
Elham Torbati
Microbiology Department, Islamic Azad University, North Tehran Branch, Tehran, Islamic Republic of Iran
Romina Karimzadeh Ghassab
Department of Clinical Biochemistry, Faculty of Medicine, Iran University of Medical Sciences, Tehran, Islamic Republic of Iran
Navid Dadashpour Davachi
Department of Animal Science, Faculty College of Agriculture and Natural Resources, University of Tehran, Karaj, Islamic Republic of Iran
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 24 | Page No.: 2041-2045







ABSTRACT
Hepatitis C virus (HCV) is an important cause of acute and chronic hepatitis which is a disorder with a high worldwide prevalence. HCV core protein was considered as immunogenic counterpart of the HCV vaccine and it is an ideal candidate for HCV vaccine. Since cytokines such as IL-6, TNF-α and IFN-Gamma are responsible for the prevention of viral infection, this study aimed to evaluate the effectiveness of HCV core protein as a vaccine. Ten BALB/c mice were immunized with HCV core protein and after 42 days the splenocytes were isolated and the IL-6 and INF-γ secretion were measured using ELISpot technique, at the same time TNF-α was determined by ELISA in the sera. The MTT assay was done to assess the viability of the cultured splenocytes. For evaluating the humoral immune response against the recombinant HCV core protein the DOT Blot test was used. Data was compared using one-way ANOVA test and significant results were considered at p<0.05. In the present study the IL-6, INF-γ and TNF-α levels were dramatically higher in the immunized mice compared to the control group (respectively, 22.9±1.26; 18.53±3.87; 53.96±4.54 and p<0.05). The immunized mice with recombinant HCV core protein showed higher amount of IL-6, INF-γ and TNF-α in the current study. Since the level of IL-6, TNF-α and IFN-γ is high in patients with acute HCV infection, thus a vaccine which could stimulate the secretion of these cytokines in advance may have a preventive role.
PDF Abstract XML References Citation
Received: March 16, 2013;
Accepted: April 05, 2013;
Published: September 16, 2013
How to cite this article
Elham Torbati, Romina Karimzadeh Ghassab and Navid Dadashpour Davachi, 2013. Recombinant HCV Core Protein and the Secretion of Associated Cytokines (IL-6, TNF-α and IFN-γ) in Immunized Mice. Pakistan Journal of Biological Sciences, 16: 2041-2045.
DOI: 10.3923/pjbs.2013.2041.2045
URL: https://scialert.net/abstract/?doi=pjbs.2013.2041.2045
DOI: 10.3923/pjbs.2013.2041.2045
URL: https://scialert.net/abstract/?doi=pjbs.2013.2041.2045
INTRODUCTION
Hepatitis C virus (HCV) is a main cause of both acute and chronic hepatitis which is a disease with a significant global impact. Based on WHO report, about 170 million individual (consisting 3% of world population) are the victims of HCV infection (Murata et al., 2003; Frank et al., 2010). However the prevalence of the HCV are different in various regions which is higher among Asians and Africans, on the other hand, humeral immune response, which plays the key role in protective immunity of various viral infections and it has not been observed that this immune response could have any actual role in viral clearance in patients with acute HCV infection or chronic hepatitis C treatment (Murata et al., 2003), to find a proper vaccine which could stimulate the Cellular responses specially cytokine secretions, could help to prevent or decrease the incidence of the infection among these populations.
One of the viral features is its acute reaction by the infected cell which included the activation of previous antiviral defense mechanism, commitment to apoptosis and cytokine secretion which leaded to decrease the level of viral replication and to control the viral spread (Mogensen and Paludan, 2010). Helper T cells (Th) are the main regulators that help to destruct the antigen and to reinforce the antibody (Najafizadeh et al., 2007). Th1 cytokines such as IL-2, IF-Gamma and TNF-alpha are essential for antiviral activities of immune system. The highest interleukin activity begins at the site of their production however, when entering the circulation they are biologically active, so the IL s could be measured from the target tissue and the Peripheral blood (Lapinski, 2001; Najafizadeh et al., 2007).
The HCV core protein is an extremely basic protein that constructs the viral nucleocapsid by binding to the RNA. The biochemical properties of HCV-core protein have not been well described. It is consisted of 191 aa and three distinctive domains (Boulant et al., 2005; Torbati et al., 2012). HCV core protein was considered as immunogenic counterpart of the HCV vaccine. Serum antibodies against HCV core protein epitopes have been detected in HCV patients (Torbati et al., 2012). According to the conservative feature of the core gene in different HCV genotypes and the low mutation incidence in this genomic region , make this protein an ideal candidate for HCV vaccine (Torbati et al., 2012).
