
Hanafey F. Maswada
Department of Agricultural Botany, Faculty of Agriculture, Tanta University, Tanta, Egypt
Abdelnaser A. Elzaawely
Department of Agricultural Botany, Faculty of Agriculture, Tanta University, Tanta, Egypt
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 23 | Page No.: 1662-1674







ABSTRACT
This study was conducted to investigate the ecological features of three geophytes namely Asparagus stipularis, Cyperus capitatus and Stipagrostis lanata which grow naturally in the Nile Delta coast of Egypt. C. capitatus and S. lanata are growing in non-saline sandy soils and can tolerate drought stress while, A. stipularis is growing in saline and non-saline sandy and calcareous clay soils and can tolerate drought and salt stress. Multivariate analysis of the vegetation of 100 sampled stands supporting growth of the three geophytic species in the study area led to the recognition of four vegetation groups namely, (A) Alhagi graecorum, (B) Cyperus capitatus, (C) Lycium schweinfurthii var. schweinfurthii-Asparagus stipularis and (D) Juncus acutus subsp. acutus. Vegetationally, the vegetation groups associated with the three species can be distinguished into two community types. The first one is psammophytic community comprising vegetation groups A and B that may represent the non-saline sand formations (flats, hummocks and dunes). The second one is halophytic community including vegetation groups C and D that may represent the saline sand flats and salt marsh habitat types, respectively. Sodium adsorption ratio, electrical conductivity, sodium cation, chlorides, silt and sand fractions, pH value, moisture content, bicarbonates and available phosphorus were the most effective soil factors that controlling the abundance and distribution of the plant communities associated with the investigated geophytes. This study showed the ecological features of the selected geophytes in terms of their habitats, associated plant communities and the most edaphic factors controlling their richness and distribution in the study area.
PDF Abstract XML References Citation
Received: January 12, 2013;
Accepted: March 04, 2013;
Published: May 16, 2013
How to cite this article
Hanafey F. Maswada and Abdelnaser A. Elzaawely, 2013. Ecological Investigation of Three Geophytes in the Deltaic Mediterranean Coast of Egypt. Pakistan Journal of Biological Sciences, 16: 1662-1674.
DOI: 10.3923/pjbs.2013.1662.1674
URL: https://scialert.net/abstract/?doi=pjbs.2013.1662.1674
DOI: 10.3923/pjbs.2013.1662.1674
URL: https://scialert.net/abstract/?doi=pjbs.2013.1662.1674
INTRODUCTION
The Mediterranean coast of Egypt extends for 970 km from Sallum eastward to Rafah in three sections: the western coast (550 km), the middle (deltaic) coast (180 km) and the eastern (Sinai) coast (240 km) (Zahran and Willis, 1992). The Deltaic Mediterranean coast of Egypt belongs to the Mediterranean climate type and is differentiated into the following habitat types (1) Sand formations (sand mounds, flats and dunes), (2) Salt marshes (wet and dry), (3) Fertile lands, (4) Reed swamps and (5) Aquatic habitat types (Mashaly, 2001; Maswada, 2004).
Geophytes were first defined by Raunkiaer (1934) as plants with an underground perennating organ (bulb, corm, tuber or rhizome), in which leaves die back annually. Several changes have been made to that definition since. However, in most parts of the world, no evergreen plants are considered to be geophytes (Parsons, 2000). Geophytes are biogeographically widespread around the globe and have adapted to many different habitats. They are the most diverse, however, in the five Mediterranean ecosystems (Esler et al., 1999). Geophytes are considered to be most common in seasonal climates where there is a main stress period that can either be a summer drought or winter (Rossa and von Willert, 1999). Maswada (2009) studied the floristic and synecological features of geophytic plants distributed in the Deltaic Mediterranean coast of Egypt and reported that, the most distributed geophytes were Aeluropus lagopoides, Aetheorhiza bulbosa, Asparagus stipularis, Cynodon dactylon, Cyperus capitatus, Elymus farctus, Pancratium maritimum, Polygonum equisetiforme, Phragmites australis ssp. australis, Sporobolus pungens and Stipagrostis lanata.
The studied geophytic species (Asparagus stipularis, Cyperus capitatus and Stipagrostis lanata) have economic potentialities and could be utilized as promising non-traditional medical crops. Their aerial and/or underground parts contained high amounts of biologically active compounds such as total and simple phenolics, tannins, flavonoids, alkaloids, saponin and cyanogenic glycosides (Hassan and Maswada, 2012; Maswada and Elzaawely, 2013). In addition, the aboveground parts of S. lanata which are a rich source of nutrients and energy, could be utilized in a wide range as a feed for livestock (Maswada and Elzaawely, 2013). Therefore, this study was conducted in order to shed light on the ecological features of Asparagus stipularis, Cyperus capitatus and Stipagrostis lanata throughout the investigation of their soil properties, associated plant communities and the most edaphic factors controlling their richness and distribution in the study area.
