
H. Abou-Elhakam
Department of Zoology, Faculty of Science, Cairo University, Egypt
I. Rabee
Theodore Bilharz Research Institute, Giza, Egypt
S. EL Deeb
Department of Zoology, Faculty of Science, Cairo University, Egypt
A. EL Amir
Department of Zoology, Faculty of Science, Cairo University, Egypt
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 22 | Page No.: 1449-1458







ABSTRACT
Yet no vaccine to protect ruminants against liver fluke infection has been commercialized. In an attempt to develop a suitable vaccine against Fasciola gigantica (F. gigantica) infection in rabbits, using 97 kDa Pmy antigen. It was found that, the mean worm burdens and bile egg count after challenge were reduced significantly by 58.40 and 61.40%, respectively. On the other hand, immunization of rabbits with Pmy induced a significant expression of humoral antibodies (IgM, total IgG, IgG1, IgG2 and IgG4) and different cytokines (IL-6, IL-10, IL-12 and TNF-α). Among Ig isotypes, IgG2 and IgG4 were most dominant Post-infection (PI) while, recording a low IgG1 level. The dominance of IgG2 and IgG4 suggested late T helper1 (Th1) involvement in rabbit’s cellular response. While, the low IgG1 level suggested Th2 response to adult F. gigantica worm Pmy. Among all cytokines, IL-10 was the highest in rabbits immunized with Pmy PI suggesting also the enhancement of Th2 response. It was clear that the native F. gigantica Pmy is considered as a relevant candidate for vaccination against fascioliasis. Also, these data suggested the immunoprophylactic effect of the native F. gigantica Pmy which is mediated by a mixed Th1/Th2 response.
PDF Abstract XML References Citation
Received: January 13, 2013;
Accepted: March 02, 2013;
Published: May 08, 2013
How to cite this article
H. Abou-Elhakam, I. Rabee, S. EL Deeb and A. EL Amir, 2013. Protection Against Fasciola gigantica using Paramyosin Antigen as a Candidate for Vaccine Production. Pakistan Journal of Biological Sciences, 16: 1449-1458.
DOI: 10.3923/pjbs.2013.1449.1458
URL: https://scialert.net/abstract/?doi=pjbs.2013.1449.1458
DOI: 10.3923/pjbs.2013.1449.1458
URL: https://scialert.net/abstract/?doi=pjbs.2013.1449.1458
INTRODUCTION
Fascioliasis is a parasitic disease caused by the digenetic trematodes, Fasciola gigantica (F. gigantica) (tropical liver fluke) and Fasciola hepatica (F. hepatica) (temperate liver fluke), leads to a significant economic loss. Fascioliasis reduces the production of meat, milk and wool, in addition to losses due to mortalities, liver condemnation and expenditures for anthelmintics using (Awad et al., 2009).
In Egypt fascioliasis, existed since Pharaonic ages (Soliman, 2008), about 830.000 people are infected in Egypt (World Health Organization, 1995). Treatment of the disease for human is World Health Organization major concern, using Triclabendazole (TCBZ) which is effective at single dose in Egypt (Soliman, 2008). Despite that chemotherapy with TCBZ is effective in treatment, it does not prevent reinfection. Therefore, animals in endemic countries must be continuously treated with the drug. This approach is prohibitively expensive for developing countries and furthermore, promotes the threat of drug resistance (Barduagni et al., 2008).
Thus, prevention and control of the disease is important to provide a proper treatment before liver damage occurs; this will be achieved by early diagnosis of the disease and development of a proper vaccine for prophylaxis from infection (Intapan et al., 2003). Several antigens are needed for efficient diagnostic methods and/or vaccine preparation. Most of these antigens are 97, 66, 58, 54, 47 and 14 kDa Mw and are derived from tegument and surface membrane which include glutathione-S-transferase, cysteine proteases, antioxidant enzyme, fatty acid binding protein, muscle paramyosin protein, as well as hemoprotein and membrane protein (Farahnak et al., 2010).
Paramyosin (Pmy) a fibrillar membrane protein widely distributed among invertebrates but absent in vertebrates, was defined as a potential candidate antigen to develop vaccines against some helminthiases, i.e., schistosomiasis and filariasis. Several vaccination trials are under way, or have been conducted, using either the native or recombinant protein. Different levels of response and protection have been achieved in animals (Cancela et al., 2004).
This study aims to study the humoral and cellular immune response for developing a vaccine using F. gigantica Pmy antigen which is isolated from adult worm homogenate during parasite invasion to give protection against this disease.
MATERIALS AND METHODS
Animals: Sixteen Newzealand white male rabbits, weighing approximately 1.5 kg and about one and half month of age, were examined before the experiments (free from Fasciola and other parasitic infection) and maintained at the Schistosome Biological Supply Program, Theodor Bilharz Research Institute, (SBSP/TBRI), Giza, Egypt. They were kept under standard laboratory care (at 21°C, 45-55% humidity) and supplied with filtered drinking water ad libitum, 24% protein and 4% fat diet. Animal experiments have been carried out according to the internationally valid guidelines and ethical conditions.
