
S.A.M.C. Samarakoon
Faculty of Agriculture, Rajarata University of Sri Lanka, Puliyankulama, Anuradhapura, Sri Lanka
A. Balasuriya
Faculty of Agriculture, Rajarata University of Sri Lanka, Puliyankulama, Anuradhapura, Sri Lanka
R.G.A.S. Rajapaksha
Horticultural Crops Research and Development Institute, Gannoruwa, Peradeniya, Sri Lanka
W.A.R.T. Wickramarachchi
Horticultural Crops Research and Development Institute, Gannoruwa, Peradeniya, Sri Lanka
Pakistan Journal of Biological Sciences
Year: 2012 | Volume: 15 | Issue: 18 | Page No.: 863-870







ABSTRACT
Tomato Yellow Leaf Curl Virus (TYLCV) is an important plant virus on one of the economically most important vegetable crops; tomato (Lycopersicon esculentum Mill.). This had not been molecularly detected before, in Sri Lanka. TYLCV-GN-SL was isolated from apparently infected tomato plants using modified Cetyltrimethyl Ammonium Bromide (CTAB) method in Gannoruwa. Associated Begomoviruses were detected using Deng 541/Deng 540 and AV 494/AC 1048 primer pairs. TYLCV was detected for the first time in tomato in Sri Lanka using P1V/P4C, TYLCV specific primer pair. Nucleotide sequence of coat protein of isolated TYLCV-GN-SL proved that the Indian strain of ToLC virus was closely related to Tomato Leaf Curl Sri Lanka Virus (TLCV-SL: 97%) and Tomato leaf curl Geminivirus (TLCGV: 93%) through direct sequencing data. TLCV-SL was confirmed as TYLCV isolate. TYLCV was molecularly detected from major tomato growing districts like Badulla, Nuwara-Eliya, Kandy and Matale in Sri Lanka.
PDF Abstract XML References Citation
Received: October 04, 2012;
Accepted: December 10, 2012;
Published: February 12, 2013
How to cite this article
S.A.M.C. Samarakoon, A. Balasuriya, R.G.A.S. Rajapaksha and W.A.R.T. Wickramarachchi, 2012. Molecular Detection and Partial Characterization of Tomato Yellow Leaf Curl
Virus in Sri Lanka. Pakistan Journal of Biological Sciences, 15: 863-870.
DOI: 10.3923/pjbs.2012.863.870
URL: https://scialert.net/abstract/?doi=pjbs.2012.863.870
DOI: 10.3923/pjbs.2012.863.870
URL: https://scialert.net/abstract/?doi=pjbs.2012.863.870
INTRODUCTION
Tomato Yellow Leaf Curl Disease (TYLCD) caused by several viruses belonging to different species, which all together are referred to as Tomato Yellow Leaf Curl Virus (TYLCV) of the genus Begomovirus of the family Geminiviridae (Czosnek, 2008) is one of the most devastating plant disease in many tropical and subtropical regions in the world (Reddy, 2006). The virus is ranked third among ‘Top 10 plant viruses’ (Scholthof et al., 2011). In Sri Lanka, a high incidence of TYLCD was reported during April to September (Ariyarathna et al., 2004).
TYLCV is primarily transmitted by the sweet potato whitefly (Bemisia tabaci Gannadius) in a persistent and circulative manner (Melzer et al., 2009) but is not transmitted by the greenhouse whitefly (Trialeurodes vaporariorum) (Sugano et al., 2011). It is not mechanically or seed transmitted (Green and Kalloo, 1994; Cerkauskas, 2005; Czosnek, 2008).
TYLCV genome is either monopartite or bipartite and has geminate (twinned) particles size of 18-20x30 nm, consisting of two incomplete icosahedra joined together in a structure with 22 pentameric capsomeres and 110 identical protein subunits (Gafni, 2003; Czosnek, 2008). A functional coat protein with 260 amino acid length is essential for host plant infection and insect transmission in TYLCV (Noris et al., 1998). TYLCV was the first Begomovirus proven to possess a single genomic DNA (ssDNA) and it has a 2787 nt (Czosnek, 2008). It encodes six partially overlapping Open Reading Frames (ORFs) that are organized bi-directionally (Gafni, 2003) and a long (313 nt) Intergenic Region (Czosnek, 2008).
Symptom development in tomato is in 2- 3 weeks after inoculation of TYLCV but the viral DNA can be detected 7 days earlier (Ber et al., 1990) and the highest viral concentration can be detected at 4 days before symptom appearance (Green and Kalloo, 1994). Typical symptoms in tomato leaves are yellow (chlorotic) leaf edges, upward leaf cupping, leaf mottling with dark green and yellow, leathery, reduced leaf size and abnormally shaped foliage with evident vein clearing, short internodes, erect terminal and axillary shoots, proliferation of axillary branch formation, premature flower drop and loss of small fruits (Brown and Nelson, 1988; Costa and Heuvelink, 2005; Reddy, 2006).
