
A. El-Sayed
Laboratory of Molecular Epidemiology, Faculty of Veterinary Medicine, Cairo University, Egypt
A. Prince
Agricultural Research Service, USDA, 10300 Baltimore Avenue, Beltsville, Maryland 20705, United States
A. Fawzy
Laboratory of Molecular Epidemiology, Faculty of Veterinary Medicine, Cairo University, Egypt
Nadra-Elwgoud
Department of Medicine and Infectious Diseases, Faculty of Veterinary Medicine, Cairo University, Egypt
M.I. Abdou
Department of Medicine and Infectious Diseases, Faculty of Veterinary Medicine, Cairo University, Egypt
L. Omar
Central Laboratories for Evaluation of Veterinary Biologics, Abbasia, Cairo, Egypt
A. Fayed
Laboratory of Molecular Epidemiology, Faculty of Veterinary Medicine, Cairo University, Egypt
M. Salem
Laboratory of Molecular Epidemiology, Faculty of Veterinary Medicine, Cairo University, Egypt
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 11 | Page No.: 524-529







ABSTRACT
In opposite to most countries, avian influenza virus H5N1 became endemic in Egypt. Since, its first emerge in 2006 in Egypt, the virus could infect different species of birds and animals and even human. Beside the great economic losses to the local poultry industry in Egypt, the virus infected 166 confirmed human cases, 59 cases ended fatally. In the present study, the persistence of the avian influenza in the Egyptian environment was studied. For this purpose, serum samples were collected from human, cattle, buffaloes, sheep, goat, horses, donkeys, swine, sewage rats, stray dogs and stray cats. The sera were collected from Cairo and the surrounding governorates to be examined for the presence of anti-H5N1 antibodies by Haemagglutination Inhibition Test (HI) and ELISA test. Clear differences in the seroprevalence were noticed among different species and also between the results obtained by both techniques indicating the difference in test accuracy. The present data indicate wide spread of the H5N1 virus in the Egyptian environment.
PDF Abstract XML References Citation
How to cite this article
A. El-Sayed, A. Prince, A. Fawzy, Nadra-Elwgoud, M.I. Abdou, L. Omar, A. Fayed and M. Salem, 2013. Sero-prevalence of Avian Influenza in Animals and Human in Egypt. Pakistan Journal of Biological Sciences, 16: 524-529.
DOI: 10.3923/pjbs.2013.524.529
URL: https://scialert.net/abstract/?doi=pjbs.2013.524.529
DOI: 10.3923/pjbs.2013.524.529
URL: https://scialert.net/abstract/?doi=pjbs.2013.524.529
INTRODUCTION
In the first few years after its introduction to Egypt in 2006, the Highly Pathogenic Avian Influenza (HPAI) H5N1 outbreaks had a seasonal incidence and were usually accompanied with the season of bird migration (Aly et al., 2008). Later on, the diseases became endemic in Egypt, as well as in other five countries worldwide (FAO-OIE-WHO, 2011). The viral persistence in Egypt may be attributed to many factors including the early application of vaccination directly after the introduction of the virus to Egypt. The used vaccines were imported from different countries and offered, in general, a low protection level. In addition, this massive vaccination policy enhanced the evolution of genetic drift evolution under vaccination pressure (Abdel-Moneim et al., 2011a). Other additional factors enhanced the persistence of the virus include, the unorganized rural poultry production in backyards and their marketing as well as fast and randomized movement of poultry, by-products and manure. Moreover, insufficient human awareness and the unhygienic disposal of dead birds and the use of untreated wastes of poultry farms to feed farmed fish. The contaminated materials will be, in turn, fed by wild and aquatic birds and allow the virus to persist in the community (Aly et al., 2008; El-Sayed et al., 2010a). The persistence of the H5N1 infections in Egypt represents a great threat for poultry industry, Egyptian economy and even for the international public health. Local control programs of the disease started directly after the disease emerge through intensive vaccination campaign. Yet the importation of foreign vaccines, the co-existence of human and other avian flu viruses and the continuous introduction of new viruses with migratory birds led to the enrichment of the genetic pool of the virus in Egypt (Beato et al., 2013). All these factors, in addition to the segmented nature of the genome of the influenza virus, facilitated the genetic re-assortment and the continuous evolution of new viruses (Arafa et al., 2012), such as the highly virulent Gharbia isolate with case fatality 100% among humans (Recombinomics, 2006), or Tamiflu resistant isolates (Kage, 2007). The antigenic interaction between H5N1 and the other human Influenza viruses (H1 and H3) in Egypt was reported (Cattoli et al., 2011).
