
Caroline Borges Bevilacqua
Universidade Federal de Pelotas, Faculdade de Agronomia Eliseu Maciel, Dep. Fitotecnia, Programa de Pos-graduacao em Ciencia e Tecnologia de Sementes, Caixa Postal 354, CEP 96.001-970, Pelotas, RS, Brasil
Daisy Ramirez Monzon
Universidade Federal de Pelotas, Faculdade de Agronomia Eliseu Maciel, Dep. Fitotecnia, Programa de Pos-graduacao em Ciencia e Tecnologia de Sementes, Caixa Postal 354, CEP 96.001-970, Pelotas, RS, Brasil
Eduardo Venske
Universidade Federal de Pelotas, Faculdade de Agronomia Eliseu Maciel, Dep. Fitotecnia, Programa de Pos-graduacao em Ciencia e Tecnologia de Sementes, Caixa Postal 354, CEP 96.001-970, Pelotas, RS, Brasil
Supratim Basu
115 Plant Sciences Building, University of Arkansas, Fayetteville, AR-72701, USA
Paulo Dejalma Zimmer
Universidade Federal de Pelotas, Faculdade de Agronomia Eliseu Maciel, Dep. Fitotecnia, Programa de Pos-graduacao em Ciencia e Tecnologia de Sementes, Caixa Postal 354, CEP 96.001-970, Pelotas, RS, Brasil
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 22 | Page No.: 1618-1622







ABSTRACT
Low temperature or cold stress and deep sowing plays a pivotal role in limiting rice (Oryza sativa L.) productivity in the temperate rice growing regions as well as in tropical high lands worldwide. A better understanding of stress tolerance mechanism in rice plants will help to develop rice germplasm with improved field level tolerance under variable temperature and sowing deep conditions. Using previously developed stress indices, this study presents results from low temperature and deep sowing screening of four rice genotypes. A group of 25 seeds per replicate (total of 3 replicates) was subjected to stress by deep sowing (15 cm) while another group was subjected to cold stress (13°C-10 h/18°C-14 h) and the control group remained under optimum conditions (25°C and sowing depth of 1.5 cm). The Geometric Mean (GM), Stress Tolerance Index (STI) and Stress Susceptibility Index (SSI) were used to evaluate the genotypic performance under control and stress conditions. The results indicate that it was possible to identify superior genotypes for tolerance based on their stress indices. The indices although correlated, were found to be effective for the selection of genotypes with good yield potential under control and stress treatments and can now be used for genotypic screening under field conditions.
PDF Abstract XML References Citation
Received: January 15, 2013;
Accepted: March 02, 2013;
Published: May 08, 2013
How to cite this article
Caroline Borges Bevilacqua, Daisy Ramirez Monzon, Eduardo Venske, Supratim Basu and Paulo Dejalma Zimmer, 2013. Application of Stress Indices for Low Temperature and Deep Sowing Stress Screening of Rice Genotypes. Pakistan Journal of Biological Sciences, 16: 1618-1622.
DOI: 10.3923/pjbs.2013.1618.1622
URL: https://scialert.net/abstract/?doi=pjbs.2013.1618.1622
DOI: 10.3923/pjbs.2013.1618.1622
URL: https://scialert.net/abstract/?doi=pjbs.2013.1618.1622
INTRODUCTION
Low temperatures have a strong impact on the survival, growth, reproduction and distribution of plants. The plants are characterized by an inherent level of resistance to low temperatures which reduces the metabolic activity. This level of resistance can vary among individual plants and species. The cold stress is characterized by physiological perturbations, generally called low-temperature damages (Guimaraes et al., 2011; Hudak and Salaj, 1999; Yan et al., 2010; Zhang et al., 2011). The germination process followed by fast and uniform seedling emergence is essential for the successful crop development (Balbinot et al., 2001). However, if the seeds remain under the soil for long time this would impair germination which will be accompanied by abiotic stresses like low temperature or biotic stresses like pathogens (Da Cruz et al., 2007). Consequently it will decrease the crop yield and increase the productivity cost. The abiotic stresses caused owing to deep sowing are also an important factor that plays a major role in impairing plant development. Hence the long term goal for breeding is the development of germplasm with improved field tolerance under different stress conditions owing to low temperature (Cruz and Milach, 2000) and seed sowing depths.
