
Berhanu D. Bekele
Department of Plant Biotechnology, University of Agricultural Sciences, GKVK, Bangalore-560065, India
G.K. Naveen
Department of Plant Biotechnology, University of Agricultural Sciences, GKVK, Bangalore-560065, India
S. Rakhi
Department of Plant Biotechnology, University of Agricultural Sciences, GKVK, Bangalore-560065, India
H.E. Shashidhar
Department of Plant Biotechnology, University of Agricultural Sciences, GKVK, Bangalore-560065, India
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 23 | Page No.: 1714-1721







ABSTRACT
The objectives of the present study were to evaluate genetic variability parameters, correlations that exist for grain Zn concentration and yield related traits and identification of SSR markers linked to these traits in rice. One hundred seventy six Recombinant Inbred Lines (RILs) of Azucena X Moromutant were grown at University of Agricultural Sciences, Bangalore in augmented experimental design during wet seasons of 2010 and 2011. The study revealed significant genetic variability for all the traits. Grain yield per plant and grain zinc concentration showed higher phenotypic and genotypic co-efficient of variation. Significant positive correlation was observed for grain yield per plant with number of productive tillers per plant (r = 0.5) and number of tillers per plant (r = 0.4). Grain zinc concentration showed negative correlation with grain yield per plant (r = - 0.27). The path-coefficient analysis indicated the positive direct effect of number of productive tillers per plant on grain yield per plant (0.514). Grain zinc concentration showed negative direct effect on grain yield per plant (-0.186). Single-marker analysis using 26 SSR markers on RILs mapping population showed that RM212, RM263, RM6832, RM152, RM21, RM234 and RM3331 had association with grain zinc concentration and other yield related traits. But validation of these markers on fifty two rice genotypes showed that only three markers RM263, RM152 and RM21 had association with grain zinc concentration. Therefore, the genetic information generated and molecular markers identified from this study could be used for zinc biofortification programmes in rice.
PDF Abstract XML References Citation
Received: January 13, 2013;
Accepted: March 02, 2013;
Published: May 16, 2013
How to cite this article
Berhanu D. Bekele, G.K. Naveen, S. Rakhi and H.E. Shashidhar, 2013. Genetic Evaluation of Recombinant Inbred Lines of Rice (Oryza sativa L.) for Grain Zinc Concentrations, Yield Related Traits and Identification of Associated SSR Markers. Pakistan Journal of Biological Sciences, 16: 1714-1721.
DOI: 10.3923/pjbs.2013.1714.1721
URL: https://scialert.net/abstract/?doi=pjbs.2013.1714.1721
DOI: 10.3923/pjbs.2013.1714.1721
URL: https://scialert.net/abstract/?doi=pjbs.2013.1714.1721
INTRODUCTION
Rice is the major stable food for billions of people in Asia, Africa and Latin America. It is estimated that the total global area of rice is 150.8 million ha. Asia is a major producer and consumer of rice which accounts for 90 to 92% of world’s rice area and productivity, respectively. Rice is a major source of dietary energy, protein, vitamins and essential fatty acids and micronutrients for the poor and vulnerable society that relied upon it as a staple food.
Micronutrient deficiency (hidden hunger) is a major health problem which affects more than half of the world’s population and contributing to high rate of children and women's mortality (Pfeiffer and McClafferty, 2007). According to World Health Organization (2002), Zn deficiency is one of most important health problems for resource poor people who relied upon cereals as a staple food. Zn deficiency induces human health problems such as impairments in physical development, immune system, brain function, pneumonia, weight loss, growth retardation and delayed puberty in adolescents, poor appetite, delayed wound healing, etc (WHO, 1996). To address this issue, Zn food fortification and supplementation approaches are recommended but these methods are expensive and are not easily accessible for resource people living in developing countries (Bouis et al., 2003; Pfeiffer and McClafferty, 2007). Therefore, an alternative approach called Zn biofortication has been devised to reach resource poor rural society to enhance its content in staple food crops (Cakmak, 2008).
