
Jean Aghofack-Nguemezi
Department of Plant Biology, Faculty of Sciences, University of Dschang, P.O.Box 67, Dschang, Cameroon
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 24 | Page No.: 1964-1970







ABSTRACT
It is generally accepted that plastids play a major role in the synthesis of fatty acids. However, the degree of importance of the chloroplast integrity is not yet well established. In order to determine the effects of alterations in the chloroplast ultrastructure on this process Phaseolus aureus seedlings, species very sensitive to phase-shifts between light and temperature, were grown under control (12/12 h, 32/10°C, light/dark) or inverse (12/12h, 10/32°C, light/dark) conditions. Leaf sections were examined with an electron microscope and the fatty acid contents in the leaves and hypocotyls analyzed using a gas chromatograph. The electron microscopy of chloroplasts showed that unlike normal seedling leaves, there were few thylakoid membranes and no stacking of these membranes into grana occurred in the leaves of inverse seedlings. The levels of fatty acids in the leaves of normal seedlings (e.g., α-linolenic acid, 25 to 70 μg g-1) were always higher than those found in inverse seedling leaves (e.g., α-linolenic acid, 10 to 26 μg g-1). However, in leaves of both normal and inverse seedlings rhythmic fluctuations in the levels of fatty acids with 16 to 18 carbon atoms were observed. Furthermore, the fatty acid contents in hypocotyls of both types of seedlings were almost similar throughout the duration of the experiment. These results suggested that the high network density of thylakoid membranes and their stacking in places into grana are not prerequisites for the synthesis and/or conversion of fatty acids but would rather condition an optimal biogenesis rate and that light/dark cycles might be determinant factors in the induction of rhythmic fluctuations in fatty acid levels in plant leaves.
PDF Abstract XML References Citation
Received: April 25, 2013;
Accepted: May 18, 2013;
Published: September 16, 2013
How to cite this article
Jean Aghofack-Nguemezi, 2013. Rhythmic Changes in the Levels of Fatty Acids in Leaves of Phaseolus
aureus Seedlings did not Tightly Depend upon High/Low Temperatures Cycles and Alterations in Chloroplast Ultrastructure. Pakistan Journal of Biological Sciences, 16: 1964-1970.
DOI: 10.3923/pjbs.2013.1964.1970
URL: https://scialert.net/abstract/?doi=pjbs.2013.1964.1970
DOI: 10.3923/pjbs.2013.1964.1970
URL: https://scialert.net/abstract/?doi=pjbs.2013.1964.1970
INTRODUCTION
Under natural conditions, plants are exposed to light/dark cycles with temperatures in the dark periods being lower than those in the light periods. In thermoperiodic cycles, the high temperature pulse is normally interpreted by the plants as light and the low temperature pulse as darkness. There is a predominance of different processes at dark and at light, of which the dark process has a much lower optimum temperature than the light process (Lecharny et al., 1990; Went, 1944a, b).
When grown under greenhouse conditions whereby the light periods are impaired with high temperatures (inverse growth conditions), leaves of Chenopodium rubrum seedlings showed a very low level of chlorophyll and considerable changes in the structure of plastids (Schwall et al., 1985). Plants of tropical or subtropical origin (e.g., bean, tomato) are even more sensitive to phase-shifts between light and temperature (Aghofack-Nguemezi et al., 1991; King et al., 1982; Simillie, 1979). It is well established that the synthesis of fatty acids is a co-operation between plastids and other cell compartments (Wang and Hildebrand, 1988; Slack et al., 1976). Generally, fatty acids are synthesized de novo in plastids and then transported to the reticulum endoplasmic where they are further modified before their incorporation into specific complex lipid classes (Sommerville et al., 2000; Harwood and Moore, 1989; Gounaris et al., 1986; Thompson, 1989). However, the degree of implication of each chloroplast region (envelope membranes, stroma and thylakoid membranes) in the de novo synthesis of fatty acid is not yet exactly known. In the present study, we compared daily changes in the contents of fatty acids in leaves and hypocotyls of normal and inverse Phaseolus aureus seedlings in order to contribute to the elucidation of the role of chloroplast in the synthesis of fatty acids.
