
I.K. Olayemi
Department of Biological Sciences, Federal University of Technology, Minna, Nigeria
V.O. Ojo
Department of Biological Sciences, Federal University of Technology, Minna, Nigeria
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 3 | Page No.: 135-140







ABSTRACT
This study elucidated the relationships between larval habitat soil-substrate Organic Matter Content (OMC) and immature development of the mosquito Anopheles gambiae s.l. Day-old larvae of the mosquito were reared in media substrated with typical soil samples (i.e., sandy, silt, clayey and loamy soils), from established anopheline breeding sites, to provide a gradient in soil-substrate OMC. The OMC of the soil samples were determined by ignition to a constant weight; while the developing A. gambiae mosquitoes in the culture media were monitored daily for survivorship and duration of immature life stages. The results indicated significant (p<0.05) variation in OMC of the soil types (range = 11.21±2.91% in sandy to 29.83±2.96% in loamy soils). However, though Daily Larval Survival Rates (DLSR) were relatively high (range = 95.21±2.96 to 96.70±1.44%), as influenced by OMC, such values were not significantly different (p>0.05) among the soil-substrate types; results contrary to those of Larval Success Rates (LSR) (i.e., range = 52.07±13.64 to 74.39±6.60%). Daily Pupation Rate (DPR) of the mosquitoes varied significantly among the soil-substrates, ranging from 13.87±2.39% in clayey to 25.00±4.30% in loamy substrates. Soil-substrate OMC significantly extended the Duration of Immature Life Stages (DILS) of the mosquitoes only in the sandy soil type (range = 12.76±1.74 to 15.81±2.40 days). On the whole, DILS was inversely related to soil-substrate OMC. Cross-correlational analysis revealed significant positive association among most of the variables tested. The findings of this study should serve as baseline information for the development of effective environmental management strategies for malaria larval-vector control.
PDF Abstract XML References Citation
Received: December 05, 2012;
Accepted: February 13, 2013;
Published: March 16, 2013
How to cite this article
I.K. Olayemi and V.O. Ojo, 2013. Immature Development of the Malaria Vector Mosquito, Anopheles gambiae S.L. (Diptera: Culicidae), in Relation to Soil-substrate Organic Matter Content of Larval Habitats in Northcentral Nigeria. Pakistan Journal of Biological Sciences, 16: 135-140.
DOI: 10.3923/pjbs.2013.135.140
URL: https://scialert.net/abstract/?doi=pjbs.2013.135.140
DOI: 10.3923/pjbs.2013.135.140
URL: https://scialert.net/abstract/?doi=pjbs.2013.135.140
INTRODUCTION
Anopheles gambiae is the principal vector of malaria in sub-Saharan Africa, especially Nigeria (WHO, 1975; Olayemi and Ande, 2008). Malaria exerts heavy health burden on people in the region and has significantly undermined the socio-economic development of Africa (Leighton and Foster, 1993; Federal Ministry of Health, 2000; Roll-Back-Malaria, 2002; WHO, 2007). The intensity of malaria transmission in an area depends largely on population density of the mosquito vectors which, in turn, is significantly influenced by the rate and success of immature stage development (Olayemi, 2009). According to Pfaehler et al. (2006), the quality of anopheline mosquito larval habitats correlates with the nutritional conditions of larval development and may strongly influence the distribution and abundance of malaria vector mosquitoes. As a result, larval habitat selection by anopheline mosquitoes is by active choice (Bentley and Day, 1989; Lyimo et al., 1992; Blaustein et al., 2004), often dictated by the need to secure sites that will support safe immature development in the shortest time (Minakawa et al., 2005; Kershenbaum et al., 2012).
The major components of anopheline larval habitats are the water medium and soil substratum. Understanding the influence of quality of habitat on immature survival and development of mosquitoes has being disproportionately focused on the water media. To this end, the physico-chemical characteristics of anopheline larval habitats, in relation to distribution and abundance of larvae, have been well elucidated (Robert et al., 1998; Gimnig et al., 2001; Oo et al., 2002). Yet, the soil substrate of mosquito larval habitats, especially the total organic matter content is of critical importance in the availability of particulate and microbial larval diet, for rapid and successful development. Studies have shown that, as a result of the small-sized and temporary nature of anopheline larval habitats, due to seasonal variations (Li et al., 2009), the physico-chemical properties of the water media of such habitats change rapidly (Piyaratne et al., 2005; Mwangangia et al., 2007; Troyo et al., 2008) and hence, may be less impacting on larval development. The soil substrate on the other hand, is more permanent and serves as the reservoir of chemo-biological components of the habitats especially during the dry season (Pfaehler et al., 2006).