MATERIALS AND METHODS
Study area: In the present study, two sites were chosen in the northern part of the Nile Delta coast of Egypt. These sites are the Deltaic Mediterranean coast of Kafr El-Sheikh Governorate and some islands of Lake Borollus (Fig. 1).
Selected species: Three wild geophytic species namely Asparagus stipularis Forssk., Cyperus capitatus Vend. and Stipagrostis lanata (Forssk.) De Winter were selected for this study.
Vegetation analysis: After regular visits to the different sites of the study area, 100 stands (25 m2 each) representing the apparent physiognomic variations in the vegetation and environmental features were used for sampling vegetation of the different habitat types supporting the growth of A. stipularis, C. capitatus and S. lanata. The stands were distributed into 45 stands in summer-autumn season (33 in the Deltaic Mediterranean coast and 12 in Lake Borollus islands) and 55 stands in winter-spring season (40 in the Deltaic Mediterranean coast and 15 in Lake Borollus islands). The sampling processes have been carried out during 2010-2011. In each stand, relative density was estimated according to Shukla and Chandel (1989) while, relative cover was measured by using the line-intercept method (Canfield, 1941). The abundance of species as expressed by the relative values of density and cover were calculated and summed up to give an estimate of its importance value (out of 200). The description and classification of life forms of plant species were determined according to Raunkiaer (1934). Identification, nomenclature and floristic categories of plant species were carried out according to Tutin et al. (1964-1980), Zohary (1966-1972), Tackholm (1974), Dothan (1978-1986), Boulos (1999-2005) and Boulos (2009).
Soil analysis: Five soil-samples that were collected from each stand at a depth of 0-50 cm, were pooled together to form composite sample, air dried, passed through 2 mm sieve to remove gravel and debris and packed in plastic bags ready for physical and chemical analyses.
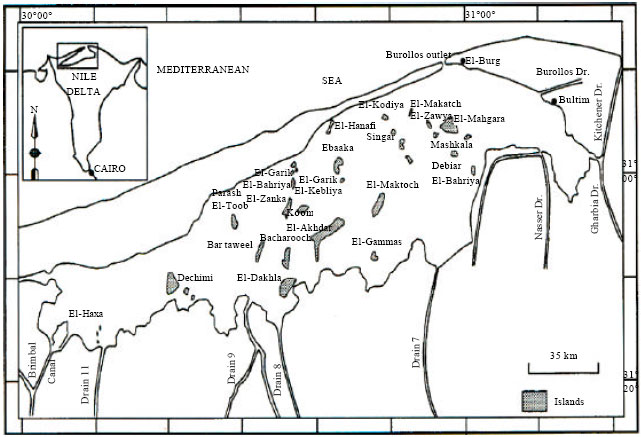 | |
| Fig. 1: | Location map showing the study area |
The procedures followed in estimating the physical and chemical soil characters were determined as described by Ryan et al. (1996).
Data and statistical analysis: Two trends of multivariate analysis of vegetation were applied, namely classification and ordination. The classification technique applied in this study, was the Two Way Indicator Species Analysis (TWINSPAN) (Hill, 1979) while the ordination techniques applied, were the Detrended Correspondence Analysis (DCA) and the Canonical Correspondence Analysis (CCA) (Ter Braak, 1988). The relationships between vegetation groups and edaphic variables are carried out by the ordination diagram produced by CCA-biplot. All statistical treatments applied in the present investigation were according to Gomez and Gomez (1984).
