
Kartikeya Tiwari
Microbial Biotechnology Laboratory, Jaipur National University, Jaipur, India-302017, India
Pakistan Journal of Biological Sciences
Year: 2013 | Volume: 16 | Issue: 19 | Page No.: 1004-1009







ABSTRACT
The present study discusses the frequency distribution and genetic diversity of novel fungal endopyte Alternaria alternata within the Pongammia pinnata plant samples. A total of ten plant samples of Pongammia pinnata, Pierre. (Karanja) were collected from specific locations of Sanganer region of Rajasthan for the isolation of fungal endophytes. Of these, maximum frequency of Alternaria alternata (22.29%) were recorded which are morphologically similar but ecologically variant. Efficacy of randomly amplified polymorphic DNA (RAPD), were assessed in seventeen individuals of the primers was GCC 180 where as 10 bands were generated by GCC 181. The similarity coefficient matrix generated for the primers was subjected to algorithm UPGMA (Unweighted Pair Group Method Analysis) and clusters were generated using NTSYS 2.02 pc program. To stabilize the level of relatedness among the seventeen ecologically variant Alternaria alternata accessions, the dendrogram was constructed, which showed that all the isolates were diversified endophytically with in the plant Pongamia pinnata.
PDF Abstract XML References Citation
Received: January 12, 2013;
Accepted: March 11, 2013;
Published: April 16, 2013
How to cite this article
Kartikeya Tiwari, 2013. Frequency Distribution and Assessment of Genetic Diversity of Novel Endophyte Alternaria alternata Accessions Isolated from Pongamia pinnata L. Pakistan Journal of Biological Sciences, 16: 1004-1009.
DOI: 10.3923/pjbs.2013.1004.1009
URL: https://scialert.net/abstract/?doi=pjbs.2013.1004.1009
DOI: 10.3923/pjbs.2013.1004.1009
URL: https://scialert.net/abstract/?doi=pjbs.2013.1004.1009
INTRODUCTION
The world is facing the scarcity of fuel and it is one of the major issues for the upcoming future. People are looking for the alternative as a biofuel. Pongamia pinnata, Pierre (family Leguminosae, sub-family Febaceae) is one of the commercially important tree of India, popularly known as ‘Karanja’ explored all over the world for biodiesel production. Recently, this species has been recognized for its high commercial value, where Karanja seeds are used for oil extraction in harnessing the biofuel potential of this plant. Endophytes with the potential of biodiesel production/bioactive metabolite production can be one of the major alternatives for this problem (Strobel, 2003; Tan and Zou, 2001; Tejesvi et al., 2007; Zou et al., 2000; Frohlich et al., 2000).
Rajasthan is one of twenty five hot spots of global biodiversity with approximately 5,000 species of flowering plants. The xerophytic conditions of the Sanganer region of Rajasthan accommodated a large number of fungal endophytes has recently surged, which has led to a considerable amount of research regarding the role of these fungi in host plants (Banerjee, 2011; Murali et al., 2006). Due to extra ordinary role of these fungi in ecosystem, it is necessary to explore them on the basis of morphological, ecological and genetic level (Anthony and Walkes, 1962; Arnold et al., 2003). Characterization of genetic diversity is a prerequisite for efficient conservation and utilization of genetic resources. Conventionally, tree species have been characterized based on morphological characters many of which are known to be influenced by environmental conditions. During the last two decades, several molecular marker techniques such as randomly amplified polymorphic DNA (Rapd), Inter Simple Sequence Repeats (ISSR) have been developed and have found wide application in diversity studies of various genetic resources due to their greater stability, efficiency and accuracy in comparison to morphology (Guo et al., 2001; 2003, Hawksworth, 1991). RAPD markers have caught the fancy of many individuals in the field of applied fungal taxonomy. RAPD markers due to their low cost and easy to handle have been used for different purposes, such as assessment of genetic variability, genetic relationship, identification of mutants etc. in a number of basidiomycetes, ascomycetes and deuteromycetes fungi (Guo et al., 2001, 2003; Hawksworth, 1991). This molecular marker is based on the PCR amplification of random locations in the genome of the fungi. The number of amplification products is directly related to the number and orientation of the sequences that are complementary to the primer in the genome. The availability of such a simple, safe and fast RAPD technique has been of great importance for diversity studies in various group of fungi especially for which the DNA sequence information is not available for generating specific primers (Collado et al., 2001).