Therefore, adequate understanding of the bio-ecological factors that condition the abundance and distribution of anopheline larvae, a pre-requisite for effective mosquito larval control, may not be achieved without a detailed evaluation of the influence of soil-substrate organic matter content on larval developmental and survival rates. Unfortunately, this aspect of anopheline larval ecology has been neglected for long, resulting in a dearth of information on the subject. In order to fill this gap in literature, this study was carried out to elucidate the relationships between soil-substrate organic matter content of anopheline larval habitats and immature development and survival rates.
MATERIALS AND METHODS
Study area: The study was carried out in Minna, the capital city of Niger state, north central Nigeria. Minna is located within longitude 6°33 E and latitude 9°27 N, covering a land area of 88 km2 with an estimated human population of 1.2 million. The area has a tropical climate with mean annual temperature, relative humidity and rainfall of 30.20°C, 61.00% and 1334.00 mm, respectively. The climate presents two distinct seasons: a rainy season between May and October, followed by a dry season (November - April). The vegetation in the area is typically grass-dominated savannah with scattered trees. The soil types range from excessively drained sandy to water-logged loamy/clay soils.
Malaria is endemic throughout the year in Minna, with relatively high prevalence rate (Olayemi et al., 2009; Omalu et al., 2012). The dominant malaria vector in the area is Anopheles gambiae (Olayemi and Ande, 2008) and the conventional anopheline larval breeding habitats include drains and temporary rain pools (Olayemi et al., 2010, 2012).
Soil collection and rearing of mosquito larvae on soil substrates: Previously known high-potential Anopheles gambiae natural breeding habitats in the study area (Olayemi and Ande, 2009; Olayemi et al., 2012), were selected for sample collection to represent the major types of soil namely, sandy, loamy, silt and clayey; to provide a gradient of soil-substrate organic matter content. The soil samples were collected from the habitats, within 5cm of the surface and were transported to the laboratory in plastic bowls (25 cm diameter and 15 cm deep), where they were flooded with bore-hole water and the top of the bowls were screened with mosquito netting, to induce hatching of wild mosquito eggs and isolate inherent immature stages for elimination. After flooding for three days, the soil samples were drained and air-dried for 2 weeks in a screened chamber to prevent wild mosquitoes from laying eggs on the soil substrates. Subsequently, 150 g of each type of soil sample were placed in four replicate larval-rearing troughs, flooded with two litres of bore-hole water and screened with mosquito, netting to exclude oviposition by wild mosquitoes. A Control experiment was also set-up in four replicates, but consisting of only 2 L of water in each trough and properly screened. The larval rearing troughs with their contents were then allowed to stand for three days, to ascertain total exclusion of wild mosquitoes, as well as condition the water media for larval rearing.
Thereafter, 50 approximately day-old first instar larvae of Anopheles gambiae s.l. mosquitoes from a colony maintained in the Laboratory of the Department of Biological Sciences, Federal University of Technology, Minna, Nigeria, were introduced to each larval-rearing trough. No extra larval food was added to the troughs and the experiment was monitored daily for mosquito immature survivorship and duration of development. The whole experiment was repeated within two weeks of terminating the first, resulting in the monitoring of 400 larvae per soil-substrate type.
Mortality rates and duration of the immature life stages were determined according to the methods of Olayemi and Ande (2009).
Daily Larval Survival Rate (DLSR) was determined by taking note of the number of larvae alive on the next day of a previous observation.
Daily Pupation Rate (DPR) was calculated as the number of larvae that metamorphosed into pupae on a daily basis.
Larval and Pupal success rates were calculated using the formula:
Si = ni/(Xni-1)x100
where, Si is success rate of a life stage; ni is number of individuals entering a life stage; Xni-1 is number of individuals that entered the preceding life stage.
