
A. A. Amin
Department of Botany, National Research Centre, Cairo, 126221, Egypt
F. A. Gharib
Department of Botany, Faculty of Science, Helwan University, Cairo, 11795, Egypt
H. F. Abouziena
Department of Botany, National Research Centre, Cairo, 126221, Egypt
Mona G. Dawood
Department of Botany, National Research Centre, Cairo, 126221, Egypt
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 24 | Page No.: 1894-1903







ABSTRACT
The response of chickpea (Cicer arientinum L. cv. Giza 3) to treatment with two plant growth regulators putrescine (Put) and Indole-3-butyric acid (IBA) at 25, 50 and 100 mg L-1 applied either alone or in combinations was studied. Spraying of Put and IBA either individually or in combination significantly increased the plant height, number and dry weight of branches, leaves and pods/plant and leaf area/plant at the two growth stages. Total photosynthetic pigments in fresh leaves were significantly promoted as a result of application of Put or IBA. Generally, application of Put and/or IBA at 100 mg L-1 produced the highest numbers of pods which resulted in substantially the highest seed yield. Put and IBA increased the seed yield by 21.3 and 19.2%, respectively, while the combination of Put at 100 mgL-1 and IBA at 50 mgL-1 increased it by 27.4%. Greatest increases in straw and biological yield/fed (38.3 and 30.4%, respectively) were noted with the combination treatment of IBA 100 mg L-1 plus Put at 100 mg L-1. Put and IBA significantly increased the nitrogen, phosphorus, potassium, total soluble sugars and total free amino acids in chickpea seeds over control, but the effects were less marked than those of their combination. This response was greater following treatment with IBA than with Put. It could be conclude that spraying Put or/and IBA on chickpea plants have promotion effects on the seeds yield criteria which have promising potential as sources of low-cost protein and minerals for possible use as food/feed supplements.
PDF Abstract XML References Citation
Received: March 11, 2013;
Accepted: March 29, 2013;
Published: September 16, 2013
How to cite this article
A. A. Amin, F. A. Gharib, H. F. Abouziena and Mona G. Dawood, 2013. Role of Indole-3-butyric Acid or/and Putrescine in Improving Productivity of Chickpea (Cicer arientinum L.) Plants. Pakistan Journal of Biological Sciences, 16: 1894-1903.
DOI: 10.3923/pjbs.2013.1894.1903
URL: https://scialert.net/abstract/?doi=pjbs.2013.1894.1903
DOI: 10.3923/pjbs.2013.1894.1903
URL: https://scialert.net/abstract/?doi=pjbs.2013.1894.1903
INTRODUCTION
Grain legumes are important crop plants for their protein rich seeds that used as a major source of dietary protein for human and livestock consumption. In addition, legumes can be efficiently used for improving soil fertility (Manchanda and Garg, 2008).
Chickpea (Cicer arietinum L.) is an important pulse crop grown and consumed all over the world, especially in the Afro-Asian countries (Jukanti et al., 2012). Chickpea is the cheapest and readily available source of protein (20.6%), (Gupta, 1987), carbohydrates (61.2%) (Gupta, 1987), rich in vitamins and minerals and is relatively free from anti-nutritional factors (Wang et al., 2010). Because of the great importance of chickpea as a cheapest and readily available source of protein food for large population groups, particularly in developing countries, many efforts have been made to improve chickpea productivity (Alvi et al., 2005; El-Kramany et al., 2009; Niari-Khamssi, 2011).
Plant growth regulators (PGRs) are representing one of the controlling factors that regulate growth, biosynthesis of chemical constituents, yield and may be improve adaptation of plants to environment. Polyamines (PAs) are polycationic compounds of low molecular weight that are present in all living organisms (Cohen, 1998). Some studies have shown the positive effects of PAs on cell division and its rate (Cvikrova et al., 1999), morphogenesis, floral initiation and development (Paschalidis and Roubelakis-Angelakis, 2005; Liu et al., 2006; Pang et al., 2007; Khan et al., 2012). PAs effectively retard senescence by retarding the loss of chlorophyll (Couee et al., 2004; Tang et al., 2004). Putrescine application (up to 5 mM) significantly increased the content of photosynthetic pigments of wheat (Triticum aestivum) plants (El-Bassiouny et al., 2008). PAs have been shown to be an integral part's response to stress (Walters, 2003; Alcazar et al., 2006; 2010). Exogenous putrescine and sermidine markedly modified the stress-induced effects in plants (Amooaghaie, 2011). Moreover, putrescine treatments significantly increased fresh and dry weights of bean plants (Nassar et al., 2003). Putrescine at 10-5 M increased grain and biological yield and grain index of wheat plant (Gupta et al., 2003). Exogenous application of putrescine and spermidine increased endogenous PGRs, particularly growth promoters (Indole-3- acetic acid, gibberellins and cytokinins) and decreased inhibitors (abscisic acid) (El-Bassiouny and Bakheta, 2005; Iqbal et al., 2006). Moreover, auxins are implicated in various metabolic processes as nucleic acid synthesis (Krishnamoorthy, 1981; Alvi et al., 2005).