Parasites: Adult live F. gigantica worms were obtained from local abattoir. Fresh bovine livers were dissected and worms were collected from biliary tracts and gall bladders.
Metacercariae of F. gigantica were collected immediately in cellophane sheets after shedding from Lymnaea natalensis snails and then gently removed by scrapping them off with two small glass plates and used for infection.
Parasitological examination: Kato-Katz concentration technique (Martin and Beaver, 1968) and formal-ether sedimentation technique (Magambo et al., 1998) were performed for all stool samples in order to identify Fasciola eggs or other helminthic ova.
Preparation of Antigen
Preparation of F. gigantica whole worm homogenate: Adult clean F. gigantica worms were homogenized in 2 volumes (vol.) of 20 mM Tris-HCl buffer (BDH Chemicals, England) containing 5 mM Phenylmethylsulfonyl Fluoride (PMSF) as a protease inhibitor (Sigma-Aldrich, Louis, USA) at 20.000 rpm using IKA T20 homogenizer (IKA, Staufen, Germany). The homogenate was centrifuged at 30.000 rpm for 30 min. The entire process of homogenization and centrifugation was performed at 4°C. The supernatant fractions were decanted and assayed for protein content and stored at -20°C until used as crude extracts. Protein content of E/S antigen was measured by the Bio Rad protein assay kit (Bradford, 1976).
Purification of F. gigantica Pmy antigen from whole worm homgenate: Purification was carried out in two steps:
DEAE-Sephadex G-25 and G-200 ion exchange chromatography: Sephadex G-25 and G-200 powder (Amersham Bioscience, Uppsala, Sweden) was swelled in about 300 mL of 0.5 M Tris-HCl buffer (pH. 7). After swelling of the DEAE-Sephadex, the initial supernatant was removed and washed extensively with 10 mM Tris-HCl buffer (pH 6.5). Then sample was dialyzed versus the eluting buffer. Pmy is washed out of the gel and its protein content was calculated (Timanova et al., 1999).
SDS-page: Electrophoresis was performed using the Laemmli system (Laemmli, 1970), 12% SDS-PAGE (1 mm) under reducing condition according to Bio-Rad Lab. Model 595, Richmond, CA, USA manufacturer, followed by testing of F. gigantica Pmy antigen for reactivity and specificity by indirect ELISA (Engvall and Perlman, 1971) method.
Experimental design: In this study 16 rabbits were divided into four groups of four rabbits each:
| • | Group A: Uninfected control group |
| • | Group B: Infected control group; rabbits were infected orally with 25 F. gigantica metacercariae |
| • | Group C: Immunized group; rabbits were immunized with 1 mg of Pmy i.m. with equal vol. of CFA as 1ry immunization on day 0. Booster doses, 1 mL of 0.5 mg Pmy mixed with an equal vol. of IFA, were administered after 2, 3 and 4 week after the initial dose as 2nd, 3rd and 4th immunizations |
| • | Group D: Infected immunized group; rabbits were immunized by the same regimen as group C. Two week after the 4th immunization, each of these animals was challenged orally with 25 F. gigantica metacercariae (Muro et al., 1997) |
Blood samples were collected weekly from all groups during the immunization schedule and every 2 week after challenge infection from groups C and D till 12 week PI. Sera were prepared, heat-inactivated and kept at -20 °C till used.
Parasitological criteria
Fluke size and weight: All infected rabbits were euthanized in the 12th week PI. Individual livers were cut into slices approximately 1 cm thick and squeezed in warm saline for fluke collection. All flukes were recorded for their net weight, length and width. The size and net weight of flukes recovered from the immunized groups were compared with flukes recovered from the infected groups.
Bile egg count: Bile was collected only from all infected rabbits (control and immunized) were cleared by repeated sedimentation/decantation of the eggs. Egg counts were performed under a light microscope by measuring a definite vol. (10 μLx5) of each sample to calculate the total eggs in the given vol. of bile as in the standard protocol.
Immunological parameters: Assessment of anti-Fasciola IgM, total IgG and its isotypes IgG1, IgG2 and IgG4 by indirect ELISA based on the original method of Engvall and Perlman (1971) with the modifications of Nilsson (1990) and Venkatesan and Wakelin (1993).
Determination of cytokine profile using sandwich ELISA prepared plates (PharMingen, San Diego, Calif.). IL-6, IL-10, IL-12 and TNF-α levels in rabbit sera were elevated. Recombinant cytokines were used as standards according to Oliveira et al. (2010).
Statistics: According to Campbell (1989), results were evaluated using one-way analysis of variance (ANOVA). Comparison between two groups was done by the Student's t-test. The data were considered significant if p<0.05, highly significant if p<0.01 and very highly significant if p<0.001. Percent Reduction (PR) in all parameters was calculated according to the equation of Fonseca et al. (2004).
