
K.A. Obirikorang
Department of Fisheries and Watershed Management, Faculty of Renewable Natural Resources, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana
D. Adjei-Boateng
Department of Fisheries and Watershed Management, Faculty of Renewable Natural Resources, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana
H.A. Madkour
National Institute of Oceanography and Fisheries, Red Sea Branch, Hurghada, Egypt
S. Amisah
Department of Fisheries and Watershed Management, Faculty of Renewable Natural Resources, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana
F.A. Otchere
Fisina Scientific Inc. Surrey, BC. Canada
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 4 | Page No.: 185-189







ABSTRACT
The length-weight relationship of the threatened freshwater clam, Galatea paradoxa (Born 1778) from the Volta Estuary, Ghana was studied over a two-year period, from March 2008 to February 2010, to aid in the development of stock assessment models for the sustainable management of the remaining clam stock. Data reported in this study were collected at monthly intervals and covered varying depths of the Estuary ranging from 0.5 to about 10 m. Overall, a total of 5276 clams with shell lengths ranging from 3.40 to 89.24 mm and total weight from 0.10 to 154.00 g were sampled during the study period. The length-weight relationships were highly significant (p<0.0001) for all the months and the b-values ranged from 2.023 (January 2010) to 3.874 (June 2009). The calculated b-values indicated that clams exhibited different growth patterns at different periods but overall, the pooled samples of 5276 individuals exhibited an isometric growth pattern (b = 3.003). The observed monthly growth patterns exhibited by G. paradoxa appeared to be largely influenced by the reproductive cycle of the organism. During the periods leading to spawning, the clams generally exhibited positive allometric growth patterns (weight increasing faster than length) which appeared to be strongly linked to the build-up of proteins and carbohydrates in their tissues. Successive negative allometric growth patterns (length increasing faster relative to weight) were, however, observed from March to June 2008 and from December 2009 to February 2010, which are possibly indicative of the loss in tissue weight that occurs as a direct result of the spawning process. It will thus be suitable to institute a close season to coincide with the spawning period of the clams to avoid the harvesting of clams during the spawning period which will enhance future recruitment of the clam stock.
PDF Abstract XML References Citation
Received: December 06, 2012;
Accepted: February 12, 2013;
Published: March 16, 2013
How to cite this article
K.A. Obirikorang, D. Adjei-Boateng, H.A. Madkour, S. Amisah and F.A. Otchere, 2013. Length-Weight Relationship of the Freshwater Clam, Galatea paradoxa (Born 1778) from the Volta Estuary, Ghana. Pakistan Journal of Biological Sciences, 16: 185-189.
DOI: 10.3923/pjbs.2013.185.189
URL: https://scialert.net/abstract/?doi=pjbs.2013.185.189
DOI: 10.3923/pjbs.2013.185.189
URL: https://scialert.net/abstract/?doi=pjbs.2013.185.189
INTRODUCTION
Length-weight data are very useful tools as far as fish sampling programs are concerned (Morato et al., 2001) mainly because size is generally considered more biologically relevant than age in fish, as most ecological and physiological factors depend more on size than on age. As noted by Erzini (1994), changes in size have important implications for different aspects of fisheries science and population dynamics. King (2007) noted that the relationships between the lengths and weights of organisms can be used to assess the well-being of individuals within a population and also to determine potential differences between separate unit stocks of the same species. Wilbur and Owen (1964) observed that bivalves are organisms that exhibit relatively large temporal variations in meat content and body weight in response to physiological conditions such as spawning and variation in environmental parameters. These variations in the equilibrium constant usually provide information on the physiological deviations in the condition of bivalves.
The freshwater clam, Galatea paradoxa (Born 1778) is a widely distributed filter-feeding organism in West and Central Africa and usually inhabits the lower reaches of most of the major Rivers extending from the Gulf of Guinea area to the River Congo (Moses, 1990). In Ghana, the main clam bed lies at the portion of the Lower Volta River at Ada Foah, where commercial exploitation takes place mainly for the flesh. The clam fishery of Ghana serves as a viable source of employment and livelihood for the surrounding riparian communities. Studies by Amador (1997) and Agbeko (2010) on the socio-economic status of the clam fishery further revealed that it constitutes a very cheap source of animal protein source to the local communities along the Volta Estuary and beyond and represents a viable option for culture-based fishery in future.
Despite its economic and nutritional importance, there is a dearth of biological information on the G. paradoxa inhabiting the Volta Estuary of Ghana. It is on this basis that this research was conducted to assess the length-weight relationship of G. paradoxa from the Volta Estuary, Ghana. The results of this research will serve as a baseline data for other researchers and aid in the development of stock assessment models for the sustainable management of the threatened clam stock at the Volta Estuary.