MATERIALS AND METHODS
Plant materials: Seeds of Phaseolus aureus ROXB were sown in pots containing moistened perlite. The pots were covered and placed in two growth cabinets under different conditions: 12/12h, 32/10°C, light/dark for normal growth conditions and 12/12h, 10/32°C, light/dark for inverse growth conditions. White fluorescent light (8.66 W m-2) was provided by Philips TLM 120 W/33 RS fluorescent tubes. Four days after sowing, the covers were removed and seedlings were watered with Hoagland’s solution (Hoagland and Arnon, 1950). Leaves and hypocotyls of six to eight days old seedlings were harvested, weighed fresh, boiled for five minutes and stored in liquid nitrogen at -20°C. At each harvestperiod, some samples were used for the determination of dry matter weight.
Electron microscopy: Leaf sections (1x1.5 mm) were fixed in 0.1 M phosphate buffer (pH 7.2) containing 2% glutaraldehyde for five hours at 20°C in darkness. Samples were subsequently buffer-washed, postfixed in 0.1 M buffer containing 1% OsO4 for three hours, buffer-washed and dehydrated through graded ethanol series. They were then infiltrated via propylene oxide series and embedded in araldite. Sections were stained for five minutes in saturated aqueous uranyl nitrate, five minutes in Pb citrate and examined with a Philips electron microscope.
Lipid extraction and analysis of fatty acids: Total lipids were extracted using a 5:1.9:0.5 mixture of chloroform, methanol and water. Fatty acids were then transmethylated using bortrifluoromethanol. A gas chromatography of fatty acid methylesters was performed as described by Aghofack-Nguemezi and Tatchago (2006). The fatty acid methylesters were identified on chromatograms in comparison with authentic standards. Fatty acid contents were determined using heptadecanoic acid as internal standard. All fatty acid standards and chemicals were purchased from Sigma.
Statistical analysis: Group comparisons were made using One-Way Analysis of Variance (ANOVA) to see if variations among total fatty acid level means were significantly greater than expected by chance. The Student-Newman-Keuls Test was used to compare means differences. In some few cases where differences between standard deviations of means were significant, the GraphPad Instat-software automatically proposed the use of rather the Nonparametric ANOVA and Kruskal-Wallis (as post-test).
RESULTS
Chloroplasts in normal seedling leaves presented the three known major characteristic structural regions: (a) a pair of outer membranes, the chloroplast envelope, (b) an amorphous matrix, the stroma and (c) a highly organized dense membrane network in the stroma formed of flat compressed vesicles (the thylakoid membranes) which are in places stacked into grana (Fig. 1a). In the leaves of inverse seedlings there were very few thylakoid membranes as compared to normal leaves.
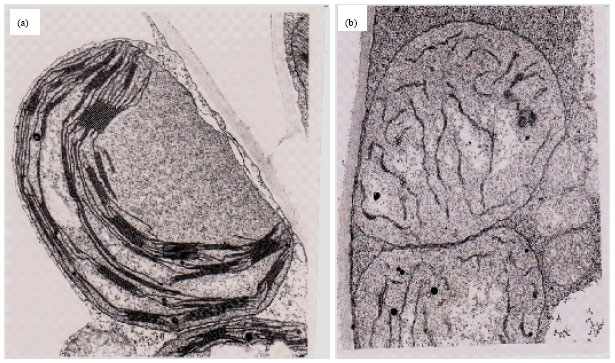 | |
| Fig. 1: | Electron micrographs of chloroplasts from leaves of the Phaseolus aureus seedlings grown under (a) Normal and (b) Inverse conditions. The magnification were X34800 and X27750, respectively |
Furthermore, the thylakoid membranes in inverse leaves were not stacked into grana (Fig. 1b).
As general trends, the levels of all fatty acids analyzed were always higher in leaves of normal than inverse seedlings (Fig. 2-6). The level of palmitic acid (C16:0) in leaves of normal seedlings peaked cyclically six hours after the onset the light period.