There is a large diversity available in rice germplasm banks and farmers fields in remote areas in Asia, Africa and other countries across the world. International Rice Research Institute (IRRI) is leading rice research institute in the world which consists of a large rice germplasm collections. Screening of 1000 rice genotypes for grain Zn concentration showed the presence of higher genetic variability for the trait that ranges from 15.3 to 58.4 ppm (Graham et al. 1999). Nagarathna et al. (2010) also reported the presence of wide genetic variability for rice grain Zn concentration ranges from 8.4 to 50.0 ppm.
Most research in plant breeding was focused on breeding for high yields, resistance to biotic factors and tolerance to abiotic stresses. Recently, the trend has changed to incorporate desired quality parameters to the grains from health and economic perspectives. There is also large genetic variation for grain Zn concentration in rice germplam accessions and this variation can be exploited in breeding programs (Welch and Graham, 2004). As a trait, grain Zn concentration is a complex trait governed by a number of genes/QTLs located on different chromosomes with different phenotypic effects (Avendano, 2000; Biradar et al., 2007; Lu et al., 2008; Garcia-Oliveira et al., 2009; Banerjee et al., 2010; Zhang et al., 2011). Characterization and identification of this complex genetic makeup of the trait has been facilitated by molecular markers. Various researchers have identified a number of QTLs for rice grain Zn concentration using different genetic materials. Avendano (2000) identified a locus for Zn concentrations in grains of rice on chromosome 5 located between OSR35 and RM267 in a study with RILs derived from Modhukar and IR26. Biradar et al. (2007) identified six loci associated with Zn concentration on chromosome 1, 4, 5, 8, 9 and 11 using 93 double haploid mapping populations obtained from IR64 X Azucena. Stangoulis et al. (2007) identified two QTLs qZN-1 and qZN-12 for grain Zn concentration with phenotypic variations of 15 and 13%, respectively. Lu et al. (2008) investigated three loci qZN-5, qZN-7 and qZN-11 for Zn concentration on chromosomes 5, 7 and 11. Garcia-Oliveira et al. (2009) also obtained three QTLs qZN-5, qZN-8 and qZn-12 for grain Zn concentration using 179 SSR markers. So far, there is no report indicating tight linkage between genes associated with Zn concentrations in the grains and molecular markers yet to be established. Marker Assisted Selection (MAS) can be applied once desired marker become available. The objective of this study is to evaluate genetic parameters, correlations that exist for grain Zn concentration and grain yield related traits among Recombinant Inbred Lines (RILs) derived from Azucena X Moromutant and identification of SSR markers linked to the trait.
MATERIALS AND METHODS
Experimental site and design: The experiment was carried out during wet seasons of 2010 and 2011 at Department of Plant Biotechnology, University of Agricultural Sciences, GKVK campus, Bangalore, India. The experiment was laid out in augmented experimental design (Federer, 1961) with 18 blocks which consisted of one hundred seventy six RILs derived from a cross between Azucena and Moromutant and two check varieties (Azucena and IR64). The same experimental design was also followed for fifty two rice genotypes used for marker validation studies. Seven rows of individual genotypes were direct sown in a plot size of two meters length with a spacing of 30 cm between the rows and 15 cm between the plants. All the experimental plants were raised by direct seeding in the main field under aerobic condition. The field management was followed the recommended packages of agronomic procedures of UAS, Bangalore. Soil was fertilized with 5 t ha-1 of farm yard manure and NPK in the ratio of 100: 50: 50 kg ha-1. N was provided in the form of urea at basal, 30 and 60 days after sowing of 50, 25 and 25%, respectively. P was provided as single super phosphate (16% P2O5) and K as murite of potash (60% K2O). Plants were grown in available rainfall conditions but irrigation was done once in five days in absence of rainfall for all the plots. The data were recorded from middle rows for days to 50% flowering, days to maturity, plant height (cm), number of tillers per plant, number of productive tillers per plant and grain yield per plant (g) at respective stages of the crop. 100 grains weight (g), grain length (mm), grain breadth (mm) and grain Zn concentrations (ppm) were recorded after harvest.