Indole-3-butyric acid significantly increased the vegetative growth criteria, seed yield and protein content in rice (Chhun et al., 2004), maize and onion plants (Amin et al., 2006; 2007). Soaking chickpea seeds with 10-8 M IAA improved nitrate reductase activity, total protein, calcium and potassium contents (Alvi et al., 2005). Moreover, improvement in nitrogen metabolism, photosynthesis and yield of chickpea were noted with the application of 10-8 M of IAA, IBA, or 4-Cl-IAA (4-chloroindole-3-acetic acid) (Hayat et al., 2009).
Considering the positive benefit that Put and IBA have had on productivity of several crop plants and to ascertain whether these two PGRs would also be able to improve morphological parameters and seed nutritional value of a cereal-based diet.
Therefore, effect of spraying chickpea plants with different concentrations of putrescine and Indole-3- butyric acid, individually or in combination on some morphological criteria, yield, as well as some metabolic constituents of the seeds were assessed in this work.
MATERIALS AND METHODS
Two field experiments were carried out at the Agricultural Experimental Station, National Research Centre, Nubaria, Behira Governorate, Egypt, during the two successive seasons of 2010/2011 and 2011/2012 to study the effect of foliar application of different concentrations of Put and IBA, individually or in combination on some growth criteria, photosynthetic pigments and yield as well as some metabolic constituents of chickpea seeds.
The experimental design was split-plot design with four replications. The IBA treatment occupied the main plots and Put treatments were allocated at random in sub-plots. The plot area was 10.5 m2 (3.5x3.0 m) and consisted of five rows 70 cm apart and the distance between hills along the row 25 cm apart.
Seeds of chickpea (Cicer arietinum L.) cv. Giza 3, were obtained from Agricultural Research Centre, Giza, Egypt and sown on 23 November in both seasons. The normal cultural treatments of growing chickpea in the location were followed. Pre-sowing, 30 kg/fed. calcium super-phosphate (15.5% P2O5) was applied to the soil. While, nitrogen fertilizer (35 kg N/fed) as ammonium nitrate (33.5% N) were applied in two equal doses before the first and second irrigation, respectively.
In both seasons, a foliar spray was applied twice to chickpea plants during vegetative growth at 45 and 60 days after sowing (DAS) with putrecine (Put) as polyamine (Polycarbonic and aliphatic nitrogenous compounds [(H3N-(CH2)3-NH-(CH2)4-NH- (CH2)3-NH3])(approx 97%) obtained from Sigma Company, USA and Indole-3-butyric acid (IBA, Aldrich). The two bioregulators were applied at 25, 50 and 100 mgL-1 either individually or in combination, in all possible permutations. Solutions were sprayed over the entire surface of plant including the adaxial and abaxial surfaces of leaves. No additives or surfactants such as Tween were added. In addition, untreated plants (control) were sprayed only with tap water.
The plant growth characters were measured at 75 and 90 days after sowing i.e., green yield as plant height, number of branches, leaves and pods/plant, dry weight of branches, leaves and pods/plant and leaf area (cm2/plant) following the suggestions of Bremner and Taha (1966). Photosynthetic pigment (chlorophylls a, b, carotenoids as well as total pigments) in the fresh leaves was determined at two growth stages (75 and 90 DAS) according to Saric et al. (1967).
At harvest time, ten guarded plants were taken out randomly from the middle two ridges of each plot to determine the mean values of yield and its related parameters, i.e. plant height, number of pods and seeds/plant, dry weight of pods/plant, seed and straw yield per plant and harvest index [(grain yield/ biological yield) x100]. Seed, straw and biological yields per feddan (fed = 4200 m2) were determined by harvesting the whole plot area.