RESULTS
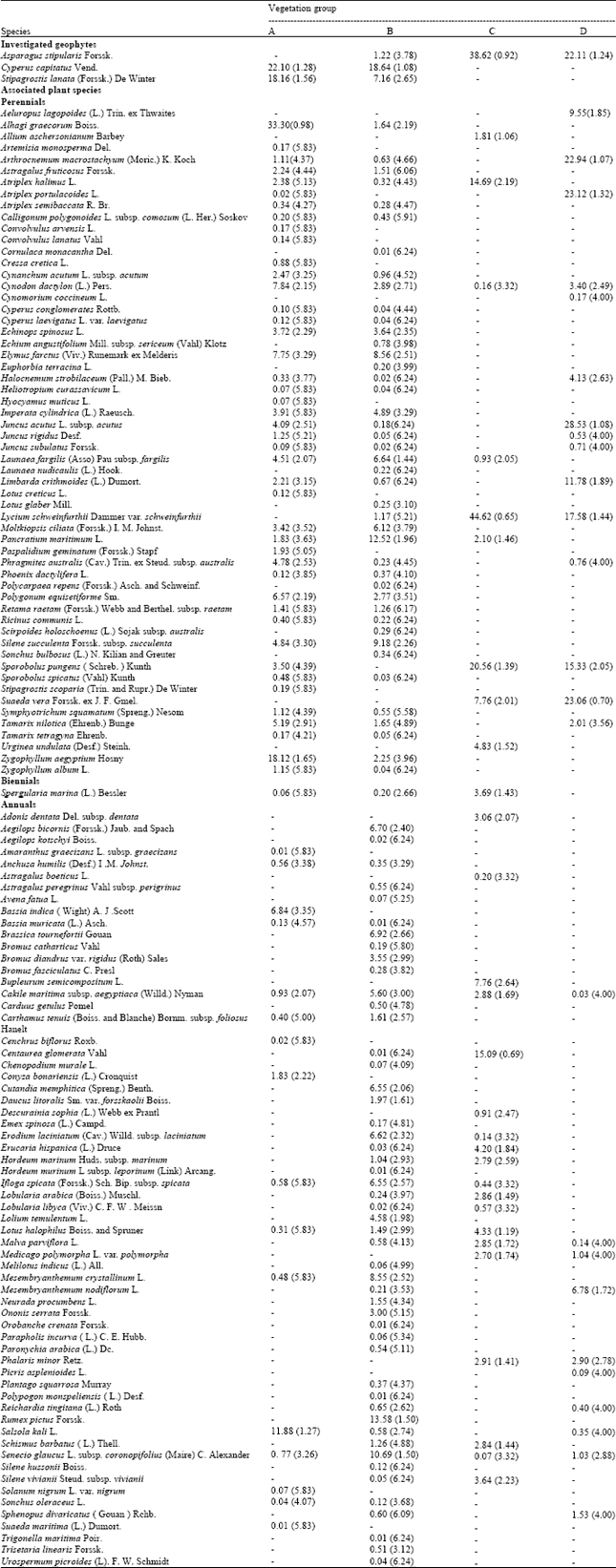
Floristic features: The investigated geophytic species and their associated plant species in the study area are composed of 125 species divided into 61 perennials, one biennial and 63 annuals (Table 1). The life-form spectrum in the study area is predominantly therophytic type (50.4%) followed by cryptophytes (16.8%) which including [geophytes (15.2%) and helophytes (1.6%)], chamaephytes (12.8%), hemi-cryptophytes (11.2%), phanerophytes (7.2%) and parasites (1.6%). The floristic analysis of the recorded species indicates that, the recorded plants are mainly belong to Mediterranean origin (81 species = 64.8%) and partially Saharo-Sindian origin (20 species = 16%), tropical origin (10 species = 8%) and cosmopolitan element (9 species = 7.2%). A further five species (4%) are belonging to different chorotypes with poor representation (Table 1). The investigated geophytes, A. stipularis, C. capitatus and S. lanata are perennials and have subterranean organs (geophytes). A. stipularis (Family: Asparagaceae) is Mediterranean and Saharo-Sindian taxon, C. capitatus (Family: Cyperaceae) is Mediterranean taxon and S. lanata (Family: Poaceae) is Saharo-Sindian taxon.
Soil properties: Results showed in Table 2 revealed that, C. capitatus and S. lanata are growing in sandy soils where sand fraction ranged between 83 and 95% while, A. stipularis is growing in sandy soils and calcareous clay soils where sand fraction ranged from 66.20-94.30% and calcium carbonates reached 45.93%. In addition, A. stipular is occurred in wet or dry soils where moisture content ranged from 0.23-28.92% while C. capitatus and S. lanata often occurred in dry soils. It is interested to note that, A. stipularis occurred in saline and non-saline soils where Electrical Conductivity (EC) ranged between 0.10-6.22 dS m-1 with mean value of 1.35 dS m-1 while C. capitatus and S. lanata often occurred in non-saline soils where the mean values of EC were 0.16 and 0.19 dS m-1, respectively. The soil supporting growth of A. stipularis is more fertile than that supporting growth of C. capitatus and S. lanata. Values of soil variables including organic matter, total nitrogen and available phosphorous and potassium adsorption ratio in the soil supporting growth of A. stipularis were higher than those of the soil supporting growth of C. capitatus and S. lanata.