Sample preparation and estimation of Zn concentrations: Seeds of RILs were harvested manually and hand threshed to avoid any contamination. The seeds were dehusked using clean pestle and mortar. The grains were washed quickly with double distilled water to remove any surface contaminants and dried in hot air oven at 70°C for 72 h. Five milliliter of 70% concentrated nitric acid (Merck) was added to 0.5 g powdered grain samples and the mixture was incubated overnight. In the next day, 5 mL of diacid mixture [Nitric acid: Perchloric acid (Merck) in the ratio of 10: 4] was added to the incubated sample. The mixture was then placed on the electric hot plate, allowing the evaporation of acid in the form of white fumes, until white precipitate was formed at the base of the flask. The digested sample was left to cool for an hour and the volume was made up to 50 mL using double-distilled water. Calibration of an instrument for estimation of an element was done using five standard solutions with appropriate dilutions. A suitable blank was run simultaneously to account for contamination from the reagents. Zn concentrations in the grain samples of rice were estimated using AAS (Perkin Elmeyer Precisely analyst 400 AAS) at 213.86 nm at quality control laboratory, ANGRU, Hydrabad.
Genotyping of RILs with SSR markers: DNA extraction for RILs, parental lines and rice genotypes were done using CTAB extraction method from 21 days young leaves tissues as the protocol described by Doyle and Doyle (1990). A total of seventy two SSR markers were screened for the existence of polymorphism for the parental genotypes (Azucena and Moromutant). Among these, twenty six SSR markers were found polymorphic in parental genotypes. These polymorphic SSR markers were used to screen 176 RILs mapping population.
Statistical analysis: The analysis of variance for all the traits was computed with “AUGMENT 1” computer programme (Agrawal and Sapra, 1995).Genetic parameters, correlation studies and path-coefficient analysis were analyzed with GENRES statistical software package (GENRES, 1994). The associations of different traits with molecular markers were analyzed with single marker analysis and regression analysis with SPSS16.0 (SPSS Inc.).
RESULTS
Analysis of variance: In the present study, analysis of variance revealed highly significant differences existed among the RILs for all the characters in a study in wet season 2010 and 2011.
Genetic variability for grain Zn concentration and yield related traits: In wet season of 2010, grain Zn concentration was 16.1 ppm (AM99) to 88.6 ppm (AM150) with an average value of 31.24 ppm whereas, in wet season of 2011, the range of variation was 22.70 ppm (AM132) to 58.80 (AM103) with an average value of 33.03 ppm. But in parental lines the mean Zn concentration in the grains of Azucena and Moromutant was 37.61 and 26.30, respectively in both years. Grain yield per plant in wet season 2010, the range of variation was 0.27 (AM164) to 22.63 (AM169) with an average value of 6.42 g whereas, in wet season 2011, the range of variation was 0.60 g (AM10 and AM68) to 28.80 g (AM7) with an average value of 9.73 g as shown on Table 1. In parental lines the mean yield per plant of Azucena and Moromutant was 4.10 and 14.28 g, respectively in both years.
Variability parameters of Genotypic co-efficient of Variation (GCV), phenotypic co-efficient of variation (PCV), broad sense heritability (h2) and genetic advance as percent of mean (GAM) in respect of all the characters in RILs grown in wet season 2010 and 2011 are shown in Table 1. In the present study, PCV and GCV were highest for number of tillers per plant (PCV = 34.21, GCV = 30.44), number of productive tillers per plant (PCV = 37.28, GCV = 31.84), grain yield per plant (PCV = 67.76, GCV = 63.70) and grain Zn concentration (PCV = 28.16, GCV = 24.91). Moderate values (10-20 %) of PCV and GCV observed for plant height, 100 grains weight and grain breadth. Low values (<10 %) of PCV and GCV were recorded for days to maturity (PCV = 4.79, GCV = 4.03) and grain length (PCV = 8.29, GCV = 8.14).
High heritability coupled with high genetic advance as percent mean were recorded for plant height (h2 =87.83, GAM = 35.34), number of tillers per plant (h2 = 79.18, GAM = 55. 80), number of productive tillers per plant (h2 = 72.95, GAM = 56.03), grain yield per plant (h2 = 90.96, GAM = 116.31), grain Zn concentration (h2 = 80.59, GAM = 66.64) and 100 grains weight (h2 = 90.86, GAM = 31.84).