Although it is known that the virus can induce interspecies infections (Marschall and Hartmann, 2008), limited studies were carried out to investigate the H5N1 situation among other animals rather than birds such as dogs (Songserm et al., 2006; Witthawat et al., 2009), cats (Klopfleisch et al., 2007) and wild mammals (Rimmelzwaan et al., 2006).
Recently and after the warning of the aggressive comeback of avian influenza by the FAO and other newly emerged subtypes as the H9N2 (FAO, 2011; El-Zoghby et al., 2012), deeper studies about reservoir and epidemiology of H5N1 are urgently required.
The present study aims to detect the sero-prevalence of H5N1 in Egyptian human population, ruminants (cattle, buffaloes, sheep and goat), swine, working animals (horses-donkeys), rodents (rats) and stray pets (dogs and cats) using highly specific ELISA system. In parallel, the work aimed to compare the accuracy of ELISA against the routinely applied Haemagglutination Inhibition (HI) test. To achieve these purposes, serum samples from the previously mentioned species were tested by ELISA and HI for the presence of anti-H5N1 antibodies.
MATERIALS AND METHODS
Sampling: Sera from human (n = 299), cattle (n = 50), buffaloes (n = 50), sheep (n = 50), goat (n = 50), horses (n = 150), donkeys (n = 36), swine (n = 93), rats (n = 72), stray pets (25 dogs, 25 cats) were collected from hotspots in Nile-Delta. Positive controls were obtained from a confirmed human patient and from experimentally vaccinated animals.
All sera were first treated to adsorb and inactivate the nonspecific agglutinins which might interfere with the reaction specificity as recommended by OIE (2005) and Rowe et al. (1999). The positive control sera were obtained from a human case with a proven H5N1 infection and from animals (2 from every tested species) which were vaccinated with a local killed H5N1 vaccine. The vaccinated animals received an additional booster dose after 2 weeks of their first inoculation. One week later, the serum samples were collected and the two sera from the same species were then pooled. The used vaccine was prepared from a native H5N1 field isolate by the Central Laboratories for Evaluation of Veterinary Biologics (CLEVB) and sold for use in poultry farms. The animals received 10 times the recommended dose for poultry.
Hemagglutination-inhibition test: The test was performed in the CLEVB according to the previously published protocols (OIE, 2005; El-Sayed et al., 2010b) using inactivated H5N1 antigen prepared from a local H5N1 Egyptian field isolates (Chicken/Egypt/9402-NAMRU3-CLEVB213/2007) on allantoic fluid of ECE according to their standard protocol. All collected sera were frozen at -20°C until being examined.
Following the serum primary treatment, the titers of the anti-avian influenza antibodies were determined by the use of hemagglutination inhibition test (HI) with 0.5% CRBC. Briefly, 25 μL of the diluted inactivated serum were mixed with an equal volume of the buffer in V-bottomed microtitre plates (Nunc, Langenselbold, Germany). Then, a double fold serial dilution of the sera was performed. This was followed by the addition of the influenza virus antigen (H5N1) containing 4 hemagglutinating units. After one hour reaction time at room temperature, 0.5% RBCs solution was added and the final result was read after 60 min. For the twofold dilution of the tested sera, Veronal buffer pH 7,2 (BIOMERIEUX, France) was used.
ELISA: As only avian specific H5N1 ELISA systems are available in Egyptian market, a protein G modified H5N1-ELISA was applied. Using The ID Screen® Influenza H5 Competition ELISA kit which detects antibodies against the Haemagglutinin H5 of the Influenza A virus in bird sera (ID VET, France). As the Protein G binds with some but not all antibodies of different species, the ELISA could be performed with sera obtained from human, cattle, buffaloes, sheep, goat, swine and horses.