Vegetation analysis
Classification of stands: The dendrogram resulting from the application of TWINSPAN classification based on the importance values of 125 species recorded in 100 sampled stands supporting growth of the investigated species indicated the distinction of four vegetation groups (Fig. 2, Table 3). Group A comprises 34 stands dominated by Alhagi graecorum with the highest mean Importance Value (IV) of 33.30. The most important species in this group include C. capitatus (IV = 22.10), Zygophyllum aegyptium (IV = 18.12) as well as the important and indicator species S. lanata and Salsola kali with importance values 18.16 and 11.88, respectively. Group B includes 39 stands dominated by C. capitatus (IV = 18.64). The most important species in this group are Rumex pictus (indicator species, IV = 13.58), Pancratium maritimum (IV = 12.52), Senecio glaucus subsp. coronopifolius (IV = 10.69), Silene succulenta subsp. succulenta (IV = 9.18), Elymus farctus (IV = 8.56) and S. lanata (IV = 7.16). Group C consists of 11 stands codominated by L. schweinfurthii var. schweinfurthii (IV = 44.62) and A. stipularis (IV = 38.62). The most important species in this group include Sporobolus pungens (IV = 20.56), Centaurea glomerata (IV = 15.09) and the indicator species Atriplex halimus (IV = 14.69). Group D comprises 16 stands dominated by Juncus acutus subsp. acutus (IV = 28.53). The most important species are Atriplex portulacoides (indicator species, IV = 23.12), Suaeda vera (IV = 23.06), Arthrocnemum macrostachyum (IV = 22.94), A. stipularis (IV = 22.11) and L. schweinfurthii var. schweinfurthii (IV = 17.58). As shown in Table 3, A. stipularis is recorded in three vegetation groups (B, C and D).
| Table 1: | Life-forms and floristic categories of investigated geophytic plants and their associated plant species in the study area |
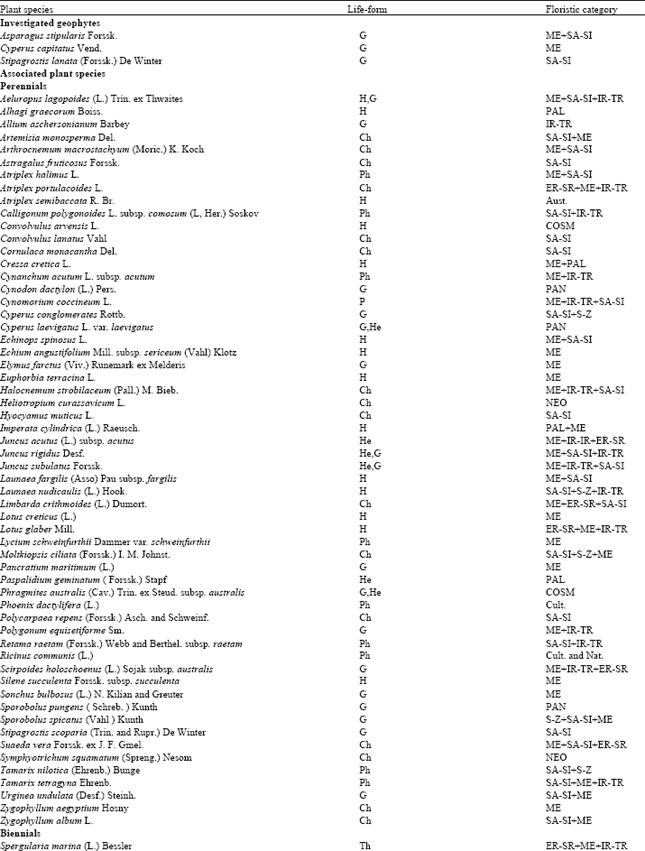 | |
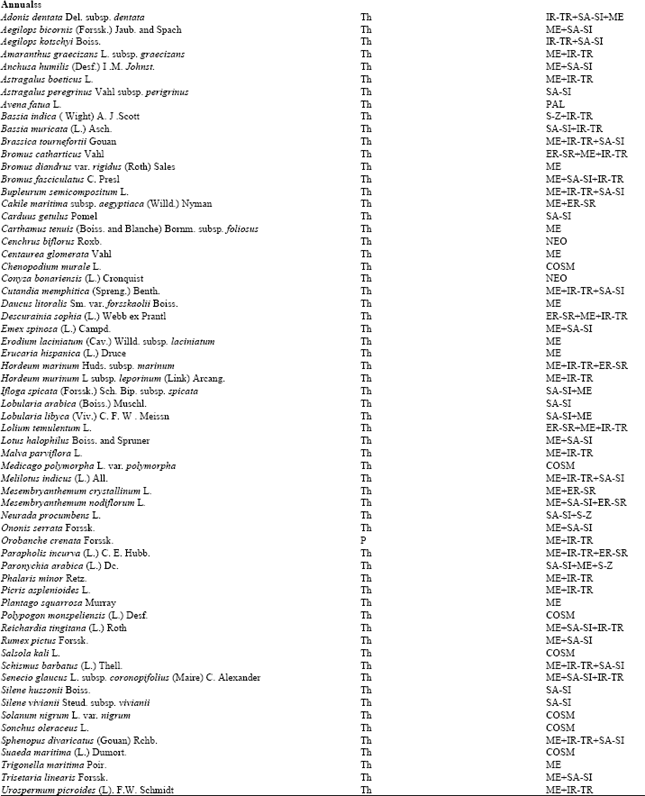 | |
| Legend to life-forms: Ph: Phanerophytes, Ch: Chamaephytes, H: Hemicryptophytes, G: Geophytes, He: Helophytes, P: Parasites, Th: Therophytes. Legend to floristic categories: COSM: Cosmopolitan, PAN: Pantropical, PAL: Palaeotropical, NEO: Neotropical, ME: Mediterranean, ER-SR: Euro-Siberian, SA-SI: Saharo-Sindian, IR-TR: Irano-Turanian, S-Z: Sudano-Zambezian, Nat.: Naturalized, Cult.: Cultivated and Aust.: Australia | |
| Table 2: | Maximum, minimum and mean values the different soil variables in the stands supporting the growth of the studied geophytic plants |