| Table 1: | Estimates of genetic parameters for different traits in RILs |
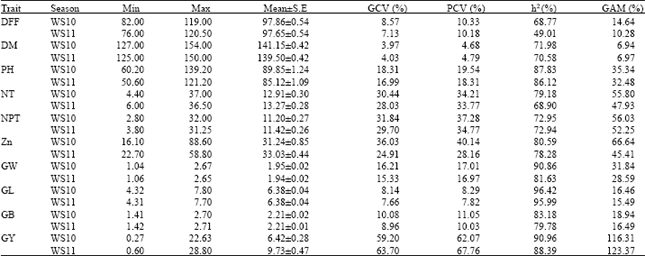 | |
| DFF: Days to 50% flowering, DM: Days to maturity, PH: Plant height (cm), NT: Number of tillers per plant, NPT: Number of productive tillers per plant, Zn: Grain Zn concentrations (ppm), GW: 100 grains weight (g), GL: Grain length (mm), GB: Grain breadth (mm), GY: Grain yield per plant (g), WS10: wet season 2010, WS11: wet season 2011, GCV: Genotypic coefficients of variation, PCV: Phenotypic coefficients of variation, h2: Heritability in broad sense and GAM: Genetic advance as percent mean | |
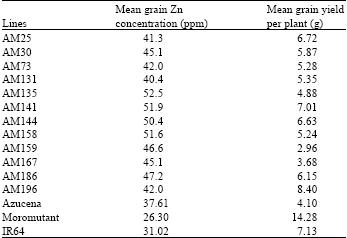
| Table 2: | Transgressant lines selected from RILs based grain zinc concentration |
 | |
High heritability coupled with moderate genetic advance as percent mean (11-20 %) was recorded for days to 50% flowering, grain length and grain breadth. High heritability coupled with low genetic advance as percent mean (0-10 %) was recorded for days to maturity (h2 = 71.98, GAM = 6.94) (Table 1).
It has also been observed that some RILs AM25, AM30, AM73, AM131, AM135, AM141, AM144, AM158, AM159, AM167, AM186 and AM196 were consistent for higher grain Zn accumulation with values ranges from 26.30 ppm to 52.5 ppm in both seasons with relatively better grain yield (Table 2).
Correlation studies for grain Zn concentration and yield related traits: Highly significant and positive correlation were observed for number of productive tillers per plant with number of tillers per plant (r = 0.97), grain yield per plant with number of productive tillers per plant (r = 0.5) and number of tillers per plant (r = 0.4). Grain Zn concentration showed positive correlation with 100 grains weight (r = 0.15) and grain length (r = 0.19) whereas, grain Zn concentration showed negative correlation with grain yield per plant (r = -0.24) as shown on Table 3. Assessment of the relationship between grain Zn concentration and grain yield per plant using linear regression showed that there was a negative correlation between these traits (Fig. 1a, b).
Path-coefficient analysis: The path-coefficient analysis indicated that high positive direct effect of number of productive tillers per plant on grain yield per plant (0.514, 1.249). Grain Zn concentration showed negative direct effect on grain yield per plant (-0.184, -0.143) as shown on Table 4, 5. Among indirect effects, number of tillers per plant had the highest positive indirect effect on grain yield per plant via number of productive tillers per plant (0.497, 1.186) whereas, number of productive tillers per plant had the highest negative indirect effect on grain yield via number of tillers plant (-0.396, -0.755) in both seasons.
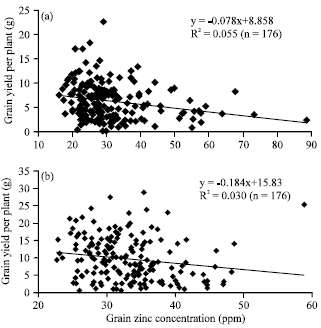 | |
| Fig. 1(a-b): | Relationship between grain Zn concentration and grain yield per plant in RILs (a) Grown in wet season 2010 and (b) Grown in wet season 2011. n: number of recombinant inbred lines |
Grain Zn concentration showed negative to low indirect effect via other characters for grain yield per plant as shown on Table 4 and 5.