Plant samples were taken from each plot and dried at 70°C for 48 h until constant dry weight. Representative dry samples of seeds were taken from each plot to determine the total soluble sugars using the phenol sulphuric method (Dubois et al., 1956). Crude protein percentage was calculated by multiplying the values of total nitrogen by 6.25 (AOAC, 1988). Phosphorus was determined according to Watanab and Olsen (1965) and potassium by Jackson (1965).The method of Rosen (1957) was used to estimate the total free amino acids.
Data for both growing seasons was carried out according to Snedecor and Cochran (1990). Data was combined since CV% was <5%. Treatments means were compared using Fisher's least significant differences (LSD) at p = 0.05.
RESULTS
Growth parameters: Data presented in Table 1 and 2 show that spraying Indole-3-butyric acid (IBA) and putrecine (Put) at 25, 50 and 100 mg L-1 individually or in combination promoted almost growth criteria compared to corresponding untreated plants.
| Table 1: | Effect of Indole-3-butyric acid (IBA) and Putrescine (Put) on growth characters of chickpea plants at (A) 75 and (B) 90 days after sowing. (Combined analysis of two seasons) |
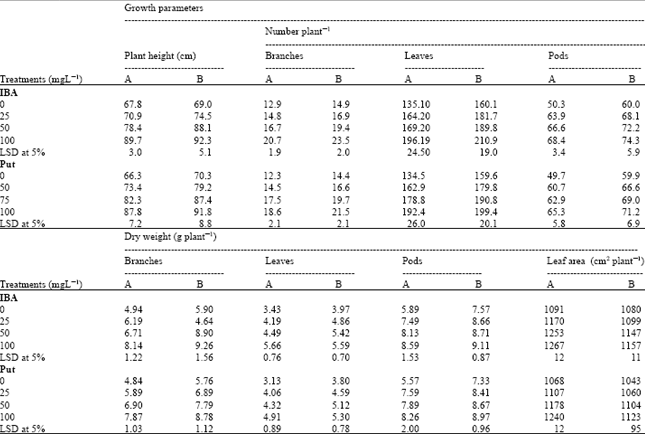 | |
In all cases, the increments in growth parameters were often highly significant in comparison with untreated control plants. The most effective treatments on most growth parameters were the combination between Put at 100 mg L-1 and IBA at 100 mg L-1 followed by IBA alone at 100 mg L-1 treatment.
When considered Put as a single factor, The results in Table 1 illustrate that application of Put up to 100 mg L-1 significantly increased the plant height, number of branches, leaves and pods/plant, dry weight of branches, leaves and pods/plant and leaf area compared to untreated plants at 75 and 90 days after sowing (DAS). Furthermore, when considering IBA as a single factor, it was noted that IBA treatments were more effective than Put treatments in increasing vegetative growth of chickpea plants at the two different growth stages (Table 1).
The increment in growth characters studied was more pronounced with spraying IBA at 100 mg L-1. An increase of 57, 42 and 20% over control occurred in dry weight of branches, leaves and pods at 90 DAS with the application of (100 mg L-1) IBA on chickpea plants, respectively. While the Put showed corresponding increments 52, 40 and 22%, respectively (Table 1).
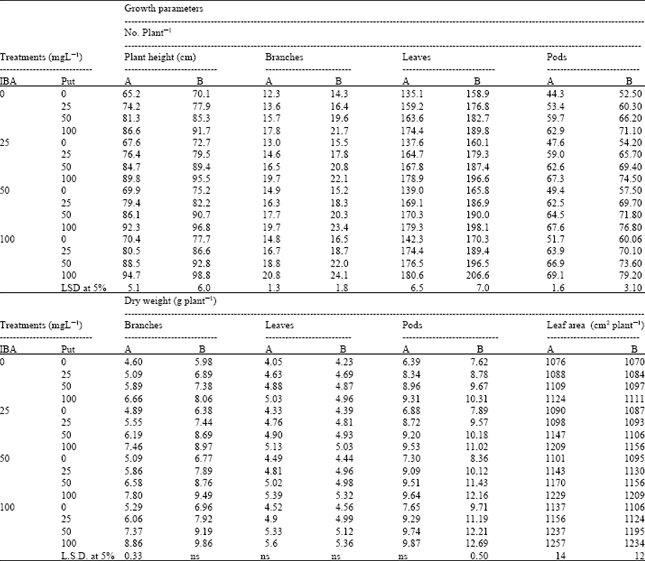
Data in Table 2 show significant increases in some growth parameters under the effect of combination of different concentrations of Put and IBA when compared to control plants. The combination treatment of IBA at 100 mg L-1 + Put at 100 mg L-1 enhanced the most growth parameters-to great extent- at the two physiological growth stages. This superior combination treatment caused a significant increase (17%) in the leaf area/plant at 75 DAS compared to untreated plants.