Symptomatological identification of TYLCV is unreliable (Momol et al., 1999) and the Polymerase Chain Reaction (PCR) methods are highly effective as a tool for rapid and large-scale diagnostics of TYLCV-infected samples (Briddon and Markham, 1995). Apart from being a tool useful for detection, the products of the diagnostic PCR reactions are suitable for further characterization of the viruses (Briddon and Markham, 1995).
Virus taxonomy must respond to the development of new technologies (Fauquet and Stanley, 2003). The analysis of DNA sequences has become the tool of choice, allowing to accurately identify the virus and to evaluate its relationship with other TYLCV isolates (Czosnek, 2008). The phylogenetic analysis is important in showing ancestral relationships among geminviruses and is critical for virus taxonomy. Often, complete genomes or gene sequences are used. However, where complete sequences are not available, partial nucleotide sequences are useful in determining relationships among new and emerging geminiviruses (Roye et al., 1999).
ToLCV affected tomato samples were tested with graft inoculation in Sri Lanka. Infected samples were detected by DNA hybridization using 6i probes, which detected the Indian monopartite tomato geminivirus by Dr. S.K. Green in Asian Vegetable Research and Development Centre (De Zoysa, 1996). Tomato leaf curl Sri Lanka virus-(Sri Lanka: Bandarawela: 1997)-ToLCSLV-(LK: Ban: 97) species has been published (Fauquet et al., 2008).
This study was the first molecular detection and partial characterization of the TYLCV in Sri Lanka.
MATERIALS AND METHODS
The experiments were conducted at the Division of Plant Pathology, Horticultural Crops Research and Development Institute (HORDI), located in the Latitude of 7°16'N and the Longitude of 80°36'E from April to September 2012.
Sample collection and total DNA extraction: Samples from the tomato plants showing typical characteristic TYLCD symptoms and healthy tomato plants grown under insect proof conditions were collected with the avoidance of cross contaminations.
Total DNA was extracted from apparently infected and healthy samples using CTAB extraction protocol previously described by Lodhi et al. (1994) with slight modifications. Approximately 150 mg of tender leaf tissues were homogenized using a mortar and pestle with 1.5 mL of pre-warmed extraction buffer (2% (v/v) CTAB, 100 mM Tris-HCl (pH 8.0), 20 mM EDTA, 1.4 M NaCl and 0.2% (v/v) β-mercaptoethanol added just before use) with 50 mg of PVP. The homogenized sample was transferred into a 1.5 mL micro centrifuge tube and incubated at 65°C for 30 min while mixing at 10 min intervals. The tube was centrifuged at 10,000 rpm for 5 sec and 750 μL of supernatant was transferred into a new tube and treated with 750 μL of chloroform: isoamyl alcohol (24:1) followed by vortexing and centrifuging at 10,000 rpm for 15 min. Aqueous layer was transferred into a new tube and 300 μL of ice-cold isopropanol was added and mixed by inverting the tube slowly. The tube was incubated overnight at -20°C. The DNA was pelleted down by centrifuging at 10,000 rpm for 15 min. The resultant pellet was washed with 500 μL of 70% (v/v) ethanol by vortexing followed by centrifugation at 10,000 rpm for 5 min. The DNA pellet was dried until all the alcohol was evaporated and dissolved in 50 μL of TE (Tris-EDTA) buffer (10 mM Tris-HCl (pH 8) and 1 mM EDTA (pH8)), incubated at 37°C for 30 min and stored at -20°C until further used.
PCR amplification: PCR was carried out with oligonucleotide primers (Table 1).
A PCR was conducted in a reaction volume (25 μL) containing 14.8 μL of sterile water, 2.5 μL of 10X PCR buffer (500 mM KCl, 100 mM Tris-HCl (pH 9.1) and 0.1% Triton™ X-100), 2 μL of dNTP (2.5 mM), 2 μL of each Deng 541/Deng 540 (10 μM), 0.5 μL of MgCl2 (25 mM), 0.2 μL of Taq polymerase (5 IU μL-1) and 1 μL of template DNA (diluted 1:25 in water). Following thermal cycle programme was performed, 1 cycle (4 min at 94°C), 30 cycles (30 sec at 94°C, 30 sec at 58°C and 45 sec at 72°C) and 1 cycle (10 min at 72°C) in a thermocycler (Labnet Inc. U.S.A, Model: Labnet Gradient).