Aggregate Immature Success Rate (AISR) was determined as mean larval and pupal success rates.
For the estimation of duration of larval and pupal life stages, the following formula was used:
Di = Ti - (ti-1)
where, Di is duration of a life stage; Ti is mean age at present metamorphosis; ti-1 is mean age at previous metamorphosis.
Aggregate Immature Duration (AID) was calculated as the sum of larval and pupal development durations.
Determination of soil organic matter content: The Organic Matter Contents (OMC) of the soil-substrate types used for culturing the mosquito larvae, was determined following standard procedures (Pfaehler et al., 2006; Okech et al., 2007). Briefly, four replicates of 10 g dry ground samples of each soil-substrate type was placed in a crucible and ignited to a constant weight, using a Bunsen flame. The OMC was estimated as the difference in the weight of soil sample and crucible before and after burning.
Statistical analysis: Mean edaphic and entomological variables among the soil-substrate types were compared for statistical significance using the Chi-square test, at p = 0.05. The relationships between the variables investigated were assessed using Linear Correlation Coefficient.
RESULTS
Organic matter contents of soil substrates: The effects of differential Soil-substrate Organic Matter Content (SOMC) of breeding habitats on the survivorship and metamorphosis of immature life stages of Anopheles gambiae in the study area are highlighted in Table 1. The Control experiment (i.e., bore-hole water only) was totally devoid of organic matter. However, the larval habitat soil-substrate types investigated contained significantly (p<0.05) variable amounts of organic matter, ranging from 11.21±2.91% in sandy to 29.83±2.96% in loamy soils. The organic matter content of the clayey and silt substrates (i.e., 16.43±4.70 and 14.58±6.83%, respectively) were not significantly different (p>0.05).
Effect of soil substrate organic matter content on survivorship of immature stages: Daily Larval Survival Rates (DLSR) were relatively high and not significantly different among the soil-substrates (range = 95.21±2.96 to 96.70±1.44%), though it was significantly lower in the Control mosquitoes (86.30±9.48%) (Table 1). On the other hand, Larval Success Rates (LSR) varied significantly among the soil substrates and no larva pupated in the control experiment. LSR responded proportionally to increase in organic matter content of soil substrates, ranging from 52.07±13.64 to 74.39±5.60% in sandy and loamy soil substrates, respectively. The patterns of distribution of pupal survivorship attributes among the soil substrates were distinctly different from those of the larval stage. For example, unlike DLSR, Daily Pupation Rate (DPR) of the mosquitoes varied significantly among the soil substrates, ranging from 13.87±2.39% in clayey substrate to 25.00±4.30% in loamy soil. Also, while LSR varied significantly, Pupal Success Rate (PSR) on other hand were mostly insignificantly (p>0.05) different among the soil-substrate types. However, the success rate of the aggregate immature stage of the mosquito species was significantly reduced by decreasing soil-substrate organic matter content. The most productive soil-substrate type was loamy, with 87.20±9.11% of larvae reared in this medium successfully emerging as adult mosquitoes while, the least was sandy soil (72.19±15.86%) (Table 1).
Effect of soil substrate organic matter content on duration of immature stages: Table 2 shows the effects of soil-substrate organic matter content on Duration of Immature Life Stages (DILS) of the mosquitoes. DILS was only significantly (p<0.05) extended in the sandy soil-substrate (range = 12.76±1.74 to 15.81±2.40 days). The patterns of distribution of duration of the larval and pupal stages were separately different from the aggregate immature stage, as well as from each other. The duration of larval stage was significantly shortest in clayey soil substrate (6.48±2.75 days) and longest (9.69±1.89 days) among the larvae raised in sandy soil-substrate medium. For the pupal stage, the duration of development though significantly shortest in the loamy substrate (4.30±0.54 days), was insignificantly different in the other three soil-substrate types (range = 5.59±0.29 to 7.00±1.28 days). Surprisingly, unlike larval and pupal success rates, the quality of the duration of these immature life stages did not follow the definite pattern of improvement with increase in organic matter content.