Photosynthetic pigments: Data presented in Table 3 show that Chl a, b, carotenoids and total photosynthetic pigments in chickpea leaves reached a maximum value at 90 DAS. Generally, foliar application with either Put or IBA at any concentration significantly increased the Chl a, b and carotenoids and consequently the total pigments more than controls at 75 and 90 DAS. The most effective concentration was 100 mg L-1 of either IBA or Put at the two growth stages.
It is worthy to mention that IBA treatments were more effective in increasing the Photosynthetic pigments than Put treatments. IBA treatments had much higher photosynthetic pigments than Put by 5 and 4% at 75 and 90 days after sowing, respectively and it increased by 28 and 27%, respectively, as compared to controls (Table 3).
| Table 2: | Effect of interaction between Indole-3-butyric acid (IBA) and Putrescine (Put) on growth characters of chickpea plants at (A) 75 and (B) 90 days after sowing. (Combined analysis of two seasons) |
 | |
| Table 3: | Effect of Indole-3-butyric acid (IBA) and Putrescine (Put) on the photosynthetic pigments content (mg/g fresh weight) in the leaves of chickpea plants at 75 and 90 days after sowing. (Combined analysis of two seasons) |
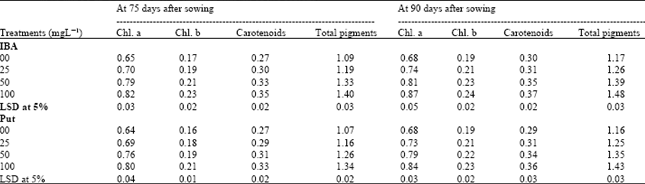 | |
On the other hand, the interactions between IBA and Put show insignificant effects on Chl. a, b, carotenoids and total photosynthetic pigments content in chickpea leaves at the two stages of growth, therefore the data were excluded.
| Table 4: | Effect of Indole-3-butyric acid (IBA), Putrescine (Put) and their interaction on yield characters of chickpea plants. (Combined analysis of two seasons) |
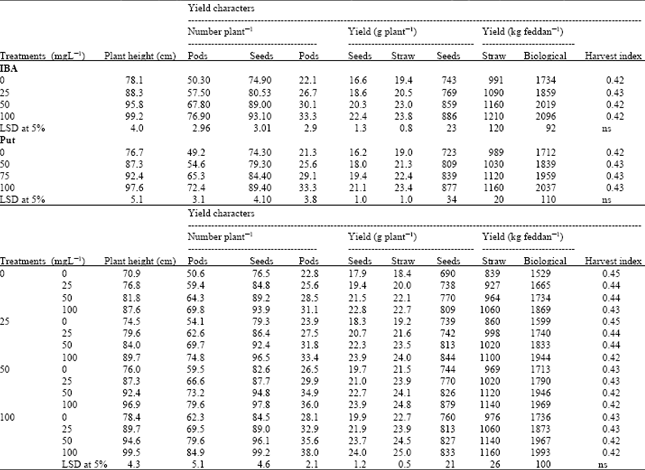 | |
Yield and its components: Data presented in Table 4 show that application of Put alone up to 100 mg L-1 significantly increased the chickpea yield criteria (number of branches, pods and seeds/plant, seed and straw yield per plant and per fed and biological yield/fed, while harvest index were not differ than control plants. This response was greater following treatment with IBA than with Put.
Application of IBA and Put increased the seed yield by 21.3, 18.3%, respectively, while the combination treatment of IBA at 50 mgL-1 and Put at 100 mgL-1 increased it by 27.4%, compared to untreated plants (Table 4).
Application of IBA exhibited more chickpea seed yield if compared to the untreated plants (Table 4). The most promising results (0.88, 1.21 and 2.10 t/fed) were obtained with 100 mg L-1 IBA for seed, straw and biological yield, respectively.
Furthermore, chickpea yield was more sensitive to the interaction between Put and IBA (Table 4). The highest increase in number of pods and seeds/plant, dry weight of pods/plant, seeds and straw yield/plant, as well as seed, straw and biological yield/fed were obtained by foliar application with 100 mg L-1 Put+100 mg L-1 IBA followed by 100 mg L-1 IBA (Table 4). On the other hand, the interaction between Put and IBA show insignificant changes in harvest index compared to the control plants.
