedded sections using the method of Jeong et al. (2007) (Se: Sodium Selenium and Zn: Chelated Zinc).
Histology: The embedded tissues were sectioned to obtain 7 μm thick sections for routine histological staining in Hematoxylin and Eosin (H and E) using the method of Ma et al. (2012).
Immunohistochemistry: The paraffin wax embedded sections were mounted on a glass slide in preparation for antigen retrieval where the slides were immersed in urea overnight and then placed in a microwave for 45 min to re-activate the antigens and proteins in the tissue sections. Primary antibody treatment involved treating the sections with biotinylated goat serum for one hour following which the sections were transferred to 1% Bovine Serum Albumin (BSA) to block non-specific protein reactions. Secondary treatment involved the use of diluted anti-CAD, Anti-p53 and Anti-Ki-67 on the pre-treated sections for one hour. The immunopositive reactions were developed using a polymer 3’3’ Diaminobenzidine Tetrachloride (DAB) with colour intensification involving the use of mathenamine silver kit. The sections were counterstained in hematoxylin and treated in 1% acid alcohol (freshly prepared).
RESULTS AND DISCUSSION
The cytoplasmic pathway was observed to be most predominant in lead toxicity. Although, oxidative stress is the primary, it is important to distinguish the resultant forms of cell death in the germinal epithelium. The apoptotic pathway was predominant as it seen in the lead treated group 2 (Fig. 1b, 2b, 3b and 4b) although necrosis was observed in the Sertoli cells closer to the basement membrane. The changes are not only cellular but in some cases membrane based (BM). The thickness of the membrane reduced in the treatment group and in some cases they are void of connective tissue. The expression of p53 was found to be prominent in the basement membrane region. This suggests alteration in membrane integrity and in essence structural conformation of the BM and the junctional complexes that forms the basis of the barrier. Although, toxicity was observed in the group treated with Pb then Selenium and Zinc (Se+Zn), it was observed that the damage was greatly reduced. The observed cellular changes were limited to slight enlargement in cell size with no obvious cytoarchitectural alteration thus confirming that at moderate doses Se+Zn can reduce the toxicity of lead either by functioning as co-factors to activate radical scavengers or by functioning as a competition to reduce the ability of Pb to alter the allosteric sites in oxidative enzymes (Fig. 1c, 2c, 3c and 4c). A second control was set up to confirm the proliferative effect of Se+Zn trace on normal germinal epithelium without toxicity, it was observed that the Se+Zn induced cell proliferation (‡ in Fig. 1d) and Ki-67 immunohistochemistry in Fig. 4d) similar to the findings of Chen et al. (2013). Cathepsin D and p53 expression were also greatly reduced showing that neither the cytoplasmic or mitochondria pathway was activated nor oxidative stress was not induced and if at all induced was not significant enough to cause any obvious structural change. The proliferative effect of Se+Zn was also further confirmed on comparison of the control with group 4 (Se+Zn) in Ki-67 studies where the Se+Zn treated (Fig. 4d) was more immunopositive for the protein that the control (Fig. 4a).
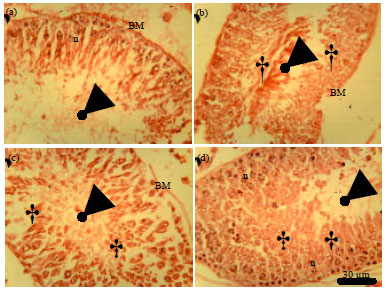 | |