Rice is a temperature-sensitive crop; low temperatures and deep sowing dramatically reduce its production. Good tolerance at the seedling stage is an important character for stable rice production. Brazil is one of the largest rice producing countries in the world. However the conditions are not always favorable which makes early seedlings rotten thereby causing massive seed loss and delayed growth (Cruz and Milach, 2000). Hence it becomes important to develop rice plants that can circumvent the atrocities of nature. Previous reports have shown the genetics and physiological responses of plants to temperature stress but yield base indices are needed for the evaluation of low temperature and deep sowing stress tolerance for applied plant breeding programs. Several yield based stress indices have been developed that may be applicable for low temperature and deep sowing tolerance. The Geometric Mean (GM) and the Stress Tolerance Index (STI) (Fernandez 1993) have been used for comparing genotypic performance across years or environments. STI was developed to identify genotypes that perform well under both stress and control conditions. The Stress Susceptibility Index (SSI) (Fischer and Maurer, 1978) is a ratio of genotypic performance under stress and non-stress conditions, adjusted for the intensity of each trial and have been found to be correlated with yield and canopy temperature in wheat (Rashid et al., 1999). These different indices may be applicable to other abiotic stress traits, such as low temperature or deep sowing stress tolerance. This study presents the low temperature and deep sowing stress screening of rice plants in Brazil for selection of cold and depth stress (deep sowing) tolerant rice genotypes with good yield potential and applicability of several stress indices for genotypic stress under the aforesaid stress treatments.
MATERIAL AND METHODS
Plant materials: Four rice genotypes were used in this study: the Brilhante (cold tolerant, Fagundes et al., 2010), IRGA 422 CL (as model rice), BRS 6 Chuí (cold susceptible, Mertz et al., 2009) and one red rice ecotype 116.
Germination test: The experiment was carried out with 100 seeds in four replicates. The seeds were set on paper roll previously dampened with sterile double distilled water using 2.5 times the paper dry mass. After sowing, these rolls were kept inside plastic bags in growth chamber at 25°C for 14 days. The evaluations were made on 7 days and 14 days respectively after sowing, according to Brazilian standards (Ministry of Agriculture and Agrarian Reform 2009). Germination Percent (GP) was calculated as described by Scott et al. (1984). The germination count was calculated as described in Brazilian standards (Ministry of Agriculture and Agrarian Reform 2009).
Cold test: The cold test was conducted as described by Vieira and Carvalho (1994) with some modifications in 4 replicates each with 100 seeds. The seeds were distributed uniformly on previously dampened filter paper and then these rolls were kept inside plastic bags in growth chamber for 7 days at 10°C and these treated seeds were then kept inside the growth chamber (25°C) for 7 days. The evaluation was made in a similar way like Germination test count.
Seed treatment: The soil was previously autoclaved and then dried in a hot air oven (70°C). The experiment was carried out in four replicates, with 25 seeds per genotype under three different conditions: For cold stress treatment the seeds were sowed at 1.5 cm depth in cups (200 mL) and kept in growth chamber under alternate temperatures (13°C-10 h/18°C-14 h) while for deep sowing stress, seeds were sowed at 15 cm deep in PVC tubes and kept in growth chamber at 25°C. Seeds sowed at a depth of 1.5 cm in cups served as control were kept in growth chamber at 25°C. The measurements of shoot length and root length were made after 7 days and 14 days respectively. The Stress Susceptibility Index (SSI): ((1- (Ys/Yp))/(1-(Xs/Xp)), Geometric Mean (GM): ((YsxYp)1/2), Stress Tolerance Index (STI): ((YpxYs)/Xp2), were determined using the equations for SSI (Fischer and Maurer, 1978), GM, STI and SI (Fernandez, 1993), respectively. The data obtained in this study were used in place of the genotypic mean values for seedling length under stress (Ys) and under control (Yp) variables, respectively, in the equations for the above indices. Xs and Xp are the mean seedling length of all genotypes per trial under stress and control conditions.
Statistical analysis: All the experimental data presented in this study are mean from three replicates. The significance of the values so obtained was evaluated by ANOVA and Tukey test (p = 0.05). The values for the indices so obtained were correlated by Pearson correlation analysis using t-test (p = 0.01 and p = 0.05) using WinSTAT version 1.0 software package.