MATERIALS AND METHODS
Study area: The study was conducted between Ada and Aveglo, at the Volta Estuary, Ghana, over a 24 month period, from March 2008 to February 2010. Ada (Latitude 05°49' 18.6"N and 000°38.46' 1"E) and Aveglo (05°53 28.2" N and 000°38' 24.7"E) are approximately 15 km apart and represent the the most active clam fishing grounds at the Volta Estuary (Fig. 1).
Data collection: Data reported in this study were collected at monthly intervals for the two-year period and covered the area between Ada and Aveglo at varying depths ranging from 0.5 m to about 10 m. At the shallow portions, samples were collected by wading through the water and hand-picking the clams. At the deeper portions however, samples were obtained from fishermen who employed the Hookah technique in harvesting clams. All size classes harvested from the fishery were included in the sample collection. Samples were transported submerged in river water in insulated chests to the laboratory and processed within 12 h. In the laboratory, individuals were cleansed to remove mud and dried with blotting paper before length and weight measurements were taken. Shell lengths (maximum antero-posterior distance) and total weight (shell weight+flesh weight) were taken using a Powerfix digital caliper (±0.01 mm) and a Sartorious BP 6100 electronic balance (± 0.01 g), respectively. The clams were blotted dry in absorbent paper prior to weighing. The length and weight data collected over the sampling period were used to evaluate the length-weight relationship of G. paradoxa using the non-linear regression equation expressed below by Ricker (1973):
W = aLb |
where, W is the total weight, L is the total length, a is the regression constant (intercept) and b is the regression constant (slope).
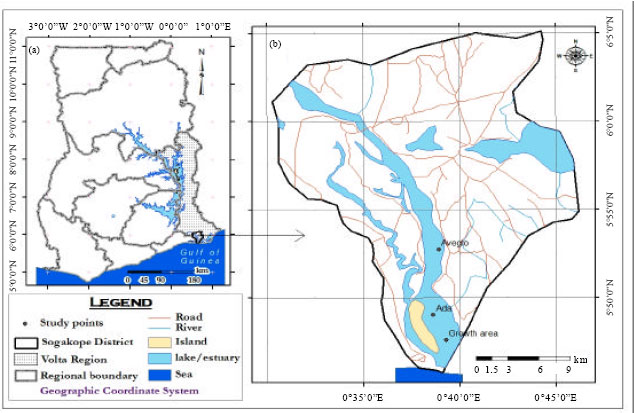 | |
| Fig. 1: | Map showing the clam sampling location between Ada and Aveglo in the Volta Estuary in Ghana |
Statistical analysis: Student’s t-test was applied to verify whether the constant b) was significantly different from 3.0, indicating the type of growth: isometric (b = 3.0), positive allometric (b>3.0) or negative allometric (b<3.0) (Spiegel, 1991). In all cases a significance level of 5% was adopted.
RESULTS AND DISCUSSION
A total of 5276 clams with shell lengths ranging from 3.40 to 89.24 mm and total weight from 0.10 to 154.00 g were measured over the sampling period. Mean length and weight values, length and weight ranges (minimum and maximum) of the clams, the parameters of length-weight relationships (a and b), the Standard Error (SE) of b and the coefficient of determination (r2) for each month are given in Table 1. Length-weight relationships for the separate sexes of G. paradoxa could not be determined because of difficulty in determining sex from any external physical characteristics or colour of the gonads. Length weight relationships were highly significant (p<0.0001) for all the months and the b-values ranged between 2.423 (January 2010) to 3.874 (June 2009). The clams exhibited different growth patterns at different periods of the study.
| Table 1: | W-L relationship parameters of G. paradoxa from the Volta Estuary, Ghana |
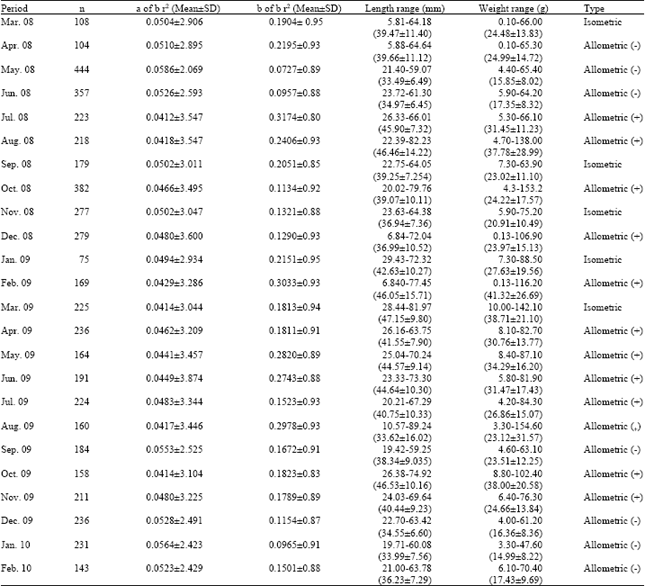 | |
| n: Sample size, SD: Standard deviation, SE: Standard error, a: Regression constant (intercept), b: Regression constant (slope), r2: Coefficient of determination | |
The findings of this study corroborated that of Wilbur and Owen (1964), who stated that, the values of equilibrium constant b lie between 2.4 and 4.5 in most of the bivalves.