The present investigation explores the endophytic association, host specificity, diversity and distribution between the novel fungal isolates and Pongamia pinnata plant samples.
MATERIALS AND METHODS
Collection of plant samples: Plants samples (Pongamia pinnata) from unique environmental niches of Sanganer region, especially those with an unusual biology and possessing novel strategies for survival were selected for study (Ananda and Sridhar, 2002). The samples were collected in sterilized polythene bags from the different locations of Sanganer region of Jaipur, during the months of July-November.
Isolation of fungal endophytes: The leaves and nodes were used as explants for isolation of fungal endophytes (Frohlich et al., 2000; Tiwari, 2011; Raviraja et al., 2006). All explants were surface-sterilized by dipping in 75% ethanol for 1 min, 4% sodium hypochlorite for 5 min followed by rinsing three times in sterilized distill water. In each Petri dish (9 cm diameter), a total of four-five processed explants were evenly spaced onto the surface of Potato Dextrose Agar (PDA) media supplemented with 200 μg mL-1 tetracycline incubated at 28°C and daily observation was recorded. The sporulating mycelia of fungi appeared on the plates were carefully isolated, sub-cultured and maintained in the pure culture (Murali et al., 2006).
Identification of fungal endophytes: The isolated endophytic fungi have been described and identified on the basis of morphological features like colony characterization, growth of fungi on different media, colour of colony (front and reserve), conidial development, size, shape, conidia and attachment of conidia (Nagamani et al., 2006). Then the fungus is grown in a slide culture by which the pores of the fungus remain undisturbed and attached to the sporophores thus facilitating in identification (Anthony and Walkes, 1962). This technique was performed for various stages of conidia formation and proper identification of the sporulating fungi. The microscopic identification of fungal endophytes was carried out by lacto phenol cotton blue staining method (Nagamani et al., 2006).
Data analysis: Data analysis was done by calculating the Colonization rate (%) of fungal endophytes which was equal to the number of segments colonized by a single endophyte divided by the total number of segments observedx100 (Raviraja, 2005; Raviraja et al., 2006).
Isolation of DNA: The fungal DNA was isolated as per the protocol (Guo et al., 2003, 2004):
| • | A 5 mm disc separated from 7 days old culture growing actively on potato dextrose agar medium was transferred in to 150 mL Erlenmeyer flask containing 30 mL of potato dextrose broth and then incubated at 28°C for 7 days. Mycelial biomass was harvested by centrifugation at 5000 rpm for 5 min then washed two times with deionized distilled water |
| • | A 2 g of biomass was grinded in lysis buffer with help of precooled mortar and pestle |
| • | Vortex the above for 1 min and spun at 7000 rpm for 10 min. Supernatant was decanted in a fresh centrifuge tube and 2 mL of compound solution was added and gently shaken for 15 sec. Five hundred microlitter of upper aqueous layer was pipette out in 1.5 mL of eppendorf tube |
| • | Nucleic acid precipitated from these aqueous phase by addition of 50 μL of 4 M ammonium acetate and 950 μL of precooled ethanol, mixed gently by inversion and incubated in deep freezer for an hour |
| • | The above eppendorf tube was spun at 10, 000 rpm for 5 min supernatant was discarded, DNA pellet was dried by inverting the tube on paper towel for 15 min and redissolved in 40 μL of TE buffer and stored at 4°C |
RNase treatment: The isolated DNA sample was raised up to 400 μL with TE buffer and to it 2 μL of RNAase A (10 mg mL-1 solution of RNAase) was added, mixed by inversion and incubated at 37°C for 30 min DNA was precipitated by addition of 50 μL of 4 M ammonium acetate and 950 μL of precooled ethanol. The content was mixed gently by inversion and spun at 10, 000 rpm for 5 min. The supernatant was discarded. The pellet was dried by inverting the tube on the paper towel for 15 min and DNA pellet was redissolved in 40 μL of TE buffer and stored at 4°C (Guo et al., 2003, 2004).
RAPD Primers: A 500 μL of TE buffer was added in spinned primer tube, vortexed and centrifuged.
PCR assays: Polymerase chain reaction (PCR) was carried out in 25 μL reaction mixture containing 12.5 μL of 2 X PCR buffer, 2 μL each of dATP, dGTP, dTTP and dCTP, Taq DNA polymerase decamer random primer and 50 ng of generic DNA. Amplification reaction performed in Thermal Cycler (Mycycler Bio Rad). The sequential steps were: 1 cycle 3 minutes at 94°C, 2 min at 40°C and 3 min at 72°C followed by 38 cycle of 1 minutes 94°C, 2 min 40°C and 2 min 72°C. The last 10 min extension at 72°C was carried out (Guo et al., 2004; Williams et al., 1990).