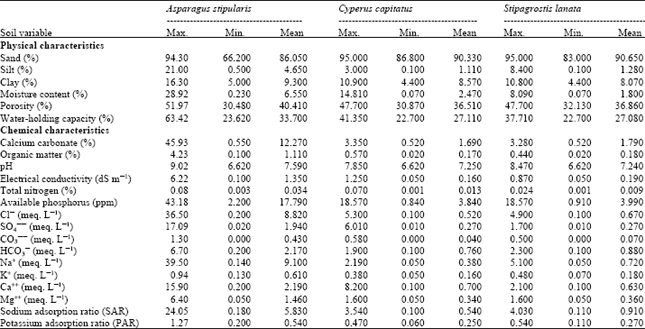 | |
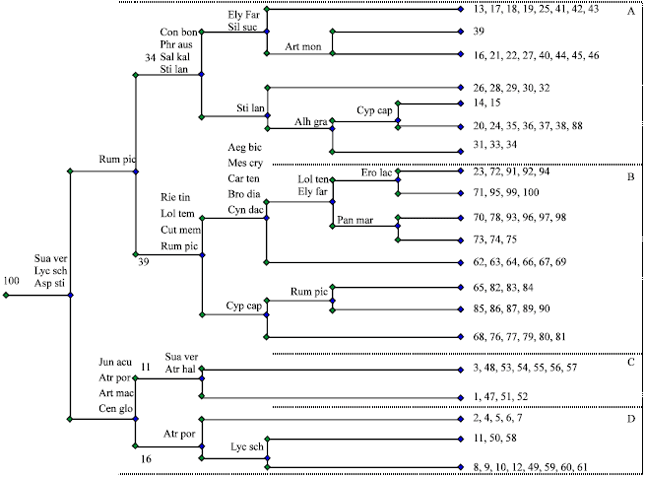 | |
| Fig. 2: | Two Way Indicator Species Analysis (TWINSPAN) dendrogram of the 100 sampled stands based on the importance values of the plant species dominated or associated with the three selected geophytic plants. The indicator species are abbreviated by the first three letters of genus and species, respectively |
| Table 3: | Mean and coefficient of variation (C.V) of the importance values (out of 200) of the investigated geophytes and the associated plant species in different vegetation groups resulting from TWINSPAN classification of sampled stands |
 | |
It is occurred as codominant species in group C (IV = 38.62) and important species in group D (IV = 22.11) while it rarely occurred in group B where it attained importance value of 1.22. C. capitatus is recorded as dominant species in group B (IV = 18.64) and as important species in group A (IV = 22.10). Also, S. lanata is recorded in two vegetation groups (A and B) as important species and indicator species in group A.
Ordination of stands: The ordination of 100 sampled stands which obtained by Detrended Correspondence Analysis (DCA) (Fig. 3) indicated that, the vegetation groups yielded by TWINSPAN classification are more or less distinguishable and having a clear pattern of segregation on the ordination planes. All the vegetation groups are located in the positive side of the first and second ordination axes. It is obvious that, group A is little superimposed with group B, whilst group C is highly superimposed with group D. Groups A and B are segregated at right side of DCA diagram. Contrary, groups C and D are separated at the left side of DCA diagram. On the other hand, groups A and D are segregated at the lower part of the DCA diagram while, groups B and C are segregated at the upper side of DCA diagram. It is interested vegetationally to denote that, the vegetation groups supporting growth the three studied species can be distinguished into the following community types (1) Group A (Alhagi graecorum) and group B (Cyperus capitatus) may represent the non-saline sand formations (flats, hummocks and dunes) and (2) Group C (Lycium schweinfurthii var. schweinfurthii-Asparagus stipularis) and group D (Juncus acutus subsp. acutus) may represent the saline sand flats and salt marsh habitat types, respectively.