Shortly, 80 μL of dilution buffer II were poured in each well, 20 μL of four HI negative samples in A1, A2, A3, A4 wells. Add 20 μL of the positive control in A5. This was followed by the addition of 20 μL of each sample to be tested to the remaining wells. After incubation for 1 h+5 min at 37°C (+2°C), the plates were washed 3 times with approximately 300 μL of the wash solution (wash solution 1x was prepared by diluting wash concentrate 20x in distilled water). Then 50 μL of the labeled protein G was added to each well. The plates were then re-incubated for 30 min +2 min at 21°C (+5°C). After that, the wells were emptied and Washed 3 times with approximately 300 μL of the wash solution. The substrate solution (50 μL) was added to each well. The plate was Incubated 10 +1 min at 21°C (+5°C) in dark. Finally, 50 μL of the stop solution was added to each well in order to stop the reaction before measuring the absorbance at 450 nm.
RESULTS
In the present work, different serum samples from different species were obtained and tested for the presence of anti-avian Influenza antibodies, including human. Anti-H5N1 could be detected in variable degrees in all investigated species with the exception of ruminants. The obtained data are summarized in Table 1. The prevalence of avian influenza in human samples is 8.7% by using HI test and 14% by ELISA. The sero-prevalence shows that the females are clearly more exposed than males (6.5% of males and 13.9% of females) in investigated samples originating from the Nile Delta (Cairo and the surrounding country side) for avian influenza. Meanwhile, 12% of Cairo residents and 8.4% of the farmers were sero-positive (Table 2).
| Table 1: | Results obtained by the use of HI and ELISA for the detection of Anti-H5N1 antibodies |
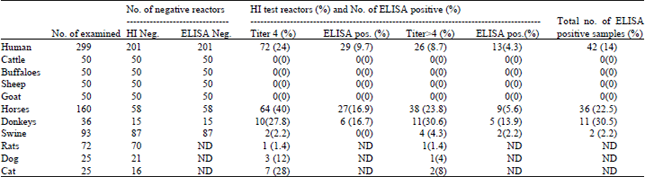 | |
| ND: Not done as no control positive serum was available or due to the poor binding affinity to protein, G: Information needed concerning the binding affinity of the different sera to protein G was derived from the Biosynthesis company available at: http://www.biosyn.com/tew.aspx?qid = 230 | |
| Table 2: | The listed data compare the percentage of sero-prevalence in the examined human sera derived from different geographic locations and according to their sex, the involved sera in this table are the sera with titer >4 by the HI test |
 | |
| In equines, the sero-prevalence of H5N1 ranged from 22.55% (ELISA) to 23.75% (HI) while in pigs and rats the prevalence of H5N1 was very low. Among dogs and cats the sero-prevalence of H5N1 is 4 and 8%, respectively (Table 1). | |
DISCUSSION
Since the first isolation of the avian influenza virus H5N1 it continued to evolve and to alter receptor specificity to enhance its binding affinity to the new host species (Watanabe et al., 2012). Therefore, the investigation of possible hidden or new sources of the virus is necessary for the performance of efficient control programs in Egypt. Although, the H5N1 could be isolated from dead mammals of different species, yet such sporadic reports did not offer a clear answer about the sero-prevalence of the disease in affected species. On the other hand and although the HI Test is the standard test for the diagnosis of avian influenza infection, different protocols are published for primary treatment of the sera before the performance of the test. The primary serum treatment aims to eliminate nonspecific reactors.
For the application of ELISA, the adaptation of commercial ELISA kits to other species rather than poultry required the use of Protein G in an extra step of the protocol. This enabled the team to use the adapted system performed with sera obtained from Human, Cattle, Buffaloes, Sheep, Goat, Swine and Horses only. Sera from other species (Rat, dog and cat) would not bind to protein G and therefore delivers false negative results.