Identification and validation of SSR markers: Among twenty six SSR markers only seven markers RM212, RM263, RM6832, RM152, RM21, RM234 and RM3331 showed association for grain Zn concentration by single-marker analysis explaining a maximum phenotypic variation of 6.50, 8.60, 8.20, 6.10, 11.70, 7.30 and 6.10%, respectively (Table 6). But validation of these seven SSR markers on different genetic background of fifty two rice genotypes showed that only three markers, RM263, RM152 and RM21 showed an association with grain Zn concentration with phenotypic variations of 10.7, 10.4 and 13.7 %, respectively.
DISCUSSION
The RILs are the product of hybridization and forwarded to F8 generation without any selection with a single seed descent method to generate transgressive segregants for grain Zn concentration. Cultivation of rice in aerobic condition is a novel approach developed by crossing of lowland varieties with upland varieties to grow in rainfed, under non-flooded environments (Bouman et al., 2007; Shashidhar, 2007). In the present study, parental line Azucena is upland japonica variety whereas, Moromutant (mutant of Moroberekan) is a lowland variety.
| Table 3: | Phenotypic correlations coefficients for different traits in RILs (n = 176) |
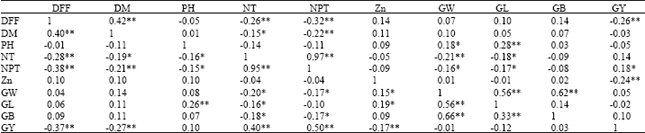 | |
| *Significant at 5%, **significant at 1%, Above half diagonal values on the table indicates correlations coefficients in wet season 2010, Below half diagonal values on the table indicates correlations coefficients in wet season 2011, Note: DFF: Days to 50 % flowering; DM: Days to maturity; PH: Plant height (cm); NT: Number of tillers per plant; NPT: Number of productive tillers per plant; Zn: Grain Zn concentrations (ppm); GW: 100 grains weight (g); GL: Grain length, (mm); GB: Grain breadth (mm); GY: Grain yield per plant (g) | |
| Table 4: | Path-coefficient analysis for different traits of RILs in wet season 2010 (n = 176) |
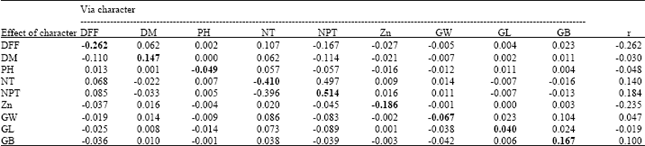 | |
| Diagonals bold indicates direct effects | |
| Table 5: | Path-coefficient analysis for different traits of RILs in wet season 2011 (n = 176) |
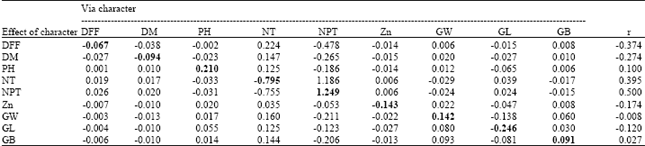 | |
| Diagonals bold indicates direct effects, Note: DFF: Days to 50 % flowering; DM: Days to maturity; PH: Plant height (cm); NT: Number of tillers per plant; NPT: Number of productive tillers per plant; Zn: Grain Zn content (ppm); GY: Grain yield per plant (g); GW: 100 grains weight (g); GL: Grain length (mm); GB: Grain breadth (mm) | |
The RILs obtained from these parental lines exhibited variability for grain yield per plant adapted to aerobic condition. RILs underwent continuous recombination during the course of stabilization, thus incorporate favourable quantitative and qualitative traits of both the parents in its progenies. In this study, twelve best stable transgressive segregants for grain Zn concentration (>40 ppm) were obtained in both seasons (Table 2). These lines have potential for Zn biofortification programmes in rice. The Zn concentration showed significant transgressive segregations with values either larger or smaller than those of the parents. Superior transgresant progenies would likely complement important favourable traits lacking by the parental genotypes. In addition to their capacity for high grain zinc concentration, they are also able to adapt to aerobic conditions.