RESULTS
Purification and reactivity of F. gigantica Pmy: Figure 1 shows the OD280 profile of the adult F. gigantica worm Pmy antigen fractions obtained following purification of whole F. gigantica worm homogenate by DEAE Sephadex G-25 ion exchange chromatography. The eluted Pmy antigen as be represented by fractions (No. 6-14) with a single peak with maximum OD value equal to 2.903 at fraction number 12.
Antigen fractions were further purified by DEAE-Sephadex G-200 ion exchange chromatography. One peak was obtained representing the column elution volume fractions which contain Pmy with OD value 2.420 at fraction number 8 (Fig. 2).
Then eluted protein fraction was analyzed by 12% SDS-PAGE under reducing conditions showing only one band at 97 kDa representing Pmy (Fig. 3).
The antigenicity of the purified target Pmy antigen was tested by indirect ELISA technique. Serum samples from animals infected with F. gigantica gave strong reactivity against Fasciola Pmy antigen with mean OD reading equal to 1.317 and very weak cross reactions were recorded with sera from animals infected with other parasites e.g., S. mansoni, hydatid and hookworm.
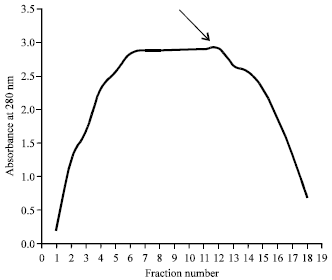 | |
| Fig. 1: | Profile of chromatography elution fractions of whole F. gigantica worm homogenate on DEAE-Sephadex G-25 ion exchange chromatography |
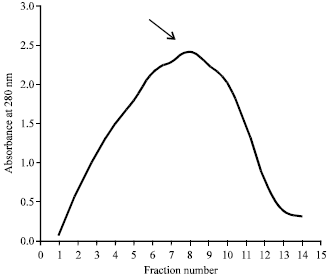 | |
| Fig. 2: | Profile of chromatography eluting fractions of whole worm homogenate on DEAE-Sephadex G-200 ion exchange chromatography |
Evaluation of F. gigantica Pmy immunization efficacy on some parasitological criteria
Effect on mean number of total worm burden: The mean total number of worm burden in rabbits infected with 25 metacercariae of F. gigantica was 12±1.1. Immunization of infected rabbits with Pmy (gr D) stimulated a very highly significant (p<0.001) decrease in mean total number of worm burden (5±1.3) with PR of 58.40%.
Effect on mean number of egg count: When Pmy immunized rabbits were infected with 25 F. gigantica metacercariae (gr D), the mean total number of egg load recorded was 3255±145, showing a very highly significant decrease (p<0.001) than infected control (gr B) (8440±213) with PR of 61.40%.
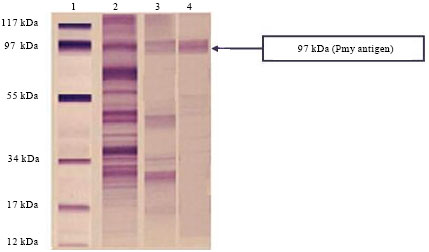 | |
| Fig. 3: | SDS-PAGE of target Pmy antigens eluted from affinity chromatography columns, Lane 1: Low molecular weight standard, Lane 2: Whole worm homogenate, Lane 3: Target Pmy antigen eluted from Sephadex G-25 and Lane 4: Purified Pmy antigen eluted from Sephadex G-200 |
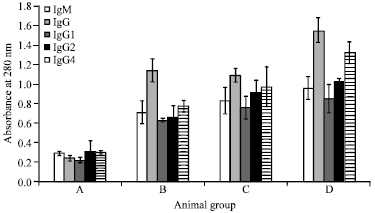 | |
| Fig. 4: | Effect of Pmy immunization on serum level of IgM, total IgG, IgG1, IgG2 and IgG4 in uninfected and infected rabbits with F. gigantica metacercariae, Columns depict Igs levels and vertical bars indicate the SD about the mean |
| Table 1: | Effect of immunization with Pmy on serum IgM, total IgG, IgG1, IgG2 and IgG4 levels in uninfected and infected rabbits with F. gigantica metacercariae |
 | |
| *Significant difference “p<0.05”, (a) Significant change with respect to uninfected control, **Highly significant difference “p<0.01”, (b) Significant change with respect to infected control, ***Very highly significant difference “p<0.001”, Data are represented as Mean±SD | |
Immunological criteria
Effect of Pmy on IgM and total IgG levels and its isotypes (IgG1, IgG2 and IgG4) in sera of different studied groups: Levels of IgM, IgG and their isotypes (IgG1, IgG2 and IgG4) of anti-Pmy injected in normal and infected rabbits were assayed using indirect ELISA (Fig. 4, Table 1).
IgM level: Results showed an expected increase in IgM level in infected control (gr B) (0.71± 0.12) in comparison to normal control (gr A) (0.29±0.02).