As shown in Table 1, the lower sero-prevalence of avian influenza when using ELISA compared to HI in general, in parallel to the failure of ELISA to identify detectable antibodies in HI negative samples indicates the higher specificity of ELISA in comparison to the used HI protocol. However, this could also be attributed to the higher affinity of serum antibodies to the native antigens rather than the foreign ones (a local H5 antigen prepared from an Egyptian field isolate was used in HI against the use of imported ELISA plates which are coated with a foreign H5 antigen). This suggestion can by supported by previous findings reporting the antigenic interaction among local influenza viruses including the H5N1 in Egypt (Cattoli et al., 2011). This may indicate that some samples with borderline HI positive titer (Titer = 4) may include antibodies against other related influenza viruses rather than H5N1. However, it is also possible that this difference in specificity is attributed to nonspecific reactors which resisted the applied inactivation process and requires extra primary treatment as recommended by Lerdsamran et al. (2011) in opposite to the protocols applied in many laboratories.
Human Sera: In Egypt, where 166 humans were infected with avian influenza (59 deaths) according to the WHO reports, great attention must be given to the sero-prevalence of the disease among human population. Little literature could be found concerning the sero-prevalence of H5N1 in human. In Thailand, all examined human sera, obtained from highly exposed persons, were negative for H5N1 antibodies (Dejpichai et al., 2009) in opposite to the prevalence of the disease in Egyptians, where 8.7% (HI) and 14% (ELISA) were positive according to the present data. The high relationship between the field isolates of H5N1 and a mammalian adapted man made H5N1 strain (Didymus, 2011) increases the fear that silent circulation of the virus among human population may lead to the adaptation of the virus or its re-combination to threaten the global public health. The sero-prevalence in the country side shows that the females are clearly more exposed than males. The obtained data indicated that 6.5% of males and 13.9% of females in investigated samples originating from the Nile Delta (Cairo and the surrounding country side) were sero-positive for avian influenza. Meanwhile, 12% of Cairo residents and 8.4 % of the farmers were sero-positive (Table 2). This confirms previous findings and disagrees with that in South East Asia (El-Sayed et al., 2010a; Abdelwhab and Hafez, 2011). The high sero-prevalence in Cairo in general and in males especially is unexpected. This may be attributed to the overcrowdings and high density of the population in Cairo which favors the spread of influenza. In a country like Egypt, where the females works mainly inside their houses, the men are more exposed to the infection in overcrowded traffics as an example.
Ruminants: Few papers concerning the susceptibility of ruminants to influenza were published. However, the ability of cattle to develop antibodies when exposed to influenza virus was reported previously (H1, H3 and H5) (Kalthoff et al., 2008; Lin et al., 2010). Meanwhile, no data are available about the susceptibility of buffaloes, sheep and goats to infection. The obtained negative results indicate either the low susceptibility of ruminants to H5N1 infection in comparison to other farm animals or due to the low number of the examined ruminants (200 animals). However, possible infection of such species cannot be ruled out as the virus jumped to different new hosts.
Horses and donkeys: Although equine influenza is known to be caused by H7N7 and H3N8, the H5N1 could also be isolated from Egyptian horses and donkeys (Abdel-Moneim et al., 2010; Abdel-Moneim et al., 2011b). The epidemiological role of equines in the spread of the disease and the prevalence of H5N1 antibodies remains unclear. In the present work, no difference in disease susceptibility between both species can be detected. The seroprevalence of H5N1 ranged from 22.55% (ELISA) to 23.75% (HI) which may indicate an epidemiological role of equines in the disease persistence.
Pigs and rats: The isolation of H5N1 from rats and pigs was reported in 1998 (Shortridge et al., 1998). Later on, 4.6% of tested sera from Egyptian pigs with HI technique reacted positively to H5N1 (14). This ratio is clearly higher than that for pigs in China and Vietnam (Choi et al., 2005; Cyranoski, 2005). The present data indicate that this average was kept over the years with a little tendency to sink. The slight decrease in H5N1 sero-prevalnce in Egyptian swine may be related to the elimination of all swine reared in the Egyptian cities during the H1N1 pandemic. Only swine reared in Monasteries could survive. Such animals have less exposure risk. However, the present data confirm the negligible role of swine in H5N1 persistence.
Rats were collected from the sewage system of different governorates in north Egypt, the prevalence of H5N1 among rats was also negligible (1.4%). This might be attributed to the limited contact between rats and birds or due a relative species resistance against AI.