Chemical constituents: Data presented in Table 5 show that application either Put or IBA at any concentration as well as their combination increased nitrogen, phosphorus, potassium, crude protein, total sugar and total free amino acids in the dry seed of chickpea compared with their control. The most effective treatment in this concern was 100 mg L-1 of either Put or IBA and their combination. Increasing the concentration of IBA from 25 to 50 and to 100 mg L-1 cause a significant increase in the values of P % only, while the rest of chemical constituents studied were not significantly affected by increasing the concentration of IBA and Put (Table 5). However the three concentration of the two bioregulators caused a significant increase in the values of chemical constituents compared to untreated plants.
| Table 5: | Effect of Indole-3-butyric acid (IBA), Putrescine (Put) and their interaction on chemical constituents of chickpea seeds (Combined analysis of two seasons) |
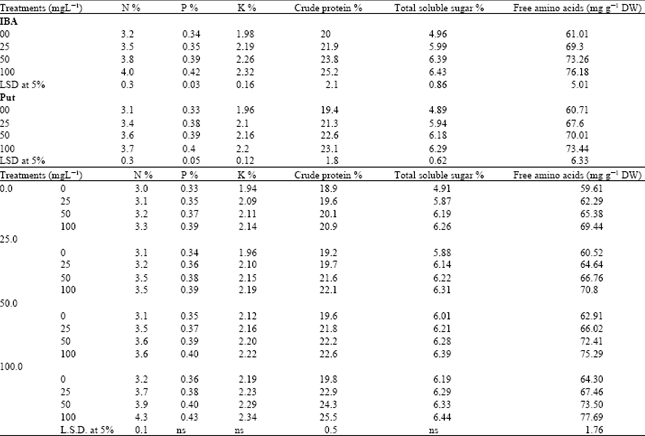 | |
Furthermore, the results in Table 5 indicate that among the treatments Put (100 mg L-1) followed by IBA (100 mg L-1) and the combination between the two bioregulators at 100 mg L-1 recorded the highest level of N, P,K, crude protein, total soluble sugars and total free amino acids.
DISCUSSION
The present study indicated that foliar application with Put and IBA up to 100 mg L-1, single or in combination greatly promoted the vegetative growth and dry matter production of branches, leaves and pods by enhancing cell division and chlorophyll accumulation. In other crops, Put stimulated growth by increasing the amount of endogenous promoters (auxin, gibberellins and cytokinins) accompanied by a decrease in the content and activity of inhibitors (ABA) (El-Bassiouny and Bakheta, 2005; El-Bassiouny et al., 2008). Abbas and El-Saeid (2012) reported that IBA treatment increased endogenous GA concentrations in lemon grass plants by 10% and 23% compared with the control for plants treated with 25 and 100 ppm, respectively. In this study, Indole-3- butyric acid has shown more pronounced effect than putrescine (100 mg L-1) in promoting growth and yield of chickpea plants may be due to increase the nitrogen uptake and regulation of different metabolic processes. Similarly, promoting effects of IBA on growth and photosynthetic pigments was obtained on maize and onion (Amin et al., 2006, 2007). Intuitively the combination of Put and IBA increased vegetative growth and dry matter production of chickpea more effective than individual treatments.
Photosynthetic pigments of chickpea leaves-at 75 and 90 DAS- were significantly enhanced by application of Put or IBA. Since, Put or IBA treatments might retard Chl destruction and/or increase their biosynthesis or stabilize the thylakoid membrane. PAs may retard senescence and chlorophyll loss by altering the stability and permeability of membranes and protecting chloroplast from senescing (Gonzalez-Aguilar et al., 1997). PAs have been shown to affect Chl stabilization and antisensecence (Da Silva, 2006; Ioannidis et al., 2009).
Put increased endogenous cytokinins, which stimulate Chl biosynthesis and chloroplast differentiation in wheat (T. vulgare) (Xie et al., 2004; El-Bassiouny et al., 2008). On the other hand, IBA increased photosynthetic pigments content due to the stimulatory effect on the amount of metabolites synthesized through enhancement of cell division and chlorophyll accumulation which leads to higher rate of photosynthesis (Ludwig-Muller, 2000). El-Tohamy et al. (2012) reported that the higher levels of putrescine significantly increased all vegetative growth parameters, yield, T.S.S. of fruits and total chlorophyll content of leaves of Cape gooseberry (Physalis peruviana L.).