| Fig. 1(a-d): | General morphology of the germinal epithelium of wild type adult wistar rats (stained with Hematoxylin and Eosin) (a) Control, (b) 100 mg kg-1 Pb-Acetate, (c) Pb-Acetate+Se+Zn and (D) Se+Zn only, Degeneration was most prominent in the lead treatment group, cell proliferation is most prominent in the Se+Zn treatment group. (‡) represents regions of cell proliferation and (†) represents regions of cellular degeneration, (n) represents normal cells of the epithelium, (BM) basement membrane, arrow head indicates the lumen of the seminiferous tubule (Scale bar is 30 μm) |
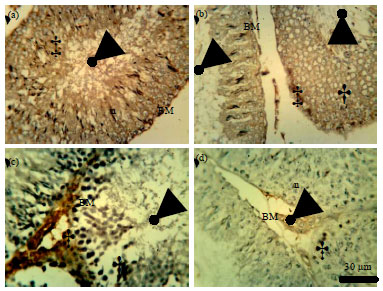 | |
| Fig. 2(a-d): | p53 Immunohistochemistry (for cell cycle dysregulation) in the germinal epithelium of wild type adult Wistar rats, (a) Control, (b) 100 mg kg 1 Pb-Acetate, (c) Pb-Acetate+Se+Zn and (d) Se+Zn only, The Pb treatment groups showed reduced p53 activity (2B) compared with the control (2A). The basement membrane (rich in spermatogonia and stem cell-lines) showed increased p53 activity in the Pb+Se+Zn (2C) and the Se+Zn treatment groups (2D), (†) represents regions of Ki-67 immunopositivity and (†) represents regions of cellular degeneration, (n) represents normal cells of the epithelium, (BM) basement membrane, arrow head indicates the lumen of the seminiferous tubule (Scale bar is 30 μm) |
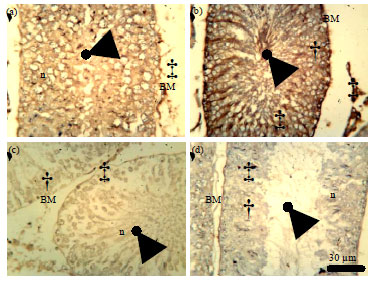 | |
| Fig. 3(a-d): | Cathepsin D (CAD) immunohistochemistry in the germinal epithelium of wild type adult Wistar rats, (a) Control, (b) 100 mg kg 1 Pb-Acetate, (c) Pb-Acetate+Se+Zn and (D) Se+Zn only. CAD activity is high in the BM of the control (3a), in the BM and degenerating cells of the lead treated group (3b), in the BM of Pb+Se+Zn and it is widely diffused in the epithelial cells of the Se+Zn treated group (3d). (‡) represents regions of CAD immunopositivity and (†) represents regions of cellular degeneration, (n) represents normal cells of the epithelium, (BM) basement membrane, arrow head indicates the lumen of the seminiferous tubule (Scale bar is 30 μm) |
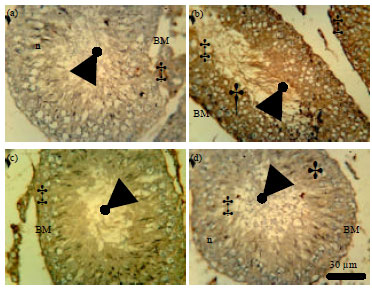 | |
| Fig. 4(a-d): | Ki-67 Immunohistochemistry (for cell proliferation) in the germinal epithelium of wild type adult Wistar rats, (a) Control, (b) 100 mg kg-1 Pb-Acetate, (c) Pb-Acetate+Se+Zn and (d) Se+Zn only. Cell proliferation marked by Ki-67 immunopositivity is higher in the Se+Zn treatment group (Fig. 4d), followed by the control (4a). (‡) represents regions of Ki-67 immunopositivity and (†) represents regions of cellular degeneration, (n) represents normal cells of the epithelium, (BM) basement membrane, arrow head indicates the lumen of the seminiferous tubule (Scale bar is 30 μm) |
Lorenzetti et al. (2012) showed that marine compound can increase sperm quality, count and vialibility via proliferation. To further buttress the role of Se+Zn trace in improving sperm quality and reducing Pb toxicity via competition and scavenging, the studies of Ayinde et al. (2012) showed that intensive vitamin C and E treatment greatly reversed the toxicity of Pb in the germinal epithelium. Weight gain was recorded and the sperm motility/count was greatly improved. The studies of Priya and Reddy, 2012 confirmed the oxidative stress pathway associated with Pb toxicity in the cells of the germinal epithelium although the mechanism described there in described cell death as cell death rather than in various pathways leading to apoptosis and necrosis. The group 4, treated with Se+Zn only showed a significant increase in body weights but testicular weights were unaffected (not shown). Histological evaluation of the rat testes revealed that rats treated with lead (Pb) only showed interstitial oedema, few spermatogoonia close to the basement membrane and an abnormal shape of Spermatids compared to the normal spermatogenic phenotypes observed in the lead, selenium and zinc treated group and the selenium and zinc only treated groups.