Dogs: Although the dogs are known to attract influenza (H3N8), yet they can also be victims for H5N1 infections (Maas et al., 2007). The seroprevalence of the disease ranged from 0% in Nigeria (Oluwayelu et al., 2011) and 15.9 or 25% in village dogs in Thailand (Butler, 2006; Witthawat et al., 2009), respectively. The dogs usually attract the infection after eating dead birds (Songserm et al., 2006). According to the represented results, the prevalence of H5 among Egyptian dogs is 4%. In the present study, dog samples were collected from the Capital Cairo, this may be the reason of being clearly lower than that in the highly exposed dogs reared in the villages of Thailand.
Cats: The outdoor cats in areas affected by HPAIV are at risk for lethal infection (Klopfleisch et al., 2007). Due to the close contact between cats and humans, the epidemiological role of cats in disease transmission needs more attention. The investigated cats here showed a sero-prevalence of 8% compared with 2,6% in Germany and Austria (Marschall et al., 2008) and 4,7% in Thailand (Witthawat et al., 2009). Affected cats may be either clinically or inapparently infected, Cats have been infected by direct contact with affected birds, especially by eating raw dead poultry. Experts fear that cats might give the virus an opportunity to adapt to mammals (Kuiken et al., 2006; Thiry et al., 2007; Marschall and Hartmann, 2008). The higher prevalence among Egyptian cats may be attributed to their nature as stray cats fed mainly on wastes as dead birds.
CONCLUSION
In conclusion, the present work reports the expansion of the host range of the avian influenza virus to infect some mammalian species. Some species were shown to be susceptible for attracting H5N1 infections such as Human, equines and cats; others are less susceptible as swine, rats and dogs. Meanwhile, none of the investigated ruminants was positive for AI. The use of the modified ELISA system delivers more sensitive results than those obtained by the routinely applied protocols of HI. Therefore, the use of Protein G dependent ELISA in the screening of avian influenza programs can be recommended according to the present data. Finally, the published situation of avian influenza in Egypt represents the tip of iceberg. An urgent and efficient control programs must be developed and applied.
ACKNOWLEDGMENT
This study is a part of a research project funded by Cairo University to investigate the hidden sources of avian influenza in the Egyptian environment.
REFERENCES
- Abdel-Moneim, A.S., A.E. Abdel-Ghany and S.A. Shany, 2010. Isolation and characterization of highly pathogenic avian influenza virus subtype H5N1 from donkeys. J. Biomed. Sci., Vol. 17.
CrossRef - Abdel-Moneim, A.S., M.A. Afifi and M.F. El-Kady, 2011. Genetic drift evolution under vaccination pressure among H5N1 Egyptian isolates. Virology, Vol. 8.
CrossRef - Abdelwhab, E.M. and H.M. Hafez, 2011. An overview of the epidemic of highly pathogenic H5N1 avian influenza virus in Egypt: Epidemiology and control challenges. Epidemiol. Infect., 139: 647-657.
CrossRefPubMedDirect Link - Aly, M.M., A. Arafa and M.K. Hassan, 2008. Epidemiological findings of outbreaks of disease caused by highly pathogenic H5N1 avian influenza virus in poultry in Egypt during 2006. Avian Dis., 52: 269-277.
CrossRefPubMedDirect Link - Beato, M.S., M. Mancin, J. Yang, A. Buratin and M. Ruffa et al., 2013. Antigenic characterization of recent H5N1 highly pathogenic avian influenza viruses circulating in Egyptian poultry. Virology, 435: 350-356.
CrossRef - Butler, D., 2006. Thai dogs carry bird-flu virus, but will they spread it? Nature, 439: 773-773.
CrossRef - Cattoli, G., M. Adelaide, N. Temperton, B. Zecchin and A. Buratin et al., 2011. Antigenic drift in H5N1 avian influenza virus in poultry is driven by mutations in major antigenic sites of the hemagglutinin molecule analogous to those for human influenza virus. J. Virol., 85: 8718-8724.
CrossRefDirect Link - Choi, Y.K., T.D. Nguyen, H. Ozaki, R.J. Webby and P. Puthavathana et al., 2005. Studies of H5N1 influenza virus infection of pigs by using viruses isolated in Vietnam and Thailand in 2004. J. Virol., 79: 10821-10825.