Promoting effects of PAs and IBA on photosynthetic pigments was observed in cucumber (Cucumber sativus L.) (He et al., 2002); common bean (Phaseolus vulgare) (Nassar et al., 2003); maize (Zea mays L.) and onion (Allium cepa L.) (Amin et al., 2011; Ghodrat et al., 2012). Put treatments concomitantly with increasing Mg levels (a structural component of Chl) retarded senescence (Kao, 1994; El-Bassiouny et al., 2008). Although, combination of Put and IBA caused insignificant increases in photosynthetic pigments of chickpea leaves (Data not shown).
Chickpea yield and quality of seeds were significantly enhanced by application of Put and IBA, especially at the highest concentration. Rice grain filling and 1000-seed weight were positively correlated with PA content (Yang et al., 1996). On the other hand, auxins (IBA) are involved in many aspects of growth and development of plants (Ludwig-Muller, 2000).
IBA promotes growth, increases building metabolites, retards senescence, enhances cell division, chlorophyll accumulation and stimulates dry matter production as a result of higher photosynthetic activity and consequently increased translocation and accumulation of microelements in plant organs (Shafey et al., 1994; Chhun et al., 2004). IBA increased vegetative growth, seed yield per plant and per feddan, seed protein content in rice (Chhun et al., 2004), maize and onion (Amin et al., 2006, 2007). The combination between Put and IBA was more effective than Put or IBA individually for increasing the productivity of chickpea plants by enhancing photosynthetic activity, accumulating dry matter and retarding senescence. Consequently, these treatments increased translocation and accumulation of certain metabolites in plant organs which affected yield quantity and quality.
Foliar application with either Put or IBA and their combination increased nitrogen, phosphorus, potassium, crude protein, total soluble sugars percent and total free amino acids in the dry seeds of chickpea notably due to their bioregulatory effect on enzymatic activity and translocation processes from leaves to seeds, linking or converting to other plant metabolites. PAs linked with particular proteins (Serafini-Fracassini and Del Duca, 2008) and this covalently bound PA-protein complexes have been reported in artichoke tubers (Helianthus tuberosus) (Dinnella et al., 1992) and tobacco (Nicotiana spp.) (Sawhney and Applewhite, 1993). PAs are able to bind with pectin and polysaccharides (D’oraci and Bagni, 1987) and increased α-amylase activity (Tipirdamaz et al., 1995). Spm and Put significantly increased the total sugar content but decreased total free amino acids in chamomile (Matricaria chamomilla) leaves (Abd El-Wahed and Krima, 2004; Abd El-Wahed and Gamal El Din, 2005). On the other hand, IBA promotes growth, increases building metabolites and consequently increases translocation and accumulation of building metabolites to seeds. The highest values of total sugar and crude protein content in soybean, wheat, rice and maize (Dybing and Lay, 1982; Ludwig-Muller, 2000; Chhun et al., 2004) and highest total free amino acids, total phenols and total Indoles in onion (Amin et al., 2007) were obtained by foliar application of IBA at 100 mg L-1.
The combination treatments of Put and IBA at 100 mg L-1 was very effective in enhancing nitrogen, phosphorus, potassium, crude protein, total sugar content and total free amino acids and consequently increased nutritional value of chickpea seeds as a cereal-based diet more than Put and IBA used individually.
In conclusion, application of Put and IBA (up to 100 mg L-1) individually or in combination, resulted in a significant increase in morphological attribute, including number of pods, seed, straw and biological yield per fed, the main consumer-related traits. The use of Put and IBA improved the biological yield and nutritional value of chickpea seeds, the highest prior in selecting high yielding chickpea plants.
REFERENCES
- Abbas, S.M. and H.M. El-Saeid, 2012. Effects of some growth regulators on oil yield, growth and hormonal content of lemon grass (Cymbopogon citrates). Botanica Serbica, 36: 97-101.
Direct Link - Abd El-Wahed, M.S.A. and M.G.E.D. Krima, 2004. Stimulation effect of spermidine and stigmsterol on growth, flowering, biochemical constituents and essential oil of chamomile plants (Chamomilla recutita L., rausch). Bulgerian J. Plant Physiol., 30: 48-60.
Direct Link - Alcazar, R., F. Marco, J.C. Cuevas, M. Patron and A. Ferrando et al., 2006. Involvement of polyamines in plant response to abiotic stress. Biotechnol. Lett., 28: 1867-1876.
CrossRefDirect Link - Alcazar, R., T. Altabella, F. Marco, C. Bortolotti and M. Reymond et al., 2010. Polyamines: Molecules with regulatory functions in plant abiotic stress tolerance. Planta, 231: 1237-1249.