| Table 1: | Mean indices of immature survivorship response of Anopheles gambiae mosquitoes to soil substrate organic matter content of larval breeding habitats in Northcentral Nigeria |
 | |
| *Values followed by same superscript alphabets in a column are not significantly different at p<0.05, **Not applicable, due to 100% mortality recorded at the larval stage | |
| Table 2: | Mean duration (days) of immature life stages of Anopheles gambiae in response to soil-substrate organic matter content of larval breeding habitats in Northcentral Nigeria |
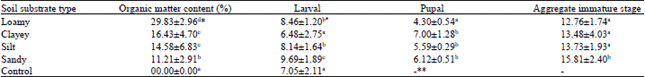 | |
| *Values followed by same superscript alphabets in a column are not significantly different at p = 0.05, **Not applicable, due to 100% mortality recorded at the larval stage | |
| Table 3: | Cross-correlation among immature survival and developmental rates of Anopheles gambiae in response to soil-substrate organic matter content of larval breeding habitats in Northcentral Nigeria |
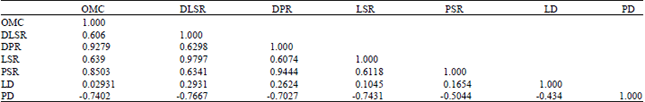 | |
| OMC: Organic matter content, PSR: Pupal success rate, LSR: Larval success rate, DLSR: Daily larval survival rate, LD: Larval duration (days), DPR: Daily pupation rate and PD: Pupal duration | |
Cross-correlation among indices of immature duration and survivorship, in response to soil substrate organic matter content: Cross-correlations among the indices of survival and developmental rates, in response to soil-substrate organic matter content of larval breeding habitats, are presented in Table 3. Strong positive correlations (r = 0.6000) existed among most of the variables tested. However, correlations between Larval Duration (LD) and all other variables were very weak (r<0.3000). Distinctly, correlations between Pupal Duration (PD) and other variables though, mostly strong (r>0.7000) except with Pupal Survival Rate (PRS) and LD (r<0.6000)0 were negative.
DISCUSSION
Organic Matter Content (OMC) varied significantly among the soil-substrate types and followed the conventional order of abundance in soil types. Generally, loamy soils are highly rich in humus (a precursor of soil organic matter) followed by silt and clayey while, sandy soils contain the least amount of humus, due to its high vulnerability to drainage. The OMC of mosquito larval habitat soil-substrates obtained in this study (range = 11.21±2.91 to 29.83±2.96%), were relatively higher than the 3.61 - 21.25% reported from Western Kenya (Pfaehler et al., 2006). Such variations may be due to differential locational vegetation cover and anthropogenic activities. However, the generally high OMC of anopheline larval habitat soil-substrate suggest increased productivity of such habitats in the study area.
While, Daily Larval Survival Rate (DLSR) was generally high and insignificantly different among soil-substrate types, the reverse was the case for Larval Success Rate (LSR), which also responded proportionally to increase in soil-substrate Organic Matter Content (OMC). This finding indicates that DLSR and LSR respond differentially to OMC of anopheline larval habitats. This difference in responses to OMC may be explained by the aspect of larval survivorship emphasized by the two variables (i.e., DLSR and LSR). While, DLSR focuses on the proportion of larvae that survive to the following day, LSR is an estimate of the proportion of larvae that successfully transform to pupa. DLSR may not be significantly affected by OMC, as such form of survivorship does not require much nutrient (supplied by organic matter) especially during the first 2 of the 4 larval instars; but requires more of genetic vigour and absence of predators (Blaustein et al., 2004); both factors favoured by the rearing of mosquito larvae under laboratory conditions, as carried out in this study. On the other hand, successful metamorphosis of mosquito larvae to pupae, as indicated by LSR, requires rapid and elaborate assimilation of teneral reserve (Timmermann and Briegel, 1999; Briegel, 2003), derivable principally from nutrients directly or indirectly supplied by soil-substrate organic matter of larval habitats (Wotton et al., 1997). The results of this study showed that larval and pupal survivorships of A. gambiae responded differentially to OMC of the soil-substrates of breeding habitats. This finding, perhaps, underscores the differential biological and environmental requirements of the two mosquito immature life stages (i.e., larval and pupal), as previously reported (Telang et al., 2007; Araujo et al., 2012).