Bivalve shell growth and shape are known to be influenced by biotic (endogenous/physiological) and abiotic (exogenous/environmental) factors. A variety of environmental factors are known to influence shell morphology and the relative proportions of many bivalve species (Gaspar et al., 2002). For examples, the type and quality of phytoplankton as a food source of the bivalve, (Alunno-Bruscia et al., 2001; Kovitvadhi et al., 2006), water quality (Buddensiek, 1995; Lajtner et al., 2004), water depth (Claxton et al., 1998; Karayucel and Karayucel, 2000; Lajtner et al., 2004), currents (Blay, 1989; Fuiman et al., 1999), water turbulence (Hinch and Bailey, 1988), type of sediment (Newell and Hidu, 1982; Lajtner et al., 2004), type of bottom (Claxton et al., 1998) and wave exposure (Akester and Martel, 2000).
The monthly growth patterns exhibited by G. paradoxa appeared to be largely influenced reproductive cycle of the organism. Studies by Etim et al. (1991) and Adjei-Boateng et al. (2011) revealed that spawning in G. paradoxa starts in June each year and is completed between October and November. This study observed that during these periods the clam growth patterns were generally isometric (growth with unchanged body proportions and specific gravity) and positive allometric (weight increasing faster than length). The allometric growth trends observed during the spawning period appear to be strongly linked to the build-up of proteins and carbohydrates in the tissues of the clams. Prior to and during the spawning months, proteins and carbohydrates are accumulated in the clam tissues for gonad tissue production, energetic storage and consumption (LaTouche and Mix, 1982; Paez-Osuna et al., 1995) leading to an increase in the tissue weight of the clam and consequently an increase in total weight relative to shell length. The successive negative allometric growth patterns (length increasing faster relative to weight) observed from March to June 2008 and from December 2009 to February 2010 are possibly indicative of the loss in tissue weight that occurs as a direct result of the spawning process. Galstoff (1964) and Etim et al. (1991) observed that the ripe clam gonad may comprise 31 to 41% of the total body weight and thus, when shed during spawning period accounts for an appreciable weight loss.
Despite the different growth patterns exhibited by the clams during different stages of the study, the overall combined samples (5276 individuals) exhibited an isometric growth pattern (b = 3.003) over the two-year period (Fig. 2).
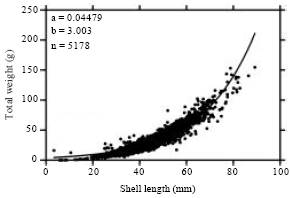 | |
| Fig. 2: | Length-weight relationship of G. paradoxa from the volta estuary, Ghana over a two-year period (March, 2008-February, 2010) |
This result confirms the findings of a study by Ofori-Danson and Amoah (2007) on the length weight relationship of Galatea paradoxa at the same location, which revealed that the bivalve in its natural habitat exhibits an isometric growth pattern (b = 2.9129).
CONCLUSION
The study revealed that the freshwater clams Galatea paradoxa exhibited different growth patterns at different periods of the study, which appeared to be largely influenced by their reproductive cycle. Overall, however, the pooled samples over the 24 month period exhibited an isometric growth pattern. The results of this study appeared to support earlier findings that spawning in G. paradoxa starts in June each year and is completed between October and November each year, thus making it imperative to institute a close season that coincides with this period to enhance stock recruitment in future.
ACKNOWLEDGEMENT
The authors are grateful to the Department of Fisheries and Watershed Management (Faculty of Renewable Natural Resources) of the Kwame Nkrumah University of Science and Technology (KNUST), Kumasi for the logistical support for this research.
REFERENCES
- Adjei-Boateng, D., K.A. Obirikorang, S. Amisah, H.A. Madkour and F.A. Otchere, 2011. Relationship between gonad maturation and heavy metal accumulation in the clam, Galatea paradoxa (Born 1778) from the Volta estuary, Ghana. Bull. Environ. Contam. Toxicol., 87: 626-632.