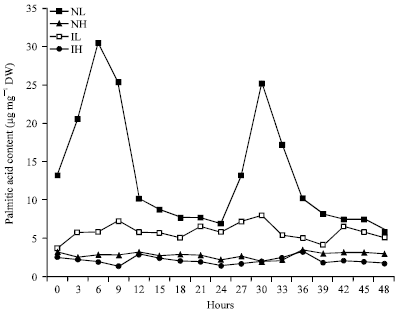 | |
| Fig. 2: | Palmitic acid levels in the leaves and hypocotyls during two 12/12h light/dark cycles. Value are means of five samples, DW: Dry mattes weight, IH: Inverse hypocottyls, Il: Inverse leaves, NH: normal hypocotyls, NL: Normal leaves |
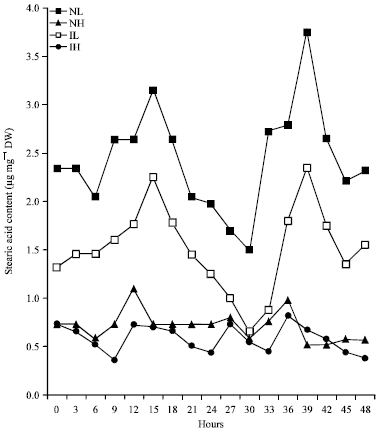 | |
| Fig. 3: | Stearic acid levels in the leaves and hypocotyls during two 12/12h, light/dark cycles. Values are means of five samples. DW: Dry mattes weight, IH: Inverse hypocottyls, Il: Inverse leaves, NH: Normal hypocotyls, NL: Normal leaves |
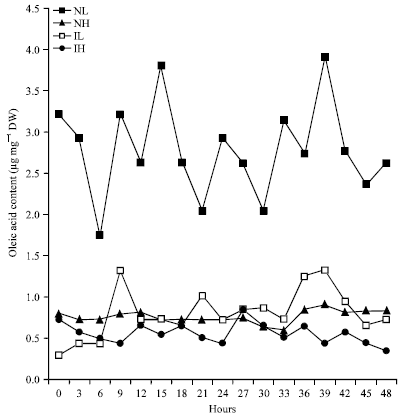 | |
| Fig. 4: | Oleic acid levels in the leaves and hypocotyls during two 12/12h light/dark cycles. Values are means of five samples. DW: Dry mattes weight, IH: Inverse hypocottyls, Il: Inverse leaves, NH: Normal hypocotyls, NL: Normal leaves |
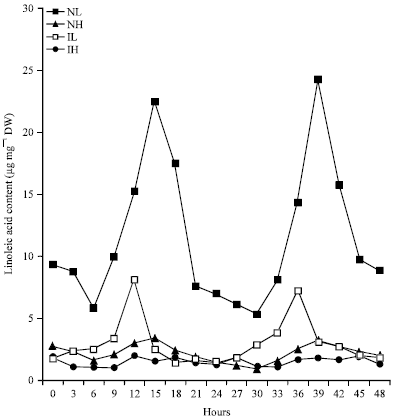 | |
| Fig. 5: | Linoleic acid levels in the leaves and hypocotyls during two 12/12h light/dark cycles. Values are means of five samples. DW: Dry mattes weight, IH: Inverse hypocottyls, Il: Inverse leaves, NH: Normal hypocotyls, NL: Normal leaves |
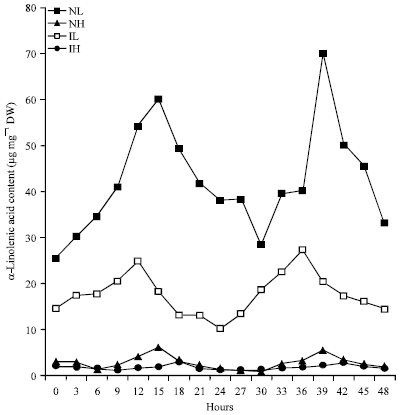 | |
| Fig. 6: | α-Linolenic acid levels in the leaves and hypocotyls during two 12/12h light/dark cycles. Values are means of five samples. DW: Dry mattes weight, IH: inverse hypocottyls, Il: Inverse leaves, NH: normal hypocotyls, NL: Normal leaves |
This fatty acid was found at almost constant concentrations in leaves of inverse seedlings and in cotyledons of both normal and inverse seedlings (Fig. 2). In the leaves of both normal and inverse seedlings the level of stearic (C18:0) also changed rhyhtmically with maximum values measured each first three hours in the dark. No clear peaks could be observed in the variations of stearic acid content in the hypocotyls of both types of seedlings (Fig. 3). The level of oleic acid (C18:1) in leaves and hypocotyls of normal and inverse seedlings remained nearly constant during the experiment (Fig. 4). Diurnal changes in the level of linoleic acid (C18:2) in leaves of normal seedlings showed maximum values three hours after the onset of the dark period. In leaves of inverse seedlings maxima in the level of C18:2 were rather measured 12 h after the start of the illumination (i.e., at the end of the light period). Concentrations of this fatty acid were almost stagnant in the hypoctyls of both normal and inverse seedlings (Fig. 5). Daily variations in the α-linolenic acid (α-C18:3) levels in leaves and hypocotyls of normal and inverse seedlings showed similar behaviour as changes in the concentrations of C18:2 (Fig. 6).