The high PCV and GCV estimates were reported by Samak et al. (2011) for grain Zn concentrations; Bisne et al. (2009), Akinwale et al. (2011) for grain yield per plant; Chauhan (1996), Elayaraja et al. (2005), Bisne et al. (2009) for number of tillers per plant and number of productive tillers per plant. Similar results of high PCV and GCV (>20%) were obtained in RILs of Azucena X Moromutant in both seasons suggested that these characters were more influenced by genetic factors than environmental factors. Hence, further trait improvement for these could be practiced by simple selection.
| Table 6: | Single-marker analysis (SMA) for SSR markers in RILs mapping population of rice for grain Zn concentration and yield related traits in wet season 2010 and 2011 |
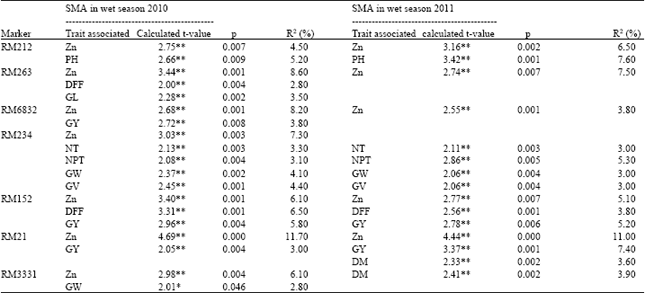 | |
| *’** are significant at 5 and 1%, respectively | |
Moderate coefficient values of PCV and GCV (10-20%) for other traits indicated considerable amount of variability whereas, low values of PCV and GCV (<10%) indicated lower amount of variability for the character.
High heritability (>60%) coupled with high genetic advance as per cent mean (>20%) were revealed for most of the characters indicating the presence of considerable variation and additive gene effects. Hence, trait improvement in breeding could be effectively done through phenotypic selection. High heritability coupled with moderate genetic advance as percent mean (11-20 %) suggests the traits were primarily under genetic control and selection for these traits can be achieved through their phenotypic performance whereas, high heritability coupled with low genetic advance as percent mean (0-10%) indicated lower proportion of genetic components in the total variability and suggested that this character was controlled by non-additive gene components.
Highly significant and positive correlation was earlier reported for grain Zn concentration with 100 grain weight by Tiwari et al. (2009), grain yield per plant with number of productive tillers per plant by Akinwale et al. (2011). The high correlations existing between different plant traits can be used to conduct indirect selection of a desired complex trait in a breeding program. On the other hand, negative correlations exhibited for grain Zn content and grain yield per plant indicated the absence of association for these traits. Thus, selection for enhancement of these traits could be executed separately but simultaneously. Similar reports on negative correlation between grain Zn concentration and grain yield per plant were revealed by Shi et al. (2008), Nagarathna et al. (2010) and Nagesh et al. (2012).
Path-coefficient analysis reveals the direct and indirect effects of different traits on grain yield per plant. Suman et al. (2006), Panwar et al. (2007) and Nagesh et al. (2012) earlier reported high positive direct effect of number of productive tillers per plant on grain yield per plant. Similar results were obtained in our study in both seasons. Chakraborti et al. (2009) reported the absence of the direct and indirect effects on grain Zn concentration on yield in maize. Grain Zn concentration showed negative direct effect on grain yield per plant in wet seasons of 2010 and 2011. Nagesh et al. (2012) reported low direct effect of grain Zn concentration towards grain yield. This indicated that grain Zn concentration had no contribution towards grain yield. Thus, grain Zn concentration cannot be used as criteria for enhancement of grain yield per plant. Selection based on number of productive tillers per plant for grain yield per plant would be most effective strategy.