The capability to monitor hepatitis C virus (HCV)-specific T cell immunity and cytokine secretion in immunized mice could provide new information about the privation of HCV infection. Elispot assay was used as well as the ELISA technique to evaluate a cross-section of HCV-core immunized mice based on our previous study (Torbati et al., 2012). To this purpose The amount of IL-6, TNF-α and IFN-γ in immunized mice with the recombinant protein were measured, these cytokines are responsible to prevent the viral infection.
MATERIALS AND METHODS
Recombinant protein: The confirmed recombinant protein from our previous study (Torbati et al., 2012) was used in order to evaluate the specified cytokines secretion.
Immunization of balb/c mice: As there is a great similarity between the balb/c mice and human immunity system mechanisms, identical (gender, age, weight, etc.,) mice were used in this study. 6-8 weeks old Female BALB/c mice (Razi Institute of Iran) were housed in approved animal care facilities during the experimental period. According to the Guiding Principles in the Care and Use of Laboratory Animals, the immunization performed as follow:
Control group: A group of 6 female balb/c mice was immunized with a total volume of 150 μL Phosphate-Buffered Salin-1X (PBS), divided in 3 footpad injections (50 μL per injection), with 2 weeks intervals.
Subject group: This group consisted of 10 balb/c mice, 50 ng of recombinant protein in complete Freund’s adjuvant (Sigma, Germany) were injected s.c in to the footpad on the first day, a booster with the same amount of protein in incomplete Freund’s adjuvant (Sigma, Germany) were injected 2 weeks later and on day 28, the mice were immunized with 50 ng of the protein in PBS. On day 42, before splenocytes isolation, the serum samples were collected by retro-orbital puncture.
Ex vivo direct enzyme-linked immunospot (ELISPOT) Assay: In order to assess the amount of T lymphocytes responsible for cytokine secretion, the splenocytes were collected using the available protocol in laboratory (BDB 2011) Fig. 1. ELISpot assay employs a capture antibody specific for Mouse, pre-coated onto a PVDF-backed microplate. Stimulated cells are directly pipetted into the wells and the immobilized antibody binds to the nearest vicinity of the cells secreting IL-6 or IFN-γ.
IFN-γ Elispot test: Elispot assay for detecting IFN-γ secreting cells specific for HCV core epitopes has been formerly described by Duenas-Carrera et al. Acosta-Rivero et al. (2009), PVDF-bottomed well plate (Millipore, Cat No. MAIPS45510) coated with 100 μL murine IFN-γ-specific antibody (Mouse IF gamma Elispot Ready-SET-Go, bioscience, CatNo.88-7384) the test carried out using the kit instructions. At the end, the plates were dried in dark room, the spots were counted using dissection microscope (Zisse, Germany).Specific spots were red round with a dark center and fairly fuzzy edges.
IL-6 Elispot test: The collected splenocytes, were then platted in to 96-well PVDF-backed coated with polyclonal antibody specific for mouse/rat IL-6 (R and D System, Cat No. EL406, USA), the procedure performed according to the kit instructions (Mouse/Rat IL-6 ELISpot Kit, Cat No. EL406, USA), after the final wash, the developed microplate analyzed, using dissection microscope. Specific spots are blue round with a dark center and slightly fuzzy edges.
TNF-α Enzyme-linked Immunosorbent assay: The sera of immunized mice were collected before scarification. Pre-immune sera were considered as negative controls. The test was performed based on the kit instruction (Mouse TNF-α Immunoassay Elisa kit, R and D System, USA) and the optical density was measured at 640 nm using the microplate reader (Sunrise Tecan, Austria).
 | |
| Fig. 1: | The collected splenocytes of immunized mice |
MTT test: In order to assess the viability of the cultured splenocytes as a complementary test, cells (5x103 cells well-1) were plated onto 96-well plates and cultured for 3 days (every 24 h the cells were transferred to a new culture), then 100 μL of MTT (Sigma, USA) (5 mg mL-1 MTT ,90 μL RPMI) added to each well and were incubated at 37°C for 4 h, following centrifuged at 1000 g for 5 min. The supernatant emptied and 200 μL of Dimethyl sulfoxide (DMSO) (Sigma, Germany) was added to each well. The stained cells were eluted with methanol (Merck, Germany) and the optical density was measured at 590 nm using the microplate reader (Sunrise Tecan, Austria).
Dot blot: In order to evaluate the humoral immune response against the recombinant HCV-core protein, the protein in different concentrations (1:100 and 1:500) were dotted on a nitrocellulose paper, the primary (Immunized sera) and secondary antibodies (conjugatedhuman anti-IgG peroxidase) were added, subsequently the presence of antibodies against recombinant HCV-core protein were detected by adding substrate (30% H2O2 and DAB) (Merck, Germany) diluted in 1 M Tris (Merck, Germany) and TBS.