The more-or-less insignificantly different distribution of Pupal Success Rates among the soil-substrate types is, however, not surprising. Being a non-feeding stage (Olayemi and Ande, 2009), mosquito pupal development and eventual metamorphosis to imago may not be affected by a nutrition-contributory factor such as soil-substrate Organic Matter Content (OMC) of breeding habitats. From the results of this study, it seems that the effects of OMC on immature development of A. gambiae either terminated at the larval stage or, perhaps, the larvae that successfully pupated were able to overcome any mitigating effects of differential OMC observed in the larval stage. However, adult teneral reserve in mosquitoes is largely dictated by food assimilation, acquired during the late larval stage (Briegel, 2003). Therefore, there is need for further studies to elucidate the influence of differential soil-substrate OMC on adult fitness and teneral reserve of adult mosquitoes, in order to confirm if such influence truly terminates at the larval stage.
The success rate of the aggregate immature stage was significantly influenced by soil-substrate OMC; results consistent with earlier observations on mosquito breeding ecology (Lee et al., 1991; Thiery et al., 1993; Wotton et al., 1997; Alahmed, 1998; Pfaehler et al. (2006), indicating that organic matter in mosquito breeding habitats enhanced growth of dietary micro-organisms, accelerated larval development and survival. The mean aggregate immature success rates obtained in this study (range = 72.19-87.20%) agreed with that reported for the same mosquito s pecies (mean immature survival rate = 81.26%), from a laboratory study on the life-table of A. gambiae in the same eco-geographical zone, during which the larvae were fed with the fish feed, Tetramin (Olayemi and Ande, 2009). The anopheline larvae raised in this study depended on the organic matter inherent in the soil-substrate of the culture media for nutrition, yet achieved immature stage success rate similar to that of larvae fed with the highly nutritious fish feed. This observation, therefore, suggests that the organic matter component of larval habitat soil-substrate provides a rich supply of mosquito larval dietary requirements for optimum development. Anopheline mosquito larval control programs must, therefore, take in to serious consideration, the organic matter richness of breeding habitats while, prioritising such sites for larviciding interventions.
CONCLUSION
Anopheline mosquito breeding habitats in Minna differed significantly in soil-substrate Organic Matter Content, which proportionally influenced mosquito immature success rate; thus, probably, affecting the density and distribution of malaria vectors in the area. However, there is need to elucidate the influence of larval habitat soil-substrate OMC on the vectorial fitness of the emerging adult mosquitoes. The findings of this study, therefore, underscore the need for the inclusion of environmental management strategies that will reduce the OMC of larval habitat soil-substrate, for effective malaria vector mosquito larviciding activities.
REFERENCES
- Araujo, M.S., L.H.S. Gil and A.A. e-Silva, 2012. Larval food quantity affects development time, survival and adult biological traits that influence the vectorial capacity of Anopheles darlingi under laboratory conditions. Malar. J., Vol. 11.
CrossRef - Alahmed, A.M.N., 1998. The effect of various manure suspensions (camel, cow and sheep) on the life cycle of Culex pipiens. Saudi J. Biol. Sci., 5: 58-63.
Direct Link - Bentley, M.D. and J.F. Day, 1989. Chemical ecology and behavioral aspects of mosquito oviposition. Annu. Rev. Entomol., 34: 401-421.
CrossRef - Blaustein, L., M. Kiawi, A. Eitam, M. Mangel and J.E. Cohen, 2004. Oviposition habitat selection in response to risk of predation in temporary pools: Mode of detection and consistency across experimental venue. Oecologia, 138: 300-305.
CrossRef - Briegel, H., 2003. Physiological bases of mosquito ecology. J. Vector Ecol., 28: 1-11.
PubMedDirect Link - Gimnig, J.E., M. Ombok, L. Kamau and W.A. Hawley, 2001. Characteristics of larval anopheline (Diptera: Culicidae) habitats in Western Kenya. J. Med. Entomol., 38: 282-288.
PubMedDirect Link - Kershenbaum, A., M. Spencer, L. Blaustein and J.E. Cohen, 2012. Modelling evolutionarily stable strategies in oviposition site selection, with varying risks of predation and intraspecific competition. Evol. Ecol., 26: 955-974.
CrossRef - Li, L., L. Bian, L. Yakob, G. Zhou and G. Yan, 2009. Temporal and spatial stability of Anopheles gambiae larval habitat distribution in Western Kenya highlands. Int. J. Health Geogr., Vol. 8.