CrossRefDirect Link - Amooaghaie, R., 2011. Role of polyamines in the tolerance of soybean to water deficit stress. World Acad. Sci., Eng. Technol., 56: 498-502.
Direct Link - Alvi, S., S. Hayat and A. Ahmed, 2005. Metabolic aspects in the germinating seeds of Cicer arietinum, supplemented with auxin and/or cations. Int. J. Agric. Biol., 7: 304-307.
Direct Link - Amin, A.A., E.S.M. Rashad and F.A.E. Gharib, 2006. Physiological responses of maize plants (Zea mays L.) To foliar application of morphactin CF125 and Indole-3-butyric acid. J. Boil. Sci., 6: 547-554.
CrossRefDirect Link - Amin, A.A., E.S.M. Rashad and H.M.H. El-Abagy, 2007. Physiological effect of indole-3-butyric acid and salicylic acid on growth, yield and chemical constituents of onion plants. J. Applied Sci. Res., 3: 1554-1563.
Direct Link - Amin, A.A., F.A. Gharib, M. El-Awadi and E.S.M. Rashad, 2011. Physiological response of onion plants to foliar application of putrescine and glutamine. Scientia Horticulturae, 129: 353-360.
CrossRef - Bremner, P.M. and M.A. Taha, 1966. Studies in potato agronomy. I. The effects of variety, seed size and spacing on growth, development and yield. J. Agric. Sci., 66: 241-252.
CrossRefDirect Link - Chhun, T., S. Takcta, S. Tsurumi and I. Masahiko, 2004. Different behaviour of indole-3-acetic acid and indole-3-butyric acid in stimulating lateral root development in rice (Oryza sativa L.). Plant Growth Regul., 43: 135-143.
Direct Link - Couee, I., I. Hummel, C. Sulmon, G. Gouesbet and A. El-Amrani, 2004. Involvement of polyamines in root development. Plant Cell Tissue Organ Culture, 76: 1-10.
CrossRefDirect Link - Cvikrova, M., P. Binarova, V. Cenklova, J. Eder and I. Machackova, 1999. Reinitiation of cell division, polyamine and aromatic monoamine levels in alfalfa explant during the induction of somatic embryogenesis. Physiol. Plant, 105: 330-335.
Direct Link - Dinnella, C., D. Serafini-Fracassini, B. Grandi and S. Del Duca, 1992. The cell cycle in Helianthus tuberosus: Analysis of polyamine-endogenous protein conjugates by transglutaminase-like activity. Plant Physiol. Biochem., 30: 531-539.
Direct Link - D'Oraci, D. and N. Bagni, 1987. In vitro interactions between polyamines and pectic substances. Biochem. Biophys. Res. Commun., 148: 1159-1163.
CrossRef - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Dybing, C.D. and C. Lay, 1982. Oil and protein in field crop with morphactin and other growth regulators for senescence delay. Crop Sci., 22: 1054-1058.
Direct Link - El-Bassiouny, H.M., H.A. Mostafa, S.A. El-Khawas, R.A. Hassanein, S.I. Khalil and A.A. Abd El-Monem, 2008. Physiological responses of wheat plant to foliar treatments with arginine or putrescine. Aust. J. Basic Applied Sci., 2: 1390-1403.
Direct Link - El-Tohamy, W.A., H.M. El-Abagy, M.A. Badr, A.A. Ghoname and S.D. Abou-Hussein, 2012. Improvement of productivity and quality of cape gooseberry (Physalis peruviana L.) by foliar application of some chemical substances. J. Applied Sci. Res., 8: 2366-2370.
Direct Link - Ghodrat, V., M.J. Rousta, M.S. Tadaion and A. Karampour, 2012. Yield and yield components of corn (Zea mays L.) in response to foliar application with indole butyric acid and gibberellic acid. Am.-Eurasian J. Agric. Environ. Sci., 12: 1246-1251.
Direct Link - Gonzalez-Aguilar, G.A., L. Zacarias, M. Mulas and M.T. Lafuente, 1997. Temperature and duration of water dips influence chilling injury, decay and polyamine content in Fortune mandarins. Postharvest Biol. Technol., 12: 61-69.
CrossRef - Gupta, S., M.L. Sharma, N.K. Gupta and A. Kumar, 2003. Productivity enhancement by putrescine in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants, 9: 279-282.