RESULTS AND DISCUSSION
Rice (Oryza sativa L.) is a model plant for the genetics and physiology of cultivated grasses, especially due to its high information content available by synteny with other cereals (Gale and Devos, 1998) and the genetic variability that it has to characters for importance of seed physiology. Furthermore, it is a major crop species from South of Brazil, which is currently responsible for approximately 60% of national production. Therefore, rice is an excellent source of research, either through regional and national economic importance, but also is of great importance by virtue of the biology of the species itself and other cereals. The ability of the plants to emerge efficiently and rapidly from the soil or under water is a desirable quality that needs to be incorporated into rice cultivars (Redona and Mackill, 1996).
| Table 1: | First count (%), Germination (%) and cold test of different rice genotypes |
 | |
| Means followed by the same capital letters differ by Tukey test at 5% level of significance | |
| Table 2: | Total seedling length (root+shoot) in the different rice genotypes after cold stress and seed sowing deep stress |
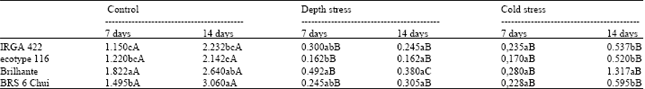 | |
| Means not followed by the same lowercase and uppercase letters in the column and in the row differ by Tukey test at 5% level of significance, respectively | |
The result presented here puts an insight into the plant vigor which was assessed by, first germination count, germination test and cold test (Table 1). Among the four genotypes (BRS6 Chui, Brilhante, IRGA 422 CL and one red rice ecotype 116) studied here it was observed that BRS6 Chui had the best physiological characteristics demonstrated by germination test and cold test which varied from 94-97% followed by Brilhante and IRGA 422 CL (87-88%) while red rice showed the less vigor (71%) Previously it has been shown by Perreto et al. (1993) that the red rice have varying rates of dormancy which is consistent with our observation. All these results were statistically significant as estimated by Tukey test using winSTAT software package. The next part of our study focused on the significance of genotype and stress-level interaction. The effect of stress level and genotypes was considered fixed because these were the only levels and genotypes of interest. The rice genotypes did not show any significant variation in shoot length among themselves after 7 days of exposure to deep sowing stress or cold stress (data not shown). However significant differences were observed in the total length (root length+shoot length) among the cultivars after 7 days of exposure to either of the stresses (Table 2). Hence in the light of this observation we can suggest that 7 days exposure was not enough for screening genotypes for cold and deep sowing tolerance at germination stage on the basis of their shoot length. Previously it has been shown that corn genotypes when exposed to cold stress (10°C) for 7 and 14 days respectively showed a detrimental effect on seedling development. Earlier work done with 494 rice genotypes for screening cold tolerance over a period of four years showed that Tomoe Mochi, Amber, Diamond Bright, BRS Querência and Japanese Grande (Japonica group) showed better performance during two years of review at 14.7°C in first year and then third year at 3.3°C to 15.6°C min and maximum (Fagundes et al., 2010). Moreover it was also reported that the cultivars from Japonica subspecies showed more tolerance to low temperature stress as compared to the Indica subspecies (Mertz et al., 2009). Our results showed that Brilhante showed more shoot length after 14 days of cold stress followed by IRGA 422 CL and red rice ecotype 116 while BRS6 Chui showed the least shoot length. All these results were statistically significant as determined by Tukey test. Hence, in the light of this observation we can conclude that BRS6 Chui is highly susceptible to cold stress as compared to the other genotypes while Brilhante was cold tolerant, this was consistent with earlier observations. However, when the plants were exposed to deep sowing stress it was observed that all the plants were highly susceptible as they showed significantly less seedling length (Table 2). Amidst all the cultivars studied here the red rice ecotype 116 showed better shoot length as compared to all the other rice genotypes under study which gives us an idea that this cultivar is moderately tolerant to deep sowing stress which is consistent with earlier findings from Yoshida (1981) who showed that 12% of red rice seeds germinated at 12 cm deep but at 16 cm germination was only 1%. Earlier when Gealy et al. (2000) tested the emergence of several ecotypes of weedy red rice in different textures of soil at a deep of 7.5 cm found that all the ecotypes emerged uniformly. Previously work done with segregating rice population for analysis of plant vigor showed that red rice ecotype 45 exhibited higher germination at 20 cm deep in the soil (Malone et al., 2007). Hence we can conclude that not only abiotic stress caused by low temperature but also stress mediated by deep sowing is detrimental for rice plant development and productivity.