Immunization of rabbits with Pmy antigen (gr C) stimulated a significant (p<0.001) elevation of IgM level (0.83±0.14) in comparison to its normal level.
| Table 2: | Effect of immunization with Pmy on serum IL-6, IL-10, IL-12 and TNF-α level in infected and uninfected rabbits with F. gigantica metacercariae |
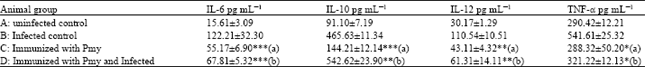 | |
| *Significant difference “p<0.05”, (a) Significant change with respect to uninfected control, **Highly significant difference “p<0.01”, (b) Significant change with respect to infected control, ***Very highly significant difference “p<0.001”, Data are represented as Mean±SD | |
After infection of preimmunized rabbits (gr D), IgM level was significantly (p<0.05) level increased (0.96±0.12), in comparison to infected control level.
Total IgG level: Infection itself causes an evident increase in the level of total IgG (1.14±0.12) (gr B) referring to normal uninfected rabbits (gr A) (0.24±0.03). Data revealed that, preinfected immunization of Pmy antigen infection (gr C) have an extraordinary significant (p<0.001) elevation of total IgG, than uninfected control group (1.09±0.07). On the other hand, immunization with Pmy preinfection increases the IgG level in serum (1.55±0.13).
IgG1 level: Infection alone (gr B) and Pmy immunization of rabbits induced a highly sig (p<0.01) elevation in IgG1 level (0.63±0.02 and 0.76±0.12, respectively) relative to normal uninfected control rabbits (gr A) (0.22±0.03). F. gigantica Infection of immunized rabbits (gr D) showed an extremely significant (p<0.01) elevation of IgG1 level (0.85±0.15) than the infected control (gr B).
IgG2 level: Again infection (gr B) caused a significant (p<0.001) elevation in the IgG2 level (0.66±0.12) and Pmy immunization (gr C) showed a very high significant (p<0.001) increase (0.91±0.13) in comparison to normal uninfected control group (0.31±0.11).
Serum IgG2 level of preimmunized infected rabbits (gr D) recorded 1.03±0.02, that considered the highest significant increase (p<0.05) (0.66±0.12).
IgG4 level: The level of IgG4 recorded a gradual Significant (p<0.01) increase from serum of normal rabbits (gr A) to infected (gr B), immunized (gr C) and preimmunized infected rabbits (gr D) as 0.30±0.02, 0.77±0.06, 0.97±0.02 and 1.32±0.11, respectively.
Effect of Pmy immunization on Th1 and Th2 cytokines level in sera of different studied groups: F. gigantica infection (gr B) induced a significant (p<0.01) elevation in the level of different cytokines as IL-6, IL-10, IL-12 and TNF-α (122.21±32.30, 465.63±11.34, 110.54±10.51 and 541.61±25.32, respectively) in comparison to normal level (gr A) (15.61±3.09, 91.10±7.19, 30.17±1.29 and 290.42±12.21, respectively) (Fig. 5, Table 2).
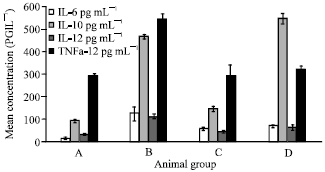 | |
| Fig. 5: | Effect of Pmy immunization on cytokine production level of IL-6, IL-10, IL-12 and TNF-α in uninfected and infected rabbits with F. gigantica metacercariae, Columns depict cytokine levels and vertical bars indicate the SD about the mean |
Immunization (gr C) alone detected a mild significant (p<0.01) increase in IL-6, IL-10 and IL-12 secretion level recording 55.17±6.9, 144.21±12.14 and 43.11±4.32, respectively. While, infection of immunized rabbits (gr D) induced a highly elevation in IL-10 (542.62±23.90), mild increase in IL-6 (67.81±5.32) and mild significant (p<0.01) decrease in IL-12 (61.31±14.11) (Fig. 5, Table 2).
On the other hand, TNF-α secretion level was affected by F. gigantica infection recording a significant elevation (541.61±25.32) referring to its normal level (290.42±12.21) (Fig. 5, Table 2).
Pmy immunization alone had no significant change on the level of TNF-α (288.32±50.2) while, its pre-infection injection protects the body by keeping the TNF-α level constant as normal (321.22±12.13) (Fig. 5, Table 2).
DISCUSSION
Fascioliasis, an infectious parasitic disease caused by F. hepatica or F. gigantica, affects millions of people worldwide. Up to 17 million people are infected and around 91.1 million are at risk of infection (Keiser and Utzinger, 2005). Today, fascioliasis is recognized as an emerging and re-emerging vector (ungulate)-borne disease with the widest latitudinal, longitudinal and altitudinal distribution known for any zoonotic disease. Hence, the World Health Organization has classified fascioliasis as an important human parasitic disease that merits international attention (Mas-Coma et al., 2005). Humans are often infected in communities where there is close human-ruminant interaction, such as in some South American communities, Egypt and Iran (Jayaraj et al., 2009).