Gel casting: Agarose gels were used for separation of PCR products. The comb was adjusted in such a way that it was part few mm from the bottom of the boat. A 1 g of agarose was weighed and transferred to Erlenmeyer flask then added 1xTBE buffer, heated till it boils and cooled under running water to reach up to 50°C. A 5 μL 100 mL-1 ethidium bromide was added from stock of 10 mg mL-1 and poured gently into the boat. After solidification TBE buffer was poured on the gel and comb was removed gently (Guo et al., 2004; Williams et al., 1990).
NTSYS: The scores (0 or 1) for each band were entered in the form of a rectangular data matrix (qualitative data matrix). Cluster analysis for the genetic distance was then carried out using UPGMA (Unweighted Pair Group Method Analysis) clustering method. The genetic distances obtained from cluster analysis through UPGMA were used to construct the dendrogram, depicting the relationships of the clones using computer program NTSYS pc version 2.02 (Rohlf, 1997).
RESULTS
Statistical analysis: A total of eight different endophytic fungal genera along with five different mycelia sterilia were found in leaves and stems of Pongamia pinnata. Among these, Alternaria alternata showed the highest colonizing frequency (22.29%), followed by Fusarium (8.7%) and Colletotrichum species (7.85%). In comparison, Heminthosporium sp. (5.09%), Curvularia sp. (5.3%) and Aspergillus sp. (3.6%), were isolated with low frequency of colonization. Phomopsis sp. (1.27%) and Cladosporium sp. (0.42%) were isolated with the very low frequency of colonization. Fusarium sp., Colletotrichum sp. was found absent in site VI and VII and IX, where as Cladosporium sp. was found absent in site II, IV, V, VI, VIII, XI and X, respectively. The nodal explants shown the highest colonization rate (91.5%) followed by the leafy explants (76.5%) and internodal explants (67.5%).
Genetic diversity: Random amplified polymorphic DNA (RAPD) techniques can be employed for detecting genetic diversity of endophytic fungi from different plants and for pre-selection of these isolates for bioactive screening program (Guo et al., 2001, 2003 and 2004). These methods not only explain the diversity of particular gene bands but also discriminate the genus, species or strains at molecular level for taxonomic description, systematic studies (Collado et al., 2001), hybrid identification and genetic mapping for wide variety of organisms (Pandey et al., 2003; Sim et al., 2010). In this technique polymerase chain reaction is performed with arbitrary 10-mer primers for molecular characterization of morphologically similar and ecologically variant isolates of fungi (Singh et al., 2009). In this analysis the 10 base primers were used during the characterization at genetic level (Operon Technologies. Inc. USA). These primers were selected from preliminary screening and on the basis of their ability to generate highly reproducible RAPD fingerprints that could be used to differentiate morphological similar fungi. Out of these five primers, GCC-181 has given maximum of 116 bands in Alternaria alternata (Fig. 1).
Total genomic DNA was isolated from different fungal isolates and treated with RNase to eliminate RNA. DNA concentration was measured by UV-absorbance method. The integrity of the isolated DNA was verified by visualization of DNA on Agarose gel (0.8%) with DNA standard uncut lambda DNA. The quality of DNA was determined as the ratio A260/A280, which ranged from 1.8 to 1.9, which is indicative of good quality microbial DNA. All the ten fungi were examined for random amplified polymorphic DNA (RAPD) markers with 10 decamer primers purchased from UBC primers procured from GCC [viz., GCC-180 (GGGCCACGCT), GCC-181 (ATGACGACGG)]. Out of 5 primers used for screening, 3 did not amplify any fragment. The reproducibility of the bands generated by these 2 primers was confirmed by replicating the amplification thrice. The total number of bands generated by 2 amplifying primers was 23 with an average amplification of 11.5 bands per primer. The average polymorphosim generated by these bands was 48.75%. The size of the amplification products ranging between 264 to 21226 bp (Fig. 1).