The Stress Susceptibility Indices (SSI) and GM for shoot length for 7 days and 14 days of deep sowing stress were correlated negatively which is not desirable. However, for the cold stress (7 days) the GM mean and CSI correlated positively, but on the contrary the GM for deep sowing and cold stress showed a positive correlation which is inconclusive (Table 3, 4).
| Table 3: | Averages for seedling shoot length (cm) subjected to stress 14 days after sowing |
 | |
| Means not followed by the same lowercase and uppercase letters in the column and in the row differ by Tukey test at 5% level of significance, respectively | |
| Table 4: | Correlation between Indices of Tolerance and Susceptibility and Geometric Mean for seedling length of the four genotypes |
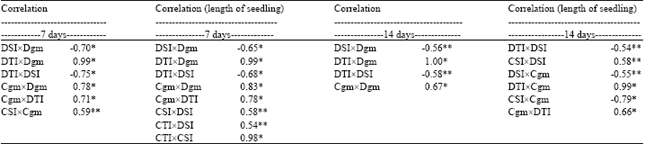 | |
| D: Deep, C: Cold, SI: Susceptibility Index; TI: Tolerance Index, GM: Geometric Mean; Statistical Significance was estimated by t test (*) and (**) significant (>0.05) and (>0, 01) of likelihood, respectively | |
This brings us to our earlier conclusion that it is not possible to screen rice genotypes for cold and deep stress after 7 days. Florido et al. (2009) have shown that the index of tolerance can help in breeding programs to permit the investigation of the performance of that germplasm under adverse environmental conditions. However the screening results for seedling length after 14 days under either of the stresses showed a positive correlation between the Tolerance Indices (STI) and GM, thereby justifying the effectivity of the methodology. Our results are consistent with previous reports on bean genotypes that showed heat tolerance index and GM proved are the most useful indices for the evaluation of genotypic performance under heat stress and they were highly correlated (Porch, 2006). Corroborating this result, Salem et al. (2010) also found that STI and GM have high positive correlation, though low correlation was observed between GM and SI while screening wheat genotypes under drought stress.
The ability of plants to tolerate the harsh effects of abiotic stress plays a critical role in improving the crop yield. Low temperature or cold stress and deep sowing stress are two major constraints in rice yield owing to global climactic change. In this evaluation of screening rice genotypes in Brazil under cold and deep sowing stress it was observed that STI and GM are the most effective stress indices for the selection of genotypes with good yield potential under stress and low-stress conditions.
ACKNOWLEDGMENT
This study was supported by CNPq (National Council for Scientific and Technological Development-Brazil), CAPES (Coordination of Improvement of Higher Education Personnel) and FAPERGS (Research Foundation of the State of Rio Grande do Sul).
REFERENCES
- Balbinot Jr., A.A., N.G. Fleck, D. Agostinetto, M.A. Rizzardi, A. Merotto Jr. and R.A. Vidal, 2001. Emergence and early growth of flooded rice cultivars influencing the competitive ability with weeds. Planta Daninha, 19: 305-316.
CrossRef - Cruz, R.P.D. and S.C.K. Milach, 2000. Breeding for cold tolerance in irrigated rice. Ciencia Rural, 30: 909-917.
CrossRef - Fischer, R.A. and R. Maurer, 1978. Drought resistance in spring wheat cultivars. I. Grain yield responses. Aust. J. Agric. Res., 29: 897-912.
CrossRefDirect Link - Gale, M.D. and K.M. Devos, 1998. Comparative genetics in the grasses. Proc. Natl. Acad. Sci. USA., 95: 1971-1974.
PubMedDirect Link - Gealy, D.R., N.E. Saldain and R.E. Talbert, 2000. Emergence of red rice (Oryza sativa) ecotypes under dry-seeded rice (Oryza sativa) culture. Weed Technol., 14: 406-412.
Direct Link - Mertz, L.M., F.A. Henning, R.C. Soares, R.F. Baldiga, F.B. Peske and D.M.D. Moraes, 2009. Physiological changes in rice seeds exposed to cold in the germination phase. Revista Brasileira Sementes, 31: 262-270.
CrossRef - Porch, T.G., 2006. Application of stress indices for heat tolerance screening of common bean. J. Agronomy Crop Sci., 192: 390-394.
CrossRef - Rashid, A., J.C. Stark, A. Tanveer and T. Mustafa, 1999. Use of canopy temperature measurements as a screening tool for drought tolerance in spring wheat. J. Agron. Crop Sci., 182: 231-237.
CrossRefDirect Link - Redona, E.D. and D.J. Mackill, 1996. Mapping quantitative trait loci for seedling vigor in rice using RFLPs. Theor. Applied Genet, 92: 395-402.
CrossRefDirect Link - Scott, S.J., R.A. Jones and W.A. Williams, 1984. Review of data analysis methods for seed germination. Crop Sci., 24: 1192-1198.
Direct Link - Zhang, S., H. Jiang, S. Peng, H. Korpelainen and C. Li, 2011. Sex-related differences in morphological, physiological and ultrastructural responses of Populus cathayana to chilling. J. Exp. Bot., 62: 675-686.
CrossRef