CrossRefDirect Link - Minakawa, N., G. Sonye and G. Yan, 2005. Relationships between occurrence of Anopheles gambiae S. l. (Diptera: Culicidae) and size and stability of larval habitats. J. Med. Entomol., 42: 295-300.
PubMed - Mwangangia, J.M., C.M. Mbogo, E.J. Muturi, J.G. Nzovu and J.I. Githureb et al., 2007. Spatial distribution and habitat characterisation of Anopheles larvae along the Kenyan coast. J. Vector Borne Dis., 44: 44-51.
Direct Link - Okech, B.A., L.C. Gouagna, G. Yan, J.I. Githure and J.C. Beier, 2007. Larval habitats of Anopheles gambiae s.s. (Diptera: Culicidae) influences vector competence to Plasmodium falciparum parasites. Malar. J., Vol. 6.
CrossRefDirect Link - Olayemi, I.K. and A.T. Ande, 2008. Relative abundance and seasonal distribution of adult mosquito species in Offa, Kwara State, Nigeria. Niger. J. Entomol., 25: 47-52.
Direct Link - Olayemi, I.K. and A.T. Ande, 2009. Life table analysis of Anopheles gambiae (diptera: culicidae) in relation to malaria transmission. J. Vector-borne Dis., 46: 295-298.
PubMedDirect Link - Olayemi, I.K., B. Idris, I.C.J. Omalu and M.O. Odeyemi, 2012. Dry season refugia breeding ecology of mosquitoes (Diptera: Culicidae) in Minna, North Central Nigeria. J. Biol. Sci., 12: 186-191.
CrossRefDirect Link - Omalu, I.C.J., C. Mgbemena, A. Mgbemena, V. Ayanwale, I.K. Olayemi, L.A. Adeniran and V. Chukwuemeka, 2012. Prevalence of congenital malaria in Minna, North Central Nigeria. J. Trop. Med.
CrossRefDirect Link - Oo, T.T., V. Storch and N. Becker, 2002. Studies on the bionomics of Anopheles dirus (Culicidae: Diptera) in Mudon, Mon State, Myanmar. J. Vector Ecol., 27: 44-54.
Direct Link - Pfaehler, O., D.O. Oulo, L.C. Govagana, J. Githure and P.M. Guerin, 2006. Influence of soil quality in the larval habitat on development of Anopheles gambiae Giles. J. Vector Ecol., 31: 400-405.
CrossRefDirect Link - Piyaratne, M.K., F.P. Amerasinghe, P.H. Amerasinghe and F. Konradsen, 2005. Physico-chemical characteristics of Anopheles culicifacies and Anopheles varuna breeding water in a dry zone stream in Sri Lanka. J. Vector Borne Dis., 42: 61-67.
Direct Link - Robert, V., H.P. Awono-Ambene and J. Thioulouse, 1998. Ecology of larval mosquitoes, with special reference to Anopheles arabiensis (Diptera: Culcidae) in market-garden wells in urban Dakar, Senegal. J. Med. Entomol., 35: 948-955.
PubMed - Telang, A., L. Frame and M.R. Brown, 2007. Larval feeding duration affects ecdysteroid levels and nutritional reserves regulating pupal commitment in the yellow fever mosquito Aedes aegypti (Diptera: Culicidae). J. Exp. Biol., 210: 854-854.
CrossRefDirect Link - Timmermann, S.E. and H. Briegel, 1999. Larval growth and biosynthesis of reserves in mosquitoes. J. Insect Physiol., 45: 461-470.
CrossRefDirect Link - Troyo, A., O. Calderon-Argedas, D.O. Fuller, M.E. Solano and A. Avendano et al., 2008. Seasonal profiles of Aedes aegypti (Diptera: Culicidae) larval habitats in an urban area of Costa Rica with a history of mosquito control. J. Vector Ecol., 33: 76-88.
Direct Link - Wotton, R.S., D.T. Chaloner, C.A. Yardley and R.W. Merritt, 1997. Growth of Anopheles mosquito larvae on dietary microbiota in aquatic surface microlayers. Med. Vet. Entomol., 11: 65-70.
CrossRef