Zn accumulation in the grains of rice is a complex trait which involves a number of Quantitative Trait Loci (QTLs) distributed to different regions of the chromosome (Grusak and Dellapenna, 1999; Avendano, 2000; Lu et al., 2008; Garcia-Oliveira et al., 2009; Zhang et al., 2011). Hence, QTLs responsible for this trait can be easily identified with closely linked molecular markers. So far, there is no report indicating tight linkage of a marker to grain Zn concentration in rice. To facilitate tagging of the markers to this trait, preliminary steps are required to characterize molecular markers linked to loci responsible for Zn accumulation in the grains of rice. In single-marker analysis, each marker is assumed as a potential QTL and the marker should show phenotypic variation on the trait. In RILs, a total of fourteen alleles were identified with seven polymorphic SSR markers associated with grain Zn content. It has been found that RM212, located on chromosome one was closely associated with grain Zn concentration and plant height. In earlier report, Stangoulis et al. (2007) identified a QTL flanking to this marker for Zn concentration on chromosomes one explaining 15% of the total phenotypic variation in rice. RM263 located on chromosome two was linked to grain Zn concentration, days to 50% flowering and grain length. This marker was earlier reported by Thanh et al. (2006) to flank a QTL responsible for root weight to shoot ratio in rice. RM6832 located on chromosome three linked to grain Zn concentration and grain yield. There are a number of reports showing the high correlations that exist between grain Zn and Fe concentration (Shi et al., 2008; Kumar et al., 2008; Tiwari et al., 2009; Anandan et al., 2011; Nagesh et al., 2012). These results also confirmed the earlier reports. RM152 located on chromosome eight was linked to grain Zn concentration, days to 50% flowering, days to maturity and grain yield. This marker was earlier reported by Garcia-Oliveira et al. (2009) for its association with grain Zn concentration accounted for the largest proportion of phenotypic variation for QTL located on chromosome 8 in rice. RM21 located on chromosome five was linked with grain Zn concentration, grain yield, days to maturity and harvest index. Avendano (2000), Lu et al. (2008) and Garcia-Oliveira et al. (2009) identified a QTL for Zn concentration on chromosomes five. Validation of putative marker is used to confirm its reproducibility for utility in marker assisted selection. In the present work, seven SSR markers identified for their association with grain Zn concentrations in RILs mapping populations were further validated on different genetic background of fifty two rice genotypes showed that only three markers, RM263, RM152 and RM21 showed association with grain Zn concentration. Thus, these markers could be used for marker-assisted selection for Zn biofortification programmes with further scrutiny.
CONCLUSION
The present study revealed that there is higher genetic variability for grain Zn concentration among RILs mapping population and superior transgressant lines promising for trait improvement are obtained. Correlation studies showed that grain Zn concentration had negative correlation with grain yield per plant. Thus, breeding for high grain Zn concentration in the grains and enhancement of grain yield per plant could be executed separately and traits can be introgressed into a single plant. Molecular analysis showed that three SSR markers RM263, RM21 and RM152 are associated with grain Zn concentration in the grains of rice explaining 10.7, 10.4 and 13.7 % phenotypic variation, respectively. These markers can be used for rice grain Zn biofortification programmes with further detail characterization.
ACKNOWLEDGMENT
The authors are thankful to the Department of Biotechnology, New Delhi, India for the financial support (grant number Ab/Ac 8270); Indian Council for Cultural Relations and Ethiopian government for providing fellowship to the first author; and University of Agricultural Sciences, GKVK, Bangalore for providing facilities to accomplish this study.
REFERENCES
- Agrawal, R.C. and R.L. Sapra, 1995. AUGMENT 1: A microcomputer based program to analyze augmented randomized complete block design. Indian J. Plant Genet. Res., 8: 61-69.
Direct Link - Akinwale, M.G., G. Gregorio, F. Nwilene, B.O. Akinyele, S.A. Ogunbayo and A.C. Odiyi, 2011. Heritability and correlation coefficient analysis for yield and its components in rice (Oryza sativa L.). Afr. J. Plant Sci., 5: 207-212.
Direct Link - Anandan, A., G. Rajiv, R. Eswaran and M. Prakash, 2011. Genotypic variation and relationships between quality traits and trace elements in traditional and improved rice (Oryza sativa L.) genotypes. J. Food Sci., 76: H122-H130.
CrossRefDirect Link - Biradar, H., M.V. Bhargavi, R. Sasalwad, R. Parama and S. Hittalmani, 2007. Identification of QTL associated with silicon and Zn content in rice (Oryza sativa L.) and their role in blast disease resistance. Indian J. Genet., 67: 105-109.
Direct Link - Bisne, R., A.K. Sarawgi and S.B. Verulkar, 2009. Study of heritability genetic advance and variability for yield contributing characters in rice. Bangladesh J. Agril. Res., 34: 175-179.
CrossRef - Bouis, H.E., B.M. Chassy and J.O. Ochanda, 2003. Genetically modified food crops and their contribution to human nutrition and food quality. Trends Food Sci. Technol., 14: 191-209.
CrossRefDirect Link - Cakmak, I., 2008. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil, 302: 1-17.