A PCR was conducted in a reaction volume (25 μL) containing 13.4 μL of sterile water, 2.5 μL of 10X PCR buffer (500 mM KCl, 100 mM Tris-HCl (pH 9.1) and 0.1% Triton™ X-100), 2 μL of dNTP (2.5 mM), 2 μL of each AV 494/AC 1048 (10 μM), 1.0 μL of MgCl2 (25 mM), 0.1 μL of Taq polymerase (5 IU μL-1) and 2 μL of template DNA (diluted 1:25 in water). Following thermal cycle programme was performed, 1 cycle (5 min at 94°C), 30 cycles (30 sec at 94°C, 30 sec at 50°C and 45 sec at 72°C) and 1 cycle (10 min at 72°C).
| Table 1: | Oligonucleotide primers used to detect TYLCV Begomovirus associated with tomato |
 | |
| K: G or T, R: A or G, S: C or G, W: A or T, Y: C or T, B: C, G or T, V: A, C or G | |
PCR was conducted in a reaction volume (25 μL) containing 14.3 μL of sterile water, 2.5 μL of 10X PCR buffer (500 mM KCl, 100 mM Tris-HCl (pH 9.1) and 0.1% Triton™ X-100), 2 μL of dNTP (2.5 mM), 2 μL of P1V/ P4C (10 μM), 1.0 μL of MgCl2 (25 mM), 0.2 μL of Taq polymerase (5 IU μL-1) and 1 μL of template DNA (diluted 1:25 in water). Following thermal cycle programme was performed, 1 cycle (4 min at 94°C), 30 cycles (1 min at 94°C, 1 min at 55°C and 2 min at 72°C) and 1 cycle (10 min at 72°C). PCR products were stored at-20°C.
PCR products (5 μL of each) were subjected to 1% (w/v) agarose gel electrophoresis with 2 μL of loading dye at 80 volts for 1 hour in TBE buffer and stained with ethidium bromide (0.5 μg mL-1) and visualized under UV transilluminator and photographed by a digital camera. The results were verified against DNA marker (Vivantis).
Molecular characterization: Twenty micro liters of PCR product of ~520 bp consistently amplified using Deng 541/Deng 540 from TYLCV confirmed sample was sent to the Gene Tech Pvt. Ltd, 54, Kitulwatte Road, Colombo 08, Sri Lanka for sequencing. Direct sequenced data of CP was subjected to FASTA analysis based on close sequence identity and the length of the sequences. Followed by, Begomovirus of highly similar sequences were downloaded from GenBank with the accession numbers provided by the FASTA output. Phylogenetic tree and molecular evolutionary relationships were determined through MEGA 4.0 software using neighbor-joining method (Tamura et al., 2007).
Detection of the presence of TYLCV in major tomato growing districts in Sri Lanka: Naturally infected tomato plants showing typical characteristic symptoms of TYLCD along with healthy tomato plants were collected from Badulla, Nuwara-Eliya, Kandy and Matale Districts. Characteristic features of the infected plants and varieties were recorded and photographed. Tender leaf samples were used for DNA extraction using modified CTAB method. Extracted DNA samples were amplified with degenerate universal primers (Deng et al., 1994) for Begomovirus and TYLCV specific primers (Navot et al., 1992). All the conditions as above relevant with primers. PCR products were subjected to gel electrophoresis with DNA marker (Vivantis).
RESULTS AND DISCUSSION
Molecular detection of TYLCV: Apparently infected tomato plants with yellow colour margins, dark green mid veinal area, shrinking and curling of the leaflets and the stunting of the plant (Fig. 2) were subjected to DNA extraction.
Diluted DNA sample (1:25) amplified with the degenerate primers (Deng et al., 1994) yielded the 520 bp size of band (Fig. 1a) suggesting the presence of a Begomovirus in suspected sample. For the identification and confirmation of all the TYLC viruses and ToMoV, it had been recommended the PCR with Deng’s degenerate primers (Anonymous, 2004). PCR amplification resulting Begomovirus specific band showed the presence of Begomovirus in sample showing typical TYLCD.
 | |
| Fig. 1(a-b): | Detection of Begomovirus presence of the suspected tomato sample (a) Detection of Begomovirus in infected tomato sample using degenerate primers (Deng et al., 1994), Lanes-M: 100 bp marker (Vivantis), 1: Infected tomato sample, 2: Healthy tomato sample, 3: Water control and (b) Detection of Begomovirus using Wyatt and Brown’s and Deng’s degenerate primers, Lanes-M: 100 bp marker (Vivantis), 1: Infected bean sample, 2: Infected tomato sample (Wyatt and Brown’s degenerate primers), 3: Infected cassava sample (