Vegetation-soil relationships
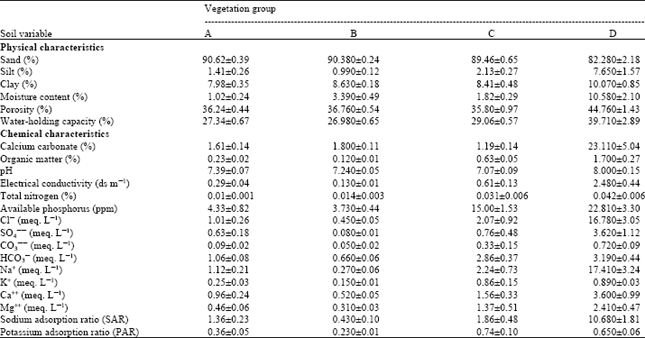
Relationship between soil variables and vegetation groups: Soil variables of the four vegetation groups in different habitats derived from TWINSPAN classification are presented in Table 4. The soil texture in all groups is formed mainly of coarse fraction (sand) and partly of fine fractions (silt and clay). The maximum mean percentage of sand fraction (90.62%) is attained in group A while, the minimum mean value (82.28%) is attained in group D. Group D attained the maximum mean percentages of other physical characteristics of soil samples. The minimum mean percentages of clay fraction and moisture content are found in group A. Group B attained the minimum percentages of silt fraction and water holding capacity while, the minimum percentage of porosity is in group C. Group D attained the maximum mean percentages of most chemical characteristics of the soil samples except Potassium Adsorption Ratio (PAR) which attained the maximum mean value (0.74) in group C. On the other hand, the minimum mean values of most chemical characteristics of soil samples are found in group B except CaCO3 content and pH value in group C, organic matter and EC in group B and total nitrogen in group A.
Correlation between soil variables and vegetation gradients: The relationship between vegetation and edaphic variables is indicated on the ordination diagram produced by Canonical Correspondence Analysis (CCA) of the biplot of species-environment as shown in Fig. 4. It is obvious that, the most effective soil variables controlling the distribution and abundance of psammophytic vegetation associated with the studied geophytic plants is sand fraction.
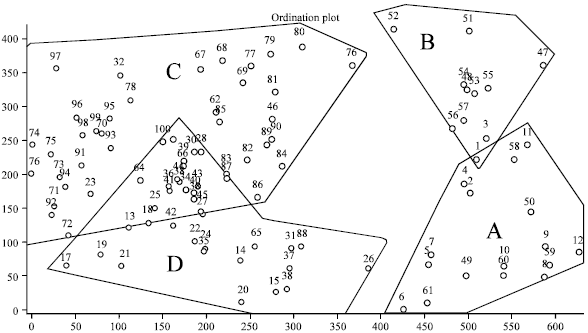 | |
| Fig. 3: | Detrended correspondence analysis (DCA) ordination diagram of the 100 stands supporting growth of the three selected geophytic species with TWINSPAN groups superimposed |
| Table 4: | Mean value and standard error of the different soil variables at depth (0-50 cm) in the stands representing the different vegetation groups of the three selected geophytic species obtained by TWINSPAN classification of the different habitats in the study area |
 | |
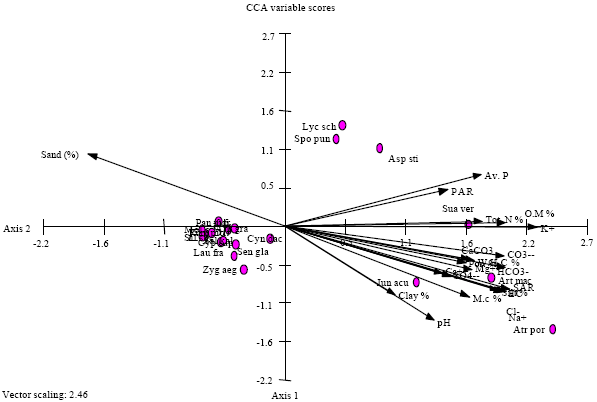 | |
| Fig. 4: | Canonical correspondence analysis (CCA) ordination diagram with edaphic variables represented by arrows of the three selected geophytic species. The indicator and preferential species are abbreviated to the first three letters of each of the genus and species, respectively |
Furthermore, Sodium Adsorption Ratio (SAR), EC, Na+, Cl¯, silt fraction, pH value, moisture content, HCO3¯ and available phosphorus are the most soil factors affecting on the distribution and abundance of halophytic vegetation. Moreover, it is clear that the dominant and/or codominant species of the halophytic vegetation (group C and D) are separated at the right side of the CCA diagram while, the dominant and/or codominant species of the psammophytic vegetation (group A and B) are segregated at the left side of the CCA diagram.