Triclabendazole is the only effective drug against early stages of the parasite; however, resistance to it has been extensively reported (Villa-Mancera et al., 2008). Diagnosis in human is based on the findings the characteristic eggs in stool and duodenal fluids (Garcia, 2001). However, negative results do not rule out infection, especially in endemic areas or in field surveys undertaken for exposed agricultural community personnel (Sabry and Mohamed, 2007). Other control measures such as early diagnosis and vaccination should be developed for sustainable control of this disease that will be a cheaper, more efficient and reliable long term solution for the prevention of infection and eradication of its transmission.
The tegument of bile-dwelling F. gigantica is the interfacing layer that helps the parasite to maintain its homeostasis and evade the hostile environment, including the host's immune attacks. So, some studies reported that tegument can work as a basis for the development of immunodiagnosis and vaccine production (Sobhon et al., 1998).
This study was done to study the humoral and cellular immune response to Pmy antigen as an attempt to develop a vaccine against this disease. These aims were achieved through vaccination of rabbits by using Pmy to prevent the infection and to eradicate its transmission.
In the present study, the purification procedure of antigen was carried out by two steps: DEAE-Sephadex G-25 and G-200 ion exchange chromatography (Timanova et al., 1999). The yield was purified Pmy antigen with protein content equal to 2.6 mg mL-1, as a single band at 97 kDa by reducing SDS-PAGE. The results were reasonable with that of Cancela et al. (2004), who purified, characterized and immunolocalized the Pmy from adult stage of F. hepatica.
The target of this study is scheduling our protection strategy, where rabbits were divided into four groups; normal uninfected group, infected control group and Pmy immunized group, Pmy immunized and infected group.
Multiple Pmy immunizations of rabbits recorded a high significant reduction in worm burden (58.4%), as anti-F. gigantica vaccine development and also, had an adverse effect on the liver egg count (61.4%). Our findings are supported by Piacenza et al. (1999) who immunized sheep with native Cathepsin L1 or 2 (CL1 and CL2) against F. hepatica elicited 33 and 34% protection levels of worm burdens reduction and 71 and 81%, reduction in egg output, respectively while a cocktail of native CP (CL1 and CL2) induced 60% as PR in fluke burden. Gobert and McManus (2005) reviewed that, early Pmy vaccination against schistosomiasis can lead to impressive reduction of S. japonicum (Philippine strain) worm burdens in mice immunized with purified Pmy from adult worms. Also, Chen et al. (2000) reported that, cloned full length cDNA of S. japonicum Pmy used to generate a recombinant vaccine which reduced the worm burden with 50-60% in pigs. Jiz et al. (2008) stated that, murine immunization studies using both biochemically purified and recombinant Pmy of S. japonica and S. mansoni have consistently demonstrated significant protection from challenge infection as 24 to 56% and 32 to 86% reduction in worm burdens, respectively.
Ramajo et al. (2001), found no reduction in the worm burden in rFABP and native FABP-vaccinated sheep but there was a significant reduction in worm size and fecal egg counts, suggesting an anti-fecundity effect of the vaccine. Raina et al. (2004) also reported that, the antigenic stimulation by the parasite FABP to the host during the course of experimental/natural infection may not be sufficient to evoke detectable antibody level in ELISA and Western Blotting (WB), thereby suggesting that FABP as such is a weak antigen. Also, there is no significant humoral response generated against F. gigantica FABP in sheep, cattle and buffaloes.
However, immunization of sheep with CL proteinase derived peptide led to a significant decrease in challenge F. gigantica worm number and/or maturity (Villa-Mancera et al., 2008) and ability to produce eggs (Mulcahy et al., 1998), but failed to influence the size of the recovered worms (Jezek et al., 2008). The vaccine would have a profound effect on pasture contamination and hence disease transmission (Dalton et al., 2003).
The presented data obviously marked IgG level increase with infection and during Pmy immunization. While IgM level was increased (but lower than IgG level) by infection and all other treatments. Thus, it may be concluded that IgG plays a prominent role in combating infection where as IgM was not as effective.
Pmy immunization recorded highly significant enhancement of all Ig isotypes. While, in case of immunization with Pmy followed by infection, Pmy was less potent in elevating Ig isotypes than immunization only without infection.
Bossaert et al. (2000) also found that, IgG1 was significantly higher than IgG2 in calves with single-dose-infected of F. hepatica to E/S products and IgG2 level elevation was associated with protection (Mulcahy et al., 1998). It is conceivable therefore that, the late strong IgG2 found in this study does put cattle better placed to eliminate the liver fluke than sheep with early and weak IgG2. However, more work needs to be done to examine the contribution of IgG2 toward protection against liver fluke.
These results suggested that, predominance of IgG2 and IgG4 than IgG1 isotypes PI correlates with a predominance of IgG2-type Igs specific for Fasciola spp. antigens at different times PI. This is mostly characteristic of type 1 response which agrees with Girones et al. (2007) who detected antibody responses in rats in the acute and chronic phase (1-21 week) of disease as a marked predominance of IgG1 over IgG2a isotypes. During the 1stweek PI, IgG1 quickly increases, whereas, IgG2a slowly increases and reaches the highest values at 5-7 week PI. Phiri et al. (2006) suggested that, E/S antigens may preferentially stimulate a Th2 T cell subset response. The late IgG2 response to F. hepatica and F. gigantica E/S products in cattle may indicate a delayed Th1 T cell subset stimulation. Moreau et al. (1998) has previously shown that, the IgG2 response to a CL vaccine correlates with reduced worm burdens in vaccinated cattle, implying a protective role for this isotype against F. hepatica. These observations raise the interesting prospect that Fasciola parasites produce a factor (s) that can suppress IgG2 responses (2009).