CrossRefDirect Link - Dejpichai, R., Y. Laosiritaworn, P. Phuthavathana, T.M. Uyeki and M. O'Reilly et al., 2009. Seroprevalence of antibodies to avian influenza virus A (H5N1) among Residents of villages with human cases, Thailand, 2005. Emerg. Infect. Dis., Vol. 15.
Direct Link - El-Sayed, A., W. Awad, A. Fayed, H.P. Hamann and M. Zschock, 2010. Avian influenza prevalence in pigs, Egypt. Emerg. Infect. Dis., 16: 726-727.
PubMedDirect Link - El-Zoghby, E.F., A.S. Arafa, M.K. Hassan, M.M. Aly and A. Selim et al., 2012. Isolation of H9N2 avian influenza virus from bobwhite quail (Colinus virginianus) in Egypt. Arch. Virol., 157: 1167-1172.
CrossRefPubMedDirect Link - Klopfleisch, R., P.U. Wolf, W. Uhl, S. Gerst and T. Harder et al., 2007. Distribution of lesions and antigen of highly pathogenic avian influenza virus A/Swan/Germany/R65/06 (H5N1) in domestic cats after presumptive infection by wild birds. Vet. Pathol., 44: 261-268.
PubMedDirect Link - Kuiken, T.H., R. Fouchier, G. Rimmelzwaan, A. Osterhaus and P. Roeder, 2006. Feline friend or potential foe? Nature, 440: 741-742.
CrossRefDirect Link - Lerdsamran, H., C. Pittayawonganon, P. Pooruk, A. Mungaomklang and S. Iamsirithaworn et al., 2011. Serological response to the 2009 pandemic influenza A (H1N1) virus for disease diagnosis and estimating the infection rate in Thai population. PLoS ONE, Vol. 6.
CrossRef - Maas, R., M. Tacken, L. Ruuls, G. Koch, E. van Rooij and N. Stockhofe-Zurwieden, 2007. Avian influenza (H5N1) susceptibility and receptors in dogs. Emerg. Infect. Dis., 13: 1219-1221.
PubMedDirect Link - Marschall, J., B. Schulz, T.C.H. Priv-Doz, T.W.V. Priv-Doz, J. Huebner, E. Huisinga and K. Hartmann, 2008. Prevalence of influenza A H5N1 virus in cats from areas with occurrence of highly pathogenic avian influenza in birds. J. Feline Med. Surg., 10: 355-358.
PubMed - Oluwayelu, D.O., C.O. Aiki-Raji, N. Che and O. Ahmadu, 2011. Prevalence of avian origin H5 and H7 influenza virus antibodies in dogs in Ibadan and Sagamu, Southwestern Nigeria. Afr. J. Biomed. Res., 14: 23-26.
Direct Link - Rimmelzwaan, G.F., D. van Riel, M. Baars, T.M. Bestebroer and G. van Amerongen et al., 2006. Influenza A virus (H5N1) infection in cats causes systemic disease with potential novel routes of virus spread within and between hosts. Am. J. Pathol., 168: 176-183.
PubMed - Rowe, T., R.A. Abernathy, J.H. Primmer, W.W. Thompson and X. Lu et al., 1999. Detection of antibody to avian influenza A (H5N1) virus in human serum by using a combination of serologic assays. J. Clin. Microbiol., 37: 937-943.
PubMedDirect Link - Shortridge, K.F., N.N. Zhou, Y. Guan, P. Gao and T. Ito et al., 1998. Characterization of avian H5N1 influenza viruses from poultry in Hong Kong. Virology, 252: 331-342.
Direct Link - Songserm, T., A. Amonsin, R. Jam-on, N. Sae-Heng and N. Pariyothorn et al., 2006. Fatal avian influenza A H5N1 in a dog. Emerg. Infect. Dis., 12: 1744-1747.
PubMedDirect Link - Witthawat, W., P. Weena, P. Kranapone, P. Pirom and C. Kridsda et al., 2009. Seroprevalence of HPAI H5N1 virus in domestic dogs and cats living in repeated outbreak areas in Thailand. Thai Ministry of Public Health. http://beid.ddc.moph.go.th/th/images/news/cheraton2122_10_52/poster/10WitthawatWiriyarat_poster.pdf.