DISCUSSION
Floristically, the life-form spectra have widely been used by ecologists and chorologists in the vegetation and floristic studies (Cain and de Oliveira Castro, 1959) and provide informations which may help in assessing the response of vegetation to variations in environmental factors (Ayyad and El-Ghareeb, 1982) and also indicate climate and microclimates (Kershaw and Looney, 1985). Raunkiaer (1934) designated the Mediterranean climate type as “therophyte climate” because of the high percentage (more than 50% of the total species) of this life-form in the Mediterranean floras. In the present work, the life-form spectrum in the study area is predominantly therophytic type (50.4%) followed by cryptophytes (16.8%) which including geophytes (15.2%) and helophytes (1.6%). This trend is similar to life-form spectra reported in the Deltaic Mediterranean coast of Egypt by Zahran et al. (1990), Shaltout et al. (1995), Khedr (1999), Mashaly (2001), Mashaly (2002), Maswada (2004), Maswada (2009), Galal and Fawzy (2007) and Hassan et al. (2009).
Phytogeographically, Egypt is the meeting point of floristic elements belonging to at least four phytogeographical regions: the African Sudano-Zambezian, the Asiatic Irano-Turanian, the Afro-Asiatic Saharo-Sindian and the Euro-Afro-Asiatic Mediterranean (El-Hadidi, 1993). The floristic analysis of the recorded species in the study area shows that, the recorded plants are mainly belong to Mediterranean origin (81 species = 64.8%) and partially Saharo-Sindian origin (20 species = 16%). Therefore, the study area is obviously belonging to the Mediterranean Territory with slightly extending into Saharo-Sindian Territory (Maswada (2009).
Edaphically, the investigated plants occurs in harsh environment where C. capitatus and S. lanata are growing in dry sandy soils and A. stipularis is growing in saline dry sandy or calcareous clay soils. Therefore, C. capitatus and S. lanata can tolerate drought stress and A. stipularis can tolerate drought and salt stress.
Vegetationally, A. stipularis and C. capitatus are dominant or codominant species in the study area while S. lanata not well. This may be due to that, A. stipularis and C. capitatus are related to the study area which they are Mediterranean taxa while S. lanata is Saharo-Sindian taxon. The vegetation analysis of the plant communities associated with the studied geophytes dealt that, there are two major plant communities, psammophytes (group A and B) and halophytes (group C and D). According to the Braun-Blanquet’s floristic association system (Braun-Blanquet, 1932), the identified vegetation groups representing the salt marshes in the present study may be related to class Arthrocnemetea while, the vegetation groups representing the sand formations may be related to class Echinopetea.
In the salt marsh and saline sand flat habitat types in the study area, the dominant and/or codominant halophytic and salt tolerant geophytic species are Juncus acutus subsp. acutus and Lycium schweinfurthii var. schweinfurthii-Asparagus stipularis. The other important halophytic plants in these habitat types include Arthrocnemum macrostachyum, Atriplex halimus, Atriplex portulacoides, Centaurea glomerata, Sporobolus pungens and Suaeda vera. All of the leading these dominant and common halophytes constitute the major part of the vegetation composition of both littoral and inland salt marshes in Egypt (Zahran, 1982). The halophytic vegetation in the present study may be related to class Salicornietea europaeae which comprises all plant communities of the salt marshes in the circumference of Mediterranean coastal belt (Zohary, 1973). Also, the halophytic vegetation in the present work may be related to Salicornion alliance (Tadros and Atta, 1958). On the other hand, the vegetation groups in the sand formations are dominated by the psammophytic plants, Alhagi graecorum and Cyperus capitatus. The other common associated psammophytic species in this habitat type comprise Elymus farctus, Pancratium maritimum, Rumex pictus, Salsola kali, Senecio glaucus subsp. coronopifolius, Silene succulenta subsp. succulent, Stipagrostis lanata and Zygophyllum aegyptium. It is obvious that, most of these plant species are either sand accumulator or sand loving species which play an important role in the formation processes and development of sand flats, mounds, hummocks, hillocks and dunes. The vegetation groups of the sand formation habitat type may be related to the Thymelaeion hirsutae alliance (El-Ghonemy and Tadros, 1970).