CrossRefDirect Link - Banerjee, S., D.J. Sharma, S.B. Verulkar and G. Chandel, 2010. Use of In-silico and semiquantitative RT-PCR approaches to develop nutrient rich rice (Oryza sativa L.). Indian J. Biotechnol., 9: 203-212.
Direct Link - Garcia-Oliveira, A.L., L. Tan, Y. Fu and C. Sun, 2009. Genetic identification of quantitative trait loci for contents of mineral nutrients in rice grain. J. Integr. Plant Biol., 51: 84-92.
CrossRefDirect Link - Graham, R., D. Senadhira, S. Beebe, C. Iglesias and I. Monasterio, 1999. Breeding for micronutrient density in edible portions of staple food crops: Conventional approaches. Field Crops Res., 60: 57-80.
CrossRefDirect Link - Grusak, M.A. and D. Dellapenna, 1999. The nutrient composition of plants to enhance human nutrition and health. Annu. Rev. Plant Physiol. Plant Mol. Biol., 50: 133-161.
Direct Link - Kumar, A.A., B.V.S. Reddy, B. Ramaiah, P.S. Reddy, K.L. Sahrawat and H.D. Upadhyaya, 2009. Genetic variability and plant character association of grain Fe and Zn in selected core collection accessions of sorghum germplasm and breeding lines. E-J. SAT Agric. Res., 7: 1-4.
Direct Link - Lu, K., L. Li, X. Zheng, Z. Zhang, T. Mou and Z. Hu, 2008. Quantitative trait loci controlling Cu, Ca, Zn, Mn and Fe content in rice grains. J. Genet., 87: 305-310.
CrossRefDirect Link - Nagarathna, T.K., A.G. Shankar and M. Udayakumar, 2010. Assessment of genetic variation in Zn acquisition and transport to seed in diversified germplasm lines of rice (Oryza sativa L.). J. Agric. Tech., 6: 171-178.
Direct Link - Nagesh, V. Ravindrababu, G. Usharani and T.D. Reddy, 2012. Grain iron and Zn association studies in rice (Oryza sativa L.) F1 progenies. Arch. Applied Sci. Res., 4: 696-702.
Direct Link - Pfeiffer, W.H. and B. McClafferty, 2007. HarvestPlus: Breeding crops for better nutrition. Crop Sci., 47: S88-S105.
CrossRefDirect Link - Samak, N.R.A., S. Hittalmani, N. Shashidhar and H. Biradar, 2011. Exploratory studies on genetic variability and genetic control for protein and micronutrient content in F4 and F5 generation of rice (Oryza sativa L.). Asian J. Plant Sci., 10: 376-379.
CrossRefDirect Link - Shi, R., H. Li, Y. Tong, R. Jing, F. Zhang and C. Zou, 2008. Identification of quantitative trait locus of zinc and phosphorus density in wheat (Triticum aestivum L.) grain. Plant Soil, 306: 95-104.
CrossRefDirect Link - Stangoulis, J.C.R., B.L. Huynh, R.M. Welch, E.Y. Choi and R.D. Graham, 2007. Quantitative trait loci for phytate in rice grain and their relationship with grain micronutrient content. Euphytica, 154: 289-294.
CrossRefDirect Link - Thanh, N.D., N.T.K. Lien, P.Q. Chung, T.Q. Trong, L.T.B. Thuy and H. Nguyen, 2006. Mapping QTLs associated with root traits related to drought resistance in vietnamese upland rice. Asean J. Sci. Tech. Dev., 23: 323-332.
Direct Link - Tiwari, V.K., N. Rawat, P. Chhuneja, K. Neelam and R. Aggarwal et al., 2009. Mapping of quantitative trait loci for grain iron and Zinc concentration in Diploid A genome wheat. J. Hered., 100: 771-776.
CrossRefDirect Link - Welch, R.M. and R.D. Graham, 2004. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot., 55: 353-364.
CrossRefDirect Link - Zhang, X., G. Zhang, L. Guo, H. Wang and D. Zeng et al., 2011. Identification of quantitative trait loci for Cd and Zn concentrations of brown rice grown in Cd-polluted soils. Euphytica, 180: 173-179.
CrossRefDirect Link