Humans infected with F. hepatica develop IgM, IgA, IgE and IgG class. De Jesus et al. (2000) reported that, naturally occurring immunological responses to the S. mansoni antigens Pmy, IrV-5, Sm-23 (MAP-3) and triose phosphate isomerase (MAP-4), stimulated individually IgG1, IgG2, IgG3, IgG4 and IgA levels and the cytokine profile. Direct correlations between infection levels (number of epg of stool) and IgG1 and IgG4 levels to SWA and Pmy were observed suggesting that these isotypes are markers for high levels of infection while people with lower infection levels produced higher levels of IgG2 specific for these antigens. The data suggested that perhaps a vaccine that induces both Th1- and Th2-like responses would be necessary to achieve higher levels of protection.
Nara et al. (2007) suggested that, IgM, IgG2 and IgG4 initiated killing for schistosomiasis adult worms by antibody-dependant cell-mediated cytotoxicity (ADCC) that means it provokes an effector immune mechanism.
These results are in agreement with that of Zhou et al. (2000), who showed that pCMVSjc97 elicited much more IgG2a and IgG2b than IgG1 antibodies, whereas, in mice immunized with recombinant Sjp97 protein, IgG1 was the dominant IgG subclass. Moreover, examination of cytokine production patterns revealed high level IFN-γ and IL-2 production profiles by antigen-stimulated splenocytes from pCMV-Sjc97 inoculated C57BL/6 mice. It is widely agreed that the IgG subclasses of IgG2a and IgG2b and cytokine pattern of IFN-γ and IL-2 are indicative of Th1 type responses. Also, Vazquez-Talavera et al. (2001) used T. solium Pmy as a vaccine candidate to give protection against cysticercosis in murine model; it was found that the predominant antibody isotype was IgG1 subclass that reveals production of IFN-γ and IL-2, suggesting a Th1 like profile.
Lopez-Moreno et al. (2003), identified CD4+ T cell epitopes of Taenia solium Pmy and he found that immune response for TPmy-peptide 176 (Tp176) is Th1 response, where IgG2a increased in serum of immunized BALB/c mice.
In contrast, Zhao et al. (2007), study of Paragonimus westermani the predominant antibody isotypes against recombinant Pmy were IgG1 and IgG4 subclasses, suggesting that during chronic helminths infections, Th2 immune response is evoked. Also, Garcia et al. (2008) identified the Sm14 and Pmy epitopes that are recognized by T cells of resistant individuals living in endemic areas for schistosomiasis. It was found that protection was associated with a Th1 type of immune response induced by Sm14 peptide immunization while immune response induced by Pmy peptide was associated with Th2 type immune response.
On the other hand, results of this study clearly demonstrate that immunization with Pmy elevate secretion of type 1 cytokines (IL-6, IL-12 and TNF-α) and type 2 cytokines (IL-10 levels) than the normal control group. Administration of Pmy induces a high significant increase in IL-6 and IL-10 while, a significant increase in IL-12 and TNF-α level in immunized groups with IL-10 predominating. In contrast, there is a suppression in all cytokines level in rabbits immunized with Pmy and infected groups except IL-10 which was elevated than the infected control group. Thus, it could be hypothised that, Pmy was potent in increasing the levels of all cytokines understudy except IL-6 before infection. While, immunization with Pmy preinfection caused a decrease in all cytokines except for IL-10 which remained slightly but significantly elevated.
These data are supported by the idea of Molina (2005) who observed that, cattle and buffaloes infected with F. gigantica had a predominant Th2 response which started early in the infection. IL-6 production in these animals apparently influenced the initiation and maintenance of type 2 immune response, thereby down-regulating Th1 response. IL-6 and IL-8 (in buffaloes) during infection with F. gigantica may thus be capable of exerting a cytotoxic effect against the fluke.
IL-6 and IL-8 have been shown to be involved in ADCC involving neutrophils. In fascioliasis, ADCC has been considered to be a mechanism by which flukes are destroyed, with the priming of neutrophils, macrophages, eosinophils and mast cells by various cytokines (Hansen et al., 1999; Piedrafita et al. 2007). Therefore, cattle, buffalo and sheep, by producing IL-4, IL-6 and IL-8 during infection with F. gigantica, may be capable of exerting a cytotoxic effect against flukes.
In some of the infected rats, TNF-α also increased at 10 week PI which does not correspond to a typical Th2 response. TNF-α is implicated in the regulation of Th2 responses in other helminthes infections, apparently regulating worm expulsion. Moreover, IL-4 and IL-10 can act synergistically, inhibiting the production of reactive nitrogen oxides which up-regulate IL-12 production and inflammatory responses (Girones et al., 2007).