Cluster analysis: The banding pattern generated and polymorphism reflected in these patterns was used to calculate the diversity among the isolates (Fig. 1). A total of 6 polymorphic bands (loci) were produced by primer GCC 180 and GCC 181. Maximum numbers of loci 6 were produced by A114 (when amplified by GCC 180). All the studied strains produced minimum number of bands i.e. 2 when amplified by GCC 180 (Fig. 1). Locus 1, 2, 3 and 4 (896, 968, 1368 and 1532 bp) (produced by GCC 180). The monomorphic bands indicating the molecular diversity of these particular genes in all the isolates. The monomorphic band (produced from GCC) express the diversity of particular loci in all the isolates. The similarity coefficient matrix generated for the primers was subjected to algorithm UPGMA (Unweighted Pair Group Method Analysis) and clusters were generated using NTSYS 2.02 pc program (16).
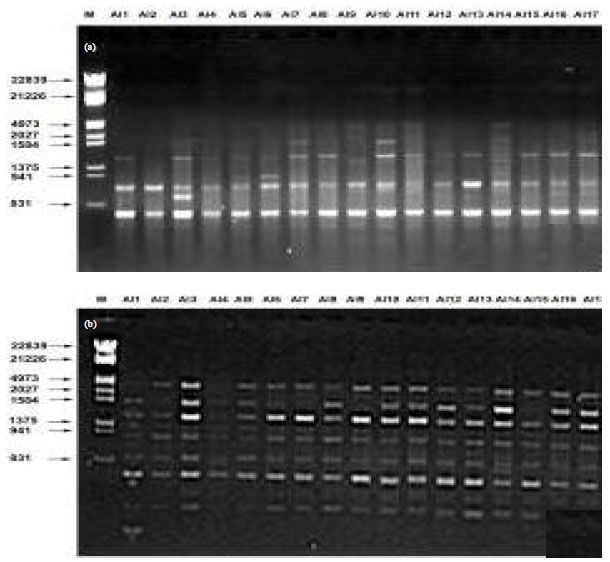 | |
| Fig. 1(a-b): | Representative profiles of Alternaria alternata (Al 1-17) accessions using RAPD primer combination (a) GCC-180 and (b) GCC-181, M is the 500 bp ladder used as size markers |
To stabilize the level of relatedness among the 17 ecologically variant isolates of Alternaria alternata, the dendrogram was constructed (Fig. 2), which showed that the total isolates categorized into three group and first group has twelve isolates, second group has four isolates and third group has only one isolate. First group is again divided into five subgroups, subgroup 1 has five isolates (Al 1, Al 8, Al 12 and Al 13), subgroup 2 has two isolates (Al 2 and Al4), subgroup 3 has four isolates (Al 3, Al 15, Al 16 and Al 17), subgroup 4 and 5 has one isolates (Al 10 and Al 6), respectively. The second group has also two subgroups, which consist of one and three isolates respectively. Two isolates Al2 and Al 4 of subgroup two (Group first) and two isolates Al 9 and Al 11 of subgroup two (Group second) have similarity value >0.9 showed that these strains /isolates are genetically near to each other (Fig. 2).
DISCUSSION
In the present investigation, genetic diversity among seventeen Alternaria alternata accessions were employed using two RAPD primers (GCC-180 and GCC-181). The high level of genetic diversity observed in this study within the limited number of accessions which were endophytically colonized and ecologically variant. Accuracy and robustness in results of genetic analysis depends mainly on the accuracy of the input data (Williams et al., 1990, Pandey et al., 2003, Tiwari, 2011).
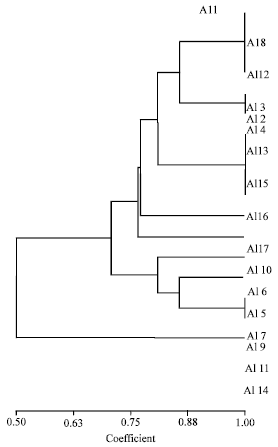 | |
| Fig. 2: | Dendrogram showing UPGMA clustering of accessions using Random Amplified Polymorphic DNA (RAPD) analysis of Alternaria alternata 1 to 17 |
The number of bands obtained in RAPD assays depends primarily on the number of selective nucleotides and their sequence. With the same primer combination, two different species with different genome complexities may produce significantly different number of bands. Thus, the total number of bands in RAPD can be modulated for a particular species to produce easily scorable profiles (Rohlf, 1997).
These results are in agreement with the results obtained by Guo et al. (2004) who reported genetic diversity and endophytic association between Alternaria alternata and Pinus tabulaeformis plant samples using RAPD analysis. Similar results were also reported by Frohlich et al. (2000) who reported endophytic relationship between fungal isolates and palm trees.