CrossRefPubMedDirect Link - Akester, R.J. and A.L. Martel, 2000. Shell shape, dysodont tooth morphology and hinge-ligament thickness in the bay mussel Mytilus trossulus correlate with wave exposure. Can. J. Zool., 78: 240-253.
CrossRef - Alunno-Bruscia, M., E. Bourget and M. Frechette, 2001. Shell allometry and length-mass-density relationship for Mytilus edulis in an experimental food-regulated situation. Mar. Ecol. Prog. Ser., 219: 177-188.
Direct Link - Buddensiek, V., 1995. The culture of juvenile freshwater pearl mussels Margaritifera margaritifera L. in cages: A contribution to conservation programmes and the knowledge of habitat requirements. Biol. Conserv., 74: 33-40.
CrossRef - Claxton, W.T., A.B. Wilson, G.L. Mackie and E.G. Boulding, 1998. A genetic and morphological comparison of shallow- and deep-water populations of the introduced dreissenid bivalve Dreissena bugensis. Can. J. Zool., 76: 1269-1276.
CrossRef - Hinch, S.G. and R.C. Bailey, 1988. Within- and among-lake variation in shell morphology of the freshwater clam Elliptio complanata (Bivalvia: Unionidae) from south-central Ontario lakes. Hydrobiologia, 157: 27-32.
CrossRef - Erzini, K., 1994. An empirical study of variability in length-at-age of marine fishes. J. Applied Ichthyol., 10: 17-41.
CrossRefDirect Link - Etim, L., E.R. Akpan and P. Muller, 1991. Temporal trends in heavy metal concentrations in the clam Egeria radiata (Bivalvia: Tellinacea: Donacidae) from the Cross River, Nigeria. Rev. Hydrobiol. Trop., 24: 327-333.
Direct Link - Fuiman, L.A., J.D. Gage and P.A. Lamont, 1999. Shell morphometry of the deep sea protobranch bivalve Ledella pustulosa in the Rockall Trough, north-east Atlantic. J. Mar. Biol. Assoc. UK., 79: 661-671.
Direct Link - Gaspar, M.B., M.N. Santos, P. Vasconcelos and C.C. Monteiro, 2002. Shell morphometric relationships of the most common bivalve species (Mollusca: Bivalvia) of the Algarve coast (southern Portugal). Hydrobiologia, 477: 73-80.
CrossRef - Karayucel, S. and I. Karayucel, 2000. The effect of environmental factors, depth and position on the growth and mortality of raft-cultured blue mussels (Mytilus edulis L.). Aquac. Res., 31: 893-899.
CrossRef - Kovitvadhi, S., U. Kovitvadhi, P. Sawangwong, A. Thongpan and J. Machado, 2006. Optimization of diet and culture environment for larvae and juvenile freshwater pearl mussels, Hyriopsis (Limnoscapha) myersiana Lea, 1856. Invert Reprod. Dev., 49: 61-70.
Direct Link - Lajtner, J., Z. Marusic, G.I.V. Klobucar, I. Maguire and R. Erben, 2004. Comparative shell morphology of the zebra mussel, Dreissena polymorpha in the Drava river (Croatia). Biologia, 59: 595-600.
Direct Link - LaTouche, Y.D. and M.C. Mix, 1982. The effects of depuration, size and sex on trace metal levels in bay mussels. Mar. Pollut. Bull., 13: 27-29.
CrossRef - Morato, T., P. Afonso, P. Lourinho, J.P. Barreiros, R.S. Santos and R.D.M. Nash, 2001. Length-weight relationships for 21 coastal fish species of the Azores, North-Eastern Atlantic. Fish. Res., 50: 297-302.
CrossRefDirect Link - Moses, B.S., 1990. Growth, biomass, mortality, production and potential yield of the West African clam, Egeria radiata (Lamarck) (Lamellibranchia, Donacidae) in the Cross River system, Nigeria. Hydrobiologia, 196: 1-15.
Direct Link - Newell, C.R. and H. Hidu, 1982. The effects of sediment type on growth rate and shell allometry in the soft shelled clam Mya arenaria (L.). J. Exp. Mar. Biol. Ecol., 65: 285-295.
CrossRef - Paez-Osuna, P., M.G. Frias-Espericueta and J.I. Osuna-Lopez, 1995. Trace metal concentrations in relation to season and gonadal maturation in the oyster Crassostrea iridescens. Mar. Environ. Res., 40: 19-31.
CrossRef - Ricker, W.E., 1973. Linear regression in fisheries research. J. Fish. Res. Board Can., 30: 409-434.
Direct Link