More recently, however, it is proposed that the secreted CLs may be involved in suppression and/or modulation of Th1 immune responses and induction of non-protective host Th2 responses (Dixit et al., 2008). An analysis of cytokine production by antigen-stimulated spleen cells of F. hepatica infected mice showed that these are predominantly of the Th2 type, i.e. production of IL-4, IL-5 and IL-10 but little or no IFN-γ (O'Neill et al., 2000). This is consistent with immunological observations in cattle which show that in the early stages of infections, mixed Th1/Th2 responses are observed but as infection progresses, a Th2 response predominates (Mulcahy et al., 1999).
CONCLUSION
In conclusion, these studies proved that native heterologous F. gigantica Pmy significantly protects rabbits against challenge infection with F. gigantica. multiple immunizations with native F. gigantica Pmy in Freund’s adjuvant resulted in high significant reduction in mean worm burden and liver egg count and elicit a high significant increase in IgM and IgG antibodies and more specifically IgG1, IgG2 and IgG4 isotypes with IgG4 isotype predominating. These data suggest that the immunoprophylactic effect of the native F. gigantica Pmy is mediated by a mixed Th1/Th2 response which indicated enhancement of immune response against fascioliasis infection.
REFERENCES
- Barduagni, P., Y. Hassanein, M. Mohamed, A.E. Wakeel, M.E. Sayed, Z. Hallaj and F. Curtale, 2008. Use of triclabendazole for treatment of patients co-infected by Fasciola spp. and Schistosoma mansoni in Behera Governorate, Egypt. Parasitol. Res., 102: 631-633.
PubMed - Bossaert, K., F. Farnir, T. Leclipteux, M. Protz, J.F. Lonneux and B. Losson, 2000. Humoral immune-response in calves to single-dose, trickle and challenge infections with Fasciola hepatica. Vet. Parasitol., 87: 103-123.
PubMed - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Cancela, M., C. Carmona, S. Rossi, B. Frangione, F. Goni and P. Berasain, 2004. Purification, characterization and immunolocalization of paramyosin from the adult stage of Fasciola hepatica. Parasitol. Res., 92: 441-448.
PubMed - Chen, H., T. Nara, X. Zeng, M. Satoh and G. Wu et al., 2000. Vaccination of domestic pig with recombinant paramyosin: Against Schistosoma japonicum in China. Vaccine, 18: 2142-2146.
CrossRef - De Jesus, A.R., I. Araujo, O.V. Bacellar, A. Magalhaes and E. Pearce et al., 2000. Human immune responses to Schistosoma mansoni vaccine candidate antigens. Infect. Immunity, 68: 2797-2803.
CrossRefDirect Link - Dixit, A.K., P. Dixit and R.L. Sharma, 2008. Immunodiagnostic/protective role of cathepsin L cysteine proteinases secreted by Fasciola species. Vet. Parasitol., 154: 177-184.
PubMed - Molina, E.C., 2005. Serum interferon-gamma and interleukins-6 and 8 during infection with Fasciola gigantica in cattle and buffaloes. Vet. Sci., 6: 135-139.
PubMed - Engvall, E. and P. Perlmann, 1971. Enzyme-Linked Immunosorbent Assay (ELISA) quantitative assay of immunoglobulin G. Immunochemistry, 8: 871-874.
CrossRefPubMedDirect Link - Farahnak, A., T. Golmohamadi and M.B.M. Rad, 2010. Carbohydrate detection and lectin isolation from tegumental tissue of Fasciola hepatica. Iran. J. Parasitol., 5: 20-24.
Direct Link - Garcia, T.C., C.T. Fonseca, L.G. Pacifico, V. Duraes Fdo and F.A. Marinho et al, 2008. Peptides containing T cell epitopes, derived from Sm 14, but not from paramyosin, induce a Th1 type of immune response, reduction in liver pathology and partial protection against Schistosoma mansoni infection in mice. Acta Trop., 106: 162-167.
PubMed - Girones, N., M.A. Valero, M.A. Garcia-Bodelon, I. Chico-Calero, C. Punzon, M. Fresno and S. Mas-Coma, 2007. Immune suppression in advanced chronic fascioliasis: an experimental study in a rat model. J. Infect. Dis., 195: 1504-1512.
PubMed - Gobert, G.N. and D.P. McManus, 2005. Update on paramyosin in parasitic worms. Parasitol. Int., 54: 101-107.
CrossRef - Hansen, D., D. Clery, S. Estuningsih, S. Widjajanti, S. Partoutomo and T. Spithill, 1999. Immune responses in Indonesian thin tail and Merino sheep during a primary infection with Fasciola gigantica: Lack of a specific IgG2 antibody response is associated with increased resistance to infection in Indonesian sheep. Int. J. Parasitol., 29: 1027-1035.
PubMed - Intapan, P.M., W. Maleewong, S. Nateeworanart, C. Wongkham, V. Pipitgool, V. Sukolapong and S. Sangmaneedet, 2003. Immunodiagnosis of human fascioliasis using an antigen of Fasciola gigantica adult worm with molecular mass of 27 kDa by a dot-ELISA. Southeast Asian J. Trop. Med. Public Health, 34: 713-717.