CONCLUSION
As a concluding remark, we can state that seventeen Alternaria alternata accessions were ecologically diversified with host specificity in the Pongamia pinnata plant samples, endophytically.
ACKNOWLEDGMENT
The author thanks Department of Botany, University of Rajasthan, Jaipur, India for providing laboratory facilities.
REFERENCES
- Ananda, K. and K.R. Sridhar, 2002. Diversity of endophytic fungi in the roots of mangrove species on the West Coast of India. Can. J. Microbiol., 48: 871-878.
CrossRefPubMedDirect Link - Anthony, E.H. and A.C. Walkes, 1962. An improvement in slide culture technique. Can. J. Microbiol., 8: 929-930.
CrossRefDirect Link - Arnold, A.E., L.C. Mejia, D. Kyllo, E.I. Rojas, Z. Maynard, N. Robbins and E.A. Herre, 2003. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA., 100: 15649-15654.
CrossRefDirect Link - Banerjee, D., 2011. Endophytic fungal diversity in tropical and subtropical plants. Res. J. Microbiol., 6: 54-62.
CrossRefDirect Link - Collado, J., G. Platas and F. Pelaez, 2001. Identification of an endophytic Nodulisporium sp. from Quercus ilex in central Spain as the anamorph of Biscogniauxia mediterranea by rDNA sequence analysis and effect of different ecological factors on distribution of the fungus. Mycologia, 93: 875-886.
Direct Link - Frohlich, J., K.D. Hyde and O. Petrini, 2000. Endophytic fungi associated with palms. Mycol. Res., 104: 1202-1212.
Direct Link - Guo, L.D., K.D. Hyde and E.C.Y. Liew, 2001. Detection and taxonomic placement of endophytic fungi within frond tissues of Livistona chinensis based on rDNA sequences. Mol. Phylogenet. Evol., 20: 1-13.
CrossRefPubMedDirect Link - Guo, L.D., G.R. Huang, Y. Wang, W.H. He, W.H. Zheng and K.D. Hyde, 2003. Molecular identification of white morphotype strains of endophytic fungi from Pinus tabulaeformis. Mycol. Res., 107: 680-688.
CrossRefPubMedDirect Link - Guo, L.D., L. Xu, W.H. Zheng and K.D. Hyde, 2004. Genetic variation of Alternaria alternate, an endophytic fungus isolated from Pinus tabulaeformis as determined by random amplified microsatellites (RAMS). Fungal Diversity, 16: 53-65.
Direct Link - Hawksworth, D.L., 1991. The fungal dimesion of biodiversity magnitude, significance and conservation. Mycol. Res., 95: 641-655.
CrossRef - Murali, T.S., T.S. Suryanarayanan and R. Geeta, 2006. Endophytic Phomopsis species host range and implications for diversity estimates. Canadian J. Microbiol., 52: 673-680.
PubMed - Raviraja, N.S., G.L. Maria and K.R. Sridhar, 2006. Antimicrobial evaluation of endophytic fungi inhabiting medicinal plants of the Western Ghats of India. Eng. Life Sci., 6: 515-520.
CrossRef - Raviraja, N.S., 2005. Fungal endophytes in five medicinal plant species from Kudremukh Range, Western Ghats of India. J. Basic. Microbiol., 45: 230-235.
CrossRef - Strobel, G.A., 2003. Endophytes as sources of bioactive products. Microbes Infect., 5: 535-544.
CrossRefPubMedDirect Link - Sim, J.H., C.H. Khoo, L.H. Lee and Y.K. Cheah, 2010. Molecular diversity of fungal endophytes isolated from garcinia mangostana and garcinia parvifolia. J. Microbiol. Biotechnol., 20: 651-658.
PubMed - Tan, R.X. and W.X. Zou, 2001. Endophytes: A rich source of functional metabolites. Nat. Prod. Rep., 18: 448-459.
PubMedDirect Link - Tejesvi, M.V., K.R. Kini, H.S. Prakash, V. Subbiah and H.S. Shetty, 2007. Genetic diversity and antifungal activity of species of Pestalotiopsis isolated as endophytes from medicinal plants. Fungal Diversity, 24: 37-54.
Direct Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link - Zou, W.X., J.C. Meng, H. Lu, G.X. Chen, G.X. Shi, T.Y. Zhang and R.X. Tan, 2000. Metabolites of Colletotrichum gloeosporidies and endophytic fungus in Artemisia mongolica. J. Natural Prod., 63: 15229-15230.
PubMed