Direct Link - Hayat, Q., S. Hayat, B. Ali and A. Ahmad, 2009. Auxin analogues and nitrogen metabolism, photosynthesis and yield of chickpea. J. Plant Nutr., 32: 1469-1485.
CrossRef - He, L., K. Nada and S. Tachibana, 2002. Effects of spermidine pretreatment through the roots on growth and photosynthesis of chilled cucumber plants (Cucumis sativus L.). J. Jpn. Soc. Hortic. Sci., 71: 490-498.
Direct Link - Ioannidis, N.E., S.M. Ortigosa, J. Veramendi, M. Pinto-Marijuan and I. Fleck et al., 2009. Remodeling of tobacco thylakoids by over-expression of maize plastidial transglutaminase. Biochim. Biophys. Acta (BBA)-Bioenergetics, 1787: 1215-1222.
CrossRef - Iqbal, A., N. Ateeq, I.A. Khalil, S. Perveen and S. Saleemmullah, 2006. Physicochemical characteristics and amino acid profile of chickpea cultivars grown in Pakistan. Foodservice Res. Int., 17: 94-101.
CrossRefDirect Link - Jukanti, A.K., P.M. Gaur, C.L.L. Gowda and R.N. Chibbar, 2012. Nutritional quality and health benefits of chickpea (Cicer arietinum L.): A review. Br. J. Nutr., 108: S11-S26.
CrossRefDirect Link - Khan, H.A., K. Ziaf, M. Amjad and Q. Iqbal, 2012. Exogenous application of polyamines improves germination and early seedling growth of hot pepper. Chilean J. Agric. Res., 72: 429-433.
Direct Link - Kao, C.H., 1994. Endogenous polyamine levels and dark-induced senescence of detached corn leaves. Bot. Bull. Acad. Sin., 35: 15-18.
Direct Link - Ludwig-Muller, J., 2000. Indole-3-butyric acid in plant growth and development. Plant Growth Regul., 32: 219-230.
CrossRefDirect Link - Manchanda, G. and N. Garg, 2008. Salinity and its effects on the functional biology of legumes. Acta Physiol. Plant, 30: 595-618.
CrossRef - Nassar, A.H., K.A. El-Tarabily and K. Sivasithamparam, 2003. Growth promotion of bean (Phaseolus vulgaris L.) by a polyamine-producing isolate of Streptomyces griseoluteus. Plant Growth Regul., 40: 97-106.
CrossRefDirect Link - Niari-Khamssi, N., 2011. Grain yield and protein of chickpea (Cicer arietinum L.) cultivars under gradual water deficit conditions. Res. J. Environ. Sci., 5: 611-616.
CrossRef - Pang, X.M., Z.Y. Zhang, X.P. Wen, Y. Ban and T. Moriguchi, 2007. Polyamines all-purpose players in response to environment stresses in plants. Plant Stress J., 1: 173-188.
Direct Link - Paschalidis, A.K. and A.K. Roubelakis-Angelakis, 2005. Sites and regulation of polyamine catabolism in the tobacco plant. Correlations with cell division/expansion, cell cycle progression and vascular development. Plant Physiol., 138: 2174-2184.
CrossRefDirect Link - Rosen, H., 1957. A modified ninhydrin colorimetric analysis for amino acids. Arch. Biochem. Biophys., 67: 10-15.
CrossRefPubMedDirect Link - Sawhney, R.K. and P.B. Applewhite, 1993. Endogenous protein-bound polyamines: Correlation with regions of cell division in tobacco leaves, internodes and ovaries. Plant Growth Regul., 12: 223-227.
CrossRef - Serafini-Fracassini, D. and S. Del Duca, 2008. Transglutaminases: Widespread cross-linking enzymes in plants. Ann. Bot., 102: 145-152.
CrossRefDirect Link - Wang, N., D.W. Hatcher, R.T. Tyler, R. Toews and E.J. Gawalko, 2010. Effect of cooking on the composition of beans (Phaseolus vulgaris L.) and chickpeas (Cicer arietinum L.). Food Res. Int., 43: 589-594.
CrossRefDirect Link - Watanab, F.S. and S.R. Olsen, 1965. Test of ascorbic acid methods for determining phosphorus and NaHCO3, extracts for soil. Soil Sci. Am. Proc., 29: 677-678.
Direct Link - Xie, Z., D. Jiag, T. Dai, Q. Jing and W. Cao, 2004. Effects of exogenous ABA and cytokinin on leaf photosynthesis and grain protein accumulation in wheat ears cultured in vitro. Plant Growth Regul., 44: 25-32.
CrossRef