Recent studies have also shown that selenium can be used in combating BPH in prostate cancer patients (Minutoli et al., 2013), proliferative and anti oxidants effects of selenium has also been described by Zeng et al. (2013). If included in nutrients it can activate antioxidant vitamins and improve activity of radical scavenger. Its ability to improve proliferation in the epithelium was similar to the discovery of Zeng et al. (2013) where selenium was described as an essential trace element for humans and animals and several findings suggest that dietary Se intake may be necessary for bone health. Such findings may relate to roles of Se in antioxidant protection, enhanced immune surveillance and modulation of cell proliferation. Elucidation of the mechanisms by which Se supports these cellular processes can lead to a better understanding of the role of this nutrient in normal bone metabolism (Zeng et al., 2013). The selenium supplement may also improve endothelial protein efficiency and enhance junctional complex activities to further combat Pb toxicity. Da Silva et al. (2012) showed that zinc can improve regeneration of liver cells by inhibiting SOCS3 expression to enhance IL-6/STAT3 regenerative and proliferative pathways (this study addresses the Ki-67 proliferative pathway). Zinc have also been described as a primary requirement for cell growth and cell proliferation (Tynga et al., 2012; Jones et al., 2013) as it is a requirement by several growth enzymes.
CONCLUSION
Pb induced toxicity can follow a mitochondria pathway (CAD) or a cytoplasmic pathway involving p53, the most predominant form of cell death is apoptosis which can result from both pathways. Se+Zn treatment improves proliferation and counters Pb toxicity by substitution, activation of enzymes and growth factors, activation of endothelial factors and activation of radical scavengers.
ACKNOWLEDGMENT
We acknowledge the contributions of Jonathan Madukwe of the Histopathology Department, National Hospital Abuja, Also the Directorate of Research, Afe Babalola University, Ado-Ekiti, Nigeria.
REFERENCES
- Ayinde, O.C., S. Ogunnowo and R.A. Ogedegbe, 2012. Influence of vitamin C and vitamin E on testicular zinc content and testicular toxicity in lead exposed albino rats. BMC Pharmacol. Toxicol., Vol. 13.
CrossRefDirect Link - Chen, L., X. Zhang, W. Zhou, Q. Qiao and H. Liang et al., 2013. The interactive effects of cytoskeleton disruption and mitochondria dysfunction lead to reproductive toxicity induced by microcystin-LR. PLoS ONE, Vol. 8.
CrossRef - Cramer, W.A., S.D. Zakharov, S. Saif Hasan, H. Zhang and D. Baniulis et al., 2011. Membrane proteins in four acts: Function precedes structure determination. Methods, 55: 415-420.
CrossRef - Da Silva, C.G., P. Studer, M. Skroch, J. Mahiou and D.C. Minussi et al., 2012. A20 promotes liver regeneration by decreasing SOCS3 expression to enhance IL-6/STAT3 proliferative signals. Hepatology, (In Press).
CrossRef - Dong, Y., W. Zhang, B. Lai, W.J. Luan, Y.H. Zhu, B.Q. Zhao and P. Zheng, 2012. Two free radical pathways mediate chemical hypoxia-induced glutamate release in synaptosomes from the prefrontal cortex. Biochim. Biophys. Acta, 1823: 493-504.
CrossRef - Du, M., J. Young, M. De Asis, J. Cipollone and C. Roskelley et al., 2013. A novel subcellular machine contributes to basal junction remodeling in the seminiferous epithelium. Biol. Reprod., (In Press).
CrossRefDirect Link - Ekici, S., H. Yang, H.G. Koch and F. Daldal, 2012. Novel transporter required for biogenesis of cbb3-type cytochrome c oxidase in Rhodobacter capsulatus. mBio, Vol. No. 1.
CrossRefDirect Link - Humphrey, D.M, R.B. Parsons, Z.N. Ludlow, T. Riemensperger and G. Esposito et al., 2012. Alternative oxidase rescues mitochondria-mediated dopaminergic cell loss in Drosophila. Hum. Mol. Genet., 21: 2698-2712.
CrossRefDirect Link - Jeong, C.W., H. Kim, S. Kim, S.H. Kim, C. Moon and T. Shin, 2007. Immunohistochemical study of flotillin-1 in rat testis with ischemia/reperfusion injury. Cell Biol. Int., 31: 609-614.
CrossRef - Jones, C., S. St-Jean, I. Frechette, D. Bergeron, N. Rivard and F. Boudreau, 2013. Identification of a novel promyelocytic leukemia zinc-finger isoform required for colorectal cancer cell growth and survival. Int. J. Cancer, (In Press).
CrossRef - Ma, K., R. Babij, E. Cortes, J.P.G. Vonsattel and E.D. Louis, 2012. Cerebellar pathology of a dual clinical diagnosis: Patients with essential tremor and dystonia. Tremor Other Hyperkinetic Movements, Vol. 2.