Direct Link - Jayaraj, R., D. Piedrafita, K. Dynon, R. Grams, T.W. Spithill and P.M. Smooker, 2009. Vaccination against fasciolosis by a multivalent vaccine of stage-specific antigens. Vet. Parasitol., 160: 230-236.
CrossRef - Jiz, M., H.W. Wu, R. Meng, S. Pond-Tor and M. Reynolds et al., 2008. Pilot-scale production and characterization of paramyosin, a vaccine candidate for Schistosomiasis japonica. Infect. Immunity, 76: 3164-3169.
CrossRefDirect Link - Keiser, J. and J. Utzinger, 2005. Emerging foodborne trematodiasis. Emerg. Infect. Dis., 11: 1507-1514.
CrossRefPubMedDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Magambo, J.K., E. Zeyhle and T.M. Wachira, 1998. Prevalence of intestinal parasites among children in southern Sudan. East Afr. Med. J., 75: 288-290.
PubMed - Martin, L.K. and P.C. Beaver, 1968. Evaluation of Kato thick smear technique for quantitative diagnosis of helminth infections. Am. J. Trop. Med. Hyg., 17: 382-391.
PubMed - Mas-Coma, S., M.D. Bargues and M.A. Valero, 2005. Fascioliasis and other plant-borne trematode zoonoses. Int. J. Parasitol., 35: 1255-1278.
CrossRefDirect Link - Moreau, E., A. Chauvin and C. Boulard, 1998. IFN-gamma and IL-10 production by hepatic lymph node and peripheral blood lymphocytes in Fasciola hepatica infected sheep. Parasite, 5: 307-315.
PubMed - Nara, T., K. Hzumi, H. Ohmae, O.S. Sy and S. Tsubota et al., 2007. Antibody Isotype Responses to Paramyosin, A Vaccine Candidate for Schistosomiasis and Their Correlations with Resistance and Fibrosis in Patients Infected with Schistosoma japonicum in Leyte, The Philippines. Am. J. Trop. Med. Hyg., 76: 384-391.
PubMed - Phiri, A.M., I.K. Phiri, C.S. Sikasunge, M. Chembensofu and J. Monrad, 2006. Comparative fluke burden and pathology in condemned and noncondemned cattle livers from selected abattoirs in Zambia. Onderstepoort. Vet. Res., 73: 275-281.
PubMed - Piacenza, L., D. Acosta, I. Basmadjian, J.P. Dalton and C. Carmona, 1999. Vaccination with cathepsin L proteinases and with leucine aminopeptidase induces high levels of protection against fascioliasis in sheep. Infect Immun., 67: 1954-1961.
PubMed - Piedrafita, D., E. Estuningsih, J. Pleasance, R. Prowse, H.W. Raadsma, E.N.T. Meeusen and T.W. Spithill, 2007. Peritoneal lavage cells of Indonesian thin tail sheep mediate antibody-dependent superoxide radical cytotoxicity in vitro against newly excysted juvenile Fasciola gigantica but not juvenile Fasciola hepatica. Infect. Immunity, 75: 1954-1963.
CrossRef - Ramajo, V., A. Oleaga, P. Casanueva, G.V. Hillyer and A. Muro, 2001. Vaccination of sheep against Fasciola hepatica with homologous fatty acid binding proteins. Vet. Parasitol., 97: 35-46.
CrossRefPubMedDirect Link - Sobhon, P., S. Anantavara, T. Dangprasert, V. Viyanant and D. Krailas et al., 1998. Fasciola gigantica: Studies of the tegument as a basis for the developments of immunodiagnosis and vaccine. Southeast Asian J. Trop. Med. Publ. Health, 29: 387-400.
PubMed - Soliman, M.F.M., 2008. Epidemiological review of human and animal fascioliasis in Egypt. J. Infect. Dev. Countries, 2: 182-189.
CrossRefDirect Link - Vazquez-Talavera, J., C.F. Solis, L.I. Terrazas and J.P. Laclette, 2001. Characterization and protective potential of the immune response to Taenia solium paramyosin in a murine model of cysticercosis. Infect. Immunity, 69: 5412-5416.
CrossRefDirect Link - Venkatesan, P. and D. Wakelin, 1993. ELISAs for parasitologists: Or lies, damned lies and ELISAs. Parasitol. Today, 9: 228-232.
CrossRefPubMedDirect Link - Villa-Mancera, A., H. Quiroz-Romero, D. Correa, F. Ibarra and M. Reyes-Perez et al., 2008. Induction of immunity in sheep to Fasciola hepatica with mimotopes of cathepsin L selected from a phage display library. Parasitology, 135: 1437-1445.
CrossRefDirect Link - Zhou, S., S. Liu, G. Song, Y. Xu and W. Sun, 2000. Protective immunity induced by the full-length cDNA encoding paramyosin of Chinese Schistosoma japonicum. Vaccine, 18: 3196-3204.
CrossRef