DISCUSSION
The results showed that there were alterations in the structure of chloroplast thylakoid membranes meanwhile the envelope membranes still remained seemingly normal in leaves of inverse Phaseolus aureus seedlings. Furthermore, the levels of all fatty acids analyzed were always higher in normal than inverse leaves. The ultrastructure of chloroplast in inverse leaves did not resemble that of the etioplast as described elsewhere (Taiz and Zeiger, 1998; Sperling et al., 1998). However, the inverse growth conditions obviously led to the formation of photosynthetically non-functional chloroplast since thylakoid membranes are sites of the light-driven reactions of photosynthesis (Opik et al., 2005). The envelope membranes are sites of diverse complex metabolic pathways linked to the physiological functions of the chloroplast (Joyard et al., 2009; Block et al., 2007; Joyard et al., 1998). They concentrate for example most the proteins involved in the chloroplast glycerolipid metabolism whereas the stroma embeds the initial steps of fatty biosynthesis (Joyard et al., 2010). A direct role of light and/or temperature in the accumulation of fatty acid (Kuiper, 1985; Cheesbrough, 1989; Williams et al., 1988) can’t be taken into account to explain the differences between the levels of fatty acids in the leaves of normal and inverse seedlings. An indirect control by light of the accumulation of fatty acids through the energy state of chloroplasts as postulated by Eastwell and Stumpf (1983) could explain why the levels of fatty acids were generally lower in inverse than normal leaves. With an abnormal ultrastructure of chloroplasts (Fig. 1b) and very low concentrations of chlorophyll (Aghofack-Nguemezi et al., 1991; Schwall et al., 1985), leaves of inverse seedlings could not efficiently convert light energy into ATP and transport electrons to NADP+. Thus, an anabolic process like fatty acid synthesis which needs ATP and NADPH+H+ was altered. Furthermore, plastids of aerial parts of inverse seedlings might lack triosephosphates resulting from photosynthetic CO2-fixation. Indeed, acetyl-SCoA, the primary substrate for fatty acid synthesis pathway, is formed in chloroplasts of green plants from triosephosphates but can also be provided to chloroplasts by the mitochondria (Gounaris et al., 1986; Stumpf and Shimakata, 1983). In yellow inverse leaves only the latter source of acetyl-SCoA could exist. Even in green plants, hypocotyls are not active sites of photosynthesis (Mohr and Schopfer, 1985). This might be the reason why the levels of fatty acids were almost similar in normal and inverse hypocotyls.
In the leaves of normal seedlings maxima in the levels of C16:0 were measured six hours after the start of the illumination whereas the levels of C18:0, C18:2 and α-C18:3 peaked three hours after the onset of the dark period. This temporal difference in the attainment of maximum concentrations in normal leaves might be due to the fact that C16:0 was synthesized de novo in the chloroplast stroma and then subsequently transported to the reticulum endoplasmic where further modifications (elongation and/or desaturation) for the production of C18 fatty acids took place (Sommerville et al., 2000; Harwood and Moore, 1989; Gounaris et al., 1986; Thompson, 1989). The absence of rhythmicity in changes in the levels of C16:0 in the inverse leaves could be related to the lack of functioning photosynthetic apparatus and as consequence a lack of light-dependent photosynthesis-based regulation of de novo synthesis of fatty acids (Eastwell and Stumpf, 1983). Furthermore, maxima in the levels of the C18:2 and α-C18:3 in inverse leaves were measured 12 h after the onset of the illumination instead of 15 h (i.e., three hours after the onset of the dark period) as observed in normal leaves. This was an indication that the desaturation reactions occurred differentially in the two types of leaves. In plants there exist two pathways of fatty acid desaturation (Mazliak, 1973; Hess, 1975): (a) the desaturation occurs after the completion of elongation and (b) the desaturation precedes the completion of the elongation. In inverse leaves C18:2 and α-C18:3 could be synthesized from C14:2 and C14:3 according to the latter pathway.
CONCLUSION
This study has revealed that the contents of fatty acids in the leaves of Phaseolus aureus changed rhythmically independently on alterations in chloroplast ultrastructure and diurnal changes in temperatures. The non-stacking of thylakoid membranes into grana rather brought about low rates of accumulation of fatty acids. Due probably to their status of photosynthetically almost inactive tissues, hypocotyls of green and yellow inverse plants contains similar amounts of fatty acids that were lower than those measured in the leaves. Detailed comparative studies on the analysis of fatty synthase activity and isoenzyme pattern, on the metabolism of radiolabeled CO2 and acetyl-SCoA in leaves of normal and inverse seedlings might bring more insight on the mechanisms of organelle co-operation during fatty acid biosynthesis.