Direct Link - Kumari, U., W. Ya Jun, B. Huat Bay and A. Lyakhovich, 2013. Evidence of mitochondrial dysfunction and impaired ROS detoxifying machinery in fanconi anemia cells. Oncogene, (In Press).
CrossRefDirect Link - Lorenzetti, A., F. Marotta, H. Yadav, G. Celep and E. Minelli et al., 2012. Improving sperm quality and spermatogenesis through a bioactive marine compound: An experimental study. Acta Biomed., 83: 108-113.
PubMedDirect Link - Mateo, M.S., M. Sanchez-Beato, J.C. Martinez, A. Orfao, J.L. Orradre and M.A. Piris, 1995. p53, Rb and bcl-2 expression during the cell cycle: A study in phytohaemagglutinin stimulated lymphocytes and microwave irradiated lymphoid tissue sections. J. Clin. Pathol., 48: 151-159.
CrossRef - Minutoli, L., A. Bitto, F. Squadrito, H. Marini and N. Irrera et al., 2013. Serenoa repens, lycopene and selenium: A triple therapeutic approach to manage benign prostatic hyperplasia. Curr. Med. Chem., (In Press).
Direct Link - Mok, K.W., P.P. Lie, D.D. Mruk, J. Mannu, P.P. Mathur, B. Silvestrini and C.Y. Cheng, 2012. The apical ectoplasmic specialization-blood-testis barrier functional axis is a novel target for male contraception. Adv. Exp. Med. Biol., 763: 334-355.
PubMedDirect Link - Musatov, A. and N.C. Robinson, 2012. Susceptibility of mitochondrial electron-transport complexes to oxidative damage. Focus on cytochrome c oxidase. Free Radical Res., 46: 1313-1326.
CrossRef - National Toxicology Program, 2007. Toxicology studies of sodium bromate (CAS No. 7789-38-0) in genetically modified (FVB Tg.AC hemizygous) mice (dermal and drinking water studies) and carcinogenicity studies of sodium bromate in genetically modified [B6.129-Trp53tm1Brd (N5) haploinsufficient] mice (drinking water studies). Natl. Toxicol. Program Genet. Modif. Model Rep., 6: 1-169.
PubMedDirect Link - Priya, P.H. and P.S. Reddy, 2012. Effect of restraint stress on lead-induced male reproductive toxicity in rats. J. Exp. Zool. Part A: Ecol. Genet. Physiol., 317: 455-465.
CrossRefDirect Link - Saewu, A., S. Asuvapongpatana, C. Chotwiwatthanakun, A. Tantiwongse, W. Weerachatyanukul and S. Thitilertdecha, 2012. Cathepsin D in human reproductive tissues: Cellular localization in testis and epididymis and surface distribution in different sperm conditions. J. Androl., 33: 726-734.
PubMed - Su, W. and X. Liu, 2013. Rab13 regulates sertoli cell permeability barrier dynamics through protein kinase A. J. Mol. Endocrinol., (In Press).
CrossRefDirect Link - Tajaddini, S., S. Ebrahimi, B. Behnam, M. Bakhtiyari and M.T. Joghataei et al., 2013. Antioxidant effect of manganese on the testis structure and sperm parameters of formalin-treated mice. Andrologia, (In Press).
CrossRef - Tynga, I.M., N.N. Houreld and H. Abrahamse, 2012. The primary subcellular localization of Zinc phthalocyanine and its cellular impact on viability, proliferation and structure of breast cancer cells (MCF-7). J. Photochem. Photobiol. B: Biol., (In Press).
CrossRef - Xiang, Y., P. Luo, Y. Cao and Z.W. Yang, 2013. Long-term effect of vasectomy on spermatogenesis in men: A morphometric study. Asian J. Androl., (In Press).
CrossRef - Xiao, X., D.D. Mruk, F.L. Cheng and C.Y. Cheng, 2012. C-Src and c-Yes are two unlikely partners of spermatogenesis and their roles in blood-testis barrier dynamics. Adv. Exp. Med. Biol., 763: 295-317.
PubMedDirect Link - Zeng, H., J.J. Cao and G.F. Combs Jr., 2013. Selenium in bone health: roles in antioxidant protection and cell proliferation. Nutrients, 5: 97-110.
CrossRef - Zeng, J., J. Racicott and C.R. Morales, 2009. The inactivation of the sortilin gene leads to a partial disruption of prosaposin trafficking to the lysosomes. Exp. Cell Res., 315: 3112-3124.
CrossRef