The application of DCA ordination in the sampled stands dealt that, vegetation groups A and B are superimposed as well as vegetation group C and D. This is may be due to the similarity between vegetation groups A and D and between vegetation group C and D. Group A and B may represent psammophytic vegetation and groups C and D may represent halophytic vegetation associated with the investigated geophytes. In the present study, SAR, EC, Na+, Cl¯, silt fraction, pH value, moisture content, HCO3¯ and available phosphorus were the most critical soil factors controlling the distribution of plant communities associated with the studied geophytic plants. This agrees more or less findings of El-Demerdash et al. (1990) in Damietta coastal land, Shaltout et al. (1995) in the Mediterranean region of the Nile Delta, Mashaly et al. (2008) in the Deltaic Mediterranean coastal habitat and Hassan et al. (2009) in Lake Borollus Protectorate of Egypt. On the other hand, the most active soil factor controlling the distribution of psammophytic vegetation in the study area is sand fraction. This agrees with findings of Shaltout et al. (1995), Mashaly (2001), Hassan et al. (2009) and Maswada (2009).
REFERENCES
- Ayyad, M.A. and R.E.M. El-Ghareeh, 1982. Salt marsh vegetation of the Western Mediterranean Desert of Egypt. Vegetatio, 49: 3-19.
CrossRefDirect Link - Cain, S.A. and G.M. de Oliveira Castro, 1959. Manual of Vegetation Analysis. Harper and Brothers, New York, USA., Pages: 325.
Direct Link - Canfield, R.H., 1941. Application of the line interception method in sampling range vegetation. J. For., 39: 388-394.
Direct Link - El-Demerdash, M.A., M.A. Zahran and M.S. Serag, 1990. On the ecology of the deltaic Mediterranean coastal land, Egypt. III. The habitat of salt marshes of Damietta-Port Said coastal region. Arab Gulf J. Scient. Res., 8: 103-119.
Direct Link - Esler, K.J., P.W. Rundel and P. Vorster, 1999. Biogeography of prostrate-leaved geophytes in semi-arid South Africa: Hypothesis on functionality. Plant Ecol., 142: 105-120.
Direct Link - Galal, T.M. and M. Fawzy, 2007. Sand dune vegetation in the coast of Nile Delta, Egypt. Global J. Environ. Res., 1: 74-85.
Direct Link - Khedr, A.H.A., 1999. Floristic composition and phytogeography in a Mediterranean Deltaic lake (Lake Burollos), Egypt. Ecologia Mediterranea, 25: 1-11.
Direct Link - Mashaly, I.A., 2001. Contribution to the ecology of the deltaic mediterranean coast, Egypt. J. Biol. Sci., 1: 628-635.
CrossRefDirect Link - Mashaly, I.A., 2002. Ecological Studies on Zygophyllum aegyptium in the Deltaic Mediterranean Coast of Egypt Pak. J. Biol. Sci., 5: 152-160.
CrossRefDirect Link - Mashaly, I.A., I.E. El-Habashy, E.F. El-Halawany and G. Omar, 2008. Habitats and plant communities in the nile delta of Egypt I. deltaic mediterranean coastal habitat. Pak. J. Biol. Sci., 11: 2532-2544.
CrossRefPubMedDirect Link - Maswada, H.F. and A.A. Elzaawely, 2013. Nutritive value of Stipagrostis lanata (Forssk.) De Winter as a feed for livestock. Asian J. Crop Sci., 5: 216-221.
CrossRefDirect Link - Parsons, R.F., 2000. Monocotyledonous geophytes: Comparison of California with Victoria, Australia. Aust. J. Bot., 48: 39-43.
Direct Link - Raunkiaer, C., 1934. The Life Forms of Plants and Statistical Plant Geography. Clarendon Press Oxford, London, United Kingdom, Pages: 632.
Direct Link - Rossa, B. and D.J. von Willert, 1999. Physiological characteristics of geophytes in semi-arid Namaqualand, South Africa. Plant Ecol., 142: 121-132.
Direct Link - Shaltout, K.H., H.F. El-Kady and Y.M. Al-Sodany, 1995. Vegetation analysis of the Mediterranean region of Nile Delta. Vegetation, 116: 73-83.
CrossRefDirect Link - Tadros, T.M. and B.A.M. Atta, 1958. The plant communities of barley fields and uncultivated desert areas of Mareotis (Egypt). Vegetatio, 8: 161-175.
CrossRefDirect Link - Zahran, M.A., M.A. El-Demerdash and I.A. Mashaly, 1990. Vegetation types of the deltaic Mediterranean coast of Egypt and their environment. J. Vegetation Sci., 1: 305-310.
CrossRefDirect Link