REFERENCES
- Aghofack-Nguemezi, J. and V. Tatchago, 2006. Changes in glycerolipid and sterol contents in leaves of Borago officinalis L. plants as related to growth and development. Pak. J. Biol. Sci., 9: 686-691.
CrossRefDirect Link - Aghofack-Nguemezi, J., A. Christmann, S. Frosch, A. Tremolieres and E. Wagner, 1991. Contrasting photo-and thermoperiod-induced changes in abscissic acid and lipid contents in leaves of mungbean seedlings. Physiol. Plant, 83: 346-352.
Direct Link - Block, M.A., R. Douce, J. Joyard and N. Rolland, 2007. Chloroplast envelope membranes: A dynamic interface between plastids and the cytosol. Photosynth. Res., 92: 225-244.
CrossRef - Cheesbrough, T.M., 1989. Changes in the enzymes for fatty acid synthesis and desaturation during acclimation of developing soybean seeds to altered growth temperature. Plant Physiol., 90: 760-764.
CrossRef - Eastwell, K.C. and P.K. Stumpf, 1983. Regulation of plant acetyl-CoA carboxylase by adenylate nucleotides. Plant Physiol., 72: 50-55.
CrossRef - Harwood, J. and T.S. Moore, 1989. Lipid metabolism in plants. Crit. Rev. Plant Sci., 8: 1-43.
CrossRefDirect Link - Joyard, J., M. Ferro, C. Masselon, D. Seigneurin-Berny, D. Salvi, J. Garin and N. Rolland, 2010. Chloroplast proteomics highlights the subcellular compartmentation of lipid metabolism. Prog. Lipid Res., 49: 128-158.
CrossRef - Joyard, J., M. Ferro, C. Masselon, D. Segneurin-Berny, D. Salvi, J. Garin and N. Rolland, 2009. Chloroplast proteomics and the compartmentation of plastidial isoprenoid biosynthetic pathways. Mol. Plant, 2: 1154-1180.
CrossRef - Joyard, J., E. Teyssier, C. Miege, D. Berny-Seigneurin and E. Marechal et al., 1998. The biochemical machinery of plastid envelope membranes. Plant Physiol., 118: 715-723.
Direct Link - King, A.I., M.S. Reid and B.D. Patterson, 1982. Diurnal changes in the chilling sensitivity of seedlings. Plant Physiol., 70: 211-213.
CrossRef - Kuiper, P.J.C., 1985. Environmental changes and lipid metabolism of higher plants. Physiologia Plantarum, 64: 118-122.
CrossRef - Lecharny, A., A. Tremolieres and E. Wagner, 1990. Correlation between the endogenous circadian rhythmicity in growth rate and fluctuations in oleic acid content in expanding stems of Chenopodium rubrum L. Planta, 182: 211-215.
CrossRef - Schwall, M., B. Kropp, V. Steinmetz and E. Wagner, 1985. Diurnal modulation of phototropic response by temperature and light in Chenopodium rubrum L. as related to stem extension rate andarginine decarboxylase activity. Photochem. Photobiol., 42: 753-757.
CrossRef - Slack, C.R., P.G. Roughan and J. Terpstra, 1976. Some properties of a microsomal oleate desaturase from leaves. Biochem. J., 155: 71-80.
Direct Link - Sperling, U., F. Franck, B. van Cleve, G. Frick, K. Apel and G.A. Armstrong, 1998. Etioplast differentiation in arabidopsis: Both PORA and PORB restore the prolamellar body and photoactive protochlorophyllide-F655 to the cop1 photomorphogenic mutant. Plant Cell, 10: 283-296.
CrossRef - Went, F.W., 1944. Plant growth under controlled conditions. Thermoperiodicity in growth and fruiting of tomato. Am. J. Bot., 31: 135-150.
Direct Link - Wang, X.M. and D.F. Hildebrand, 1988. Biosynthesis and regulation of linolenic acid in higher plants. Plant Physiol. Biochem., 26: 777-792.
Direct Link - Went, F.W., 1944. Plant growth under controlled conditions. III. Correlation between various physiological process and growth in tomato plants. Am. J. Bot., 31: 597-618.
Direct Link