Statistical analysis: Data were analyzed using SPSS version 16. The one-way ANOVA was used to compare the means. The values are significant at p<0.05.
RESULTS
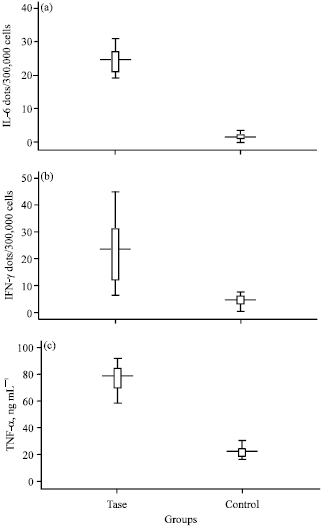
According to the result of the Elispot tests there is a significant difference between the IFN-γ level in case and control group (18.53±3.87, p = 0.001) as well as IL-6 level (22.9±1.26, p = 0.000). Moreover, similar to pervious cytokines it has been observed that the TNF-α had dramatically higher level in recombinant protein treated group compared to the control group (53.96±4.54, p = 0.000) (Fig. 2).
In this study, the MTT assay reproducibility was considerable. The cells proliferation and viability were defined by color change. The absorbance was measured at 590 nm and it was higher in test group in comparison to control group with p<0.05.
The result of dot blotting approved the humoral immune system response of immunized mice against HCV core protein. The dot formation was shown in two different concentrations (1:100 and 1:500) in test group while there was no dot formed by the control group (Fig. 3).
 | |
| Fig. 2: | IFN-γ, IL-6 and TNF-α in test and control groups |
 | |
| Fig. 3(a-c): | Dot Blot result, the sera of the control group and the immunized mice |
DISCUSSION
HCV causes silent infection and it has been reported that its existence would possibly be destructive for host cells; moreover, the core protein has been mostly detected in chronically infected HCV patients (Torbati et al., 2012). It has also been reported that Hepaciviruses core protein is unique among the Flaviviridae (Boulant et al., 2005). On the other hand, it has observed that the Cytokines and chemokines play key roles in responses against viral infections and viral immunopathology related to specific cellular receptors, a vast number of viral glycoproteins could stimulate the cytokines and chemokines secretion directly. Moreover, viral RNA and some viral proteins interfere with cellular signal transduction and transcription factor activities, Such as NF-κB that plays an important role such as expression of cytokines and chemokines is considered (Mogensen and Paludan, 2010). As a result, a protein which could stimulate the cytokine secretion would have a critical role in viral clearance and prevention. Thus the recombinant HCV-Core protein which was reported in our previous study was used and the cytokine secretion in immunized mice was analyzed in order to assess the capability of the recombinant HCV-core protein as an effective vaccine candidate (Torbati et al., 2012).
Interleukin 6 (IL-6) is a multifunctional cytokine that has critical roles in acute phase reactions, hematopoiesis, host defense and bone remodeling (Hirano et al., 1986), it has been reported by Feldmann et al. (2006) that elevated secretion of IL-6 through stimulation of TLR2 by HCV core protein can affect the pathogenesis of hepatitis C-associated B-cell non-Hodgkin's lymphoma . In another research conducted by Sekiguchi et al. (2012) they have stated that the amount of inflammatory cytokines (tumor necrosis factor) TNF-α and IL-6 is related to the onset of chronic hepatitis in mice.
Mouse TNF-α is a pleiotrophic cytokine that plays a crucial role in the hepatitis C inflammatory processes and apoptosis, TNF-α induces cell death that can be enhanced by nuclear factor kappa B (NF-κB) activation (Park et al., 2012). Mouse cells known to express TNF-α include neurons, CD8+T cells, mast cells, macrophages, monocytes and dendritic cells (Lees et al., 1999). Park et al (2012). have been reported that HCV infection increases TNF-α-induced cell death by suppressing NF-κB activation via the action of core, NS4B and NS5B proteins which could lead to immune-mediated liver injury in HCV infection (Park et al., 2012).
IFN-γ is the only member of the type II class of interferons and it is a dimerized soluble cytokine (Thiel et al., 2000). IFN-γ is critical for innate and adaptive immunity against viral and intracellular bacterial infections as well as tumor control, IFN-γ is largely produced by Natural Killer T (NKT) cells and Natural Killer (NK) and by CD4 Th1 and CD8 cytotoxic T lymphocyte (CTL) effector T cells (Schoenborn and Wilson 2007). Kang et al. (2003) has examined the HCV-specific interferon-γ responses to HCV core and NS3A proteins using enzyme-linked immunosorbent spot and it has been observed that the level of IFN-γ is increased in the immunized group as well as patients with Hepatitis C infection.
CONCLUSION
It has been concluded that the amounts of IL-6, TNF-α and IFN-γ are increased in patients with acute HCV infection, thus a vaccine which could stimulate the secretion of these cytokines in advance may have a preventive role. Based on the results of this study, a significant difference between the amounts of cytokine secretion in immunized mice in response to recombinant protein of HCV-core protein and the control group was observed.
ACKNOWLEDGMENT
We would like to thank the staff of Shahid Beheshti University of medical sciences and Tehran university of medical sciences.
REFERENCES
- Acosta-Rivero, N., J. Poutou, L. Alvarez-Lajonchere, I. Guerra and Y. Aguilera et al., 2009. Recombinant in vitro assembled hepatitis C virus core particles induce strong specific immunity enhanced by formulation with an oil-based adjuvant. Biol. Res., 42: 41-56.
PubMed - Boulant, S., C. Vanbelle, C. Ebel, F. Penin and J.P. Lavergne, 2005. Hepatitis C virus core protein is a dimeric alpha-helical protein exhibiting membrane protein features. J. Virol., 79: 11353-11365.
CrossRefDirect Link - Feldmann, G., H. D. Nischalke, J. Nattermann, B. Banas and T. Berg et al., 2006. Induction of interleukin-6 by hepatitis C virus core protein in hepatitis C-associated mixed cryoglobulinemia and B-cell non-Hodgkin's lymphoma. Clin. Cancer Res., 12: 4491-4498.
PubMed - Hirano, T., K. Yasukawa, H. Harada, T. Taga and Y. Watanabe et al., 1986. Complementary DNA for a novel human interleukin (BSF-2) that induces B lymphocytes to produce immunoglobulin. Nature, 324: 73-76.
CrossRef - Kang, F., W. Chen, X. Zhang, W. Nie and J. Fu et al., 2013. Transient liver injury associated with the early recovery of HCV-specific T-cell responses and HCV rebound in HIV-1/HCV coinfected patients undergoing highly active antiretroviral therapy. J. Acquir. Immune. Defic. Syndr., 62: 135-142.
CrossRefPubMed - Lapinski, T.W., 2001. The levels of IL-1beta, IL-4 and IL-6 in the serum and the liver tissue of chronic HCV-infected patients. Arch. Immunol. Ther. Exp. (Warsz), 49: 311-316.
PubMed - Lees, C.J., V. Apostolopoulos and I.F. McKenzie, 1999. Cytokine production from murine CD4 and CD8 cells after mannan-MUC1 immunization. J. Interferon Cytokine Res., 19: 1373-1379.
PubMed - Mogensen, T.H. and S.R. Paludan, 2010. Molecular pathways in virus-induced cytokine production. Microbiol. Mol. Biol. Rev., 65: 131-150.
CrossRef - Murata, K., M. Lechmann, M. Qiao, T. Gunji, H.J. Alter and T.J. Liang, 2003. Immunization with hepatitis C virus-like particles protects mice from recombinant hepatitis C virus-vaccinia infection. Proc. Natl. Acad. Sci. USA, 100: 6753-6758.
CrossRefPubMedDirect Link - Najafizadeh, M., N. Farhadi and B. Sarkari, 2007. The cytokine profiles in hepatitis C virus infected patients and their contribution to inflammatory responses. Shiraz E Med. J., 8: 22-27.
Direct Link - Schoenborn, J.R. and C.B. Wilson, 2007. Regulation of interferon-gamma during innate and adaptive immune responses. Adv. Immunol., 96: 41-101.
PubMed - Sekiguchi, S., K. Kimura, T. Chiyo, T. Ohtsuki and Y. Tobita et al., 2012. Immunization with a recombinant vaccinia virus that encodes nonstructural proteins of the hepatitis C virus suppresses viral protein levels in mouse liver. PLoS One.
CrossRef - Thiel, D.J., M.H. le Du, R.L. Walter, A. D'Arcy and C. Chene et al., 2000. Observation of an unexpected third receptor molecule in the crystal structure of human interferon-gamma receptor complex. Structure, 8: 927-936.
PubMed - Torbati, E., M. Bandehpour, P. Pakzad, N. Mosaffa, A. Koochaki and B. Kazmei, 2012. The evaluation of hepatitis C virus core antigen in immunized BALB/c mice. Hepat Mon., 12: 391-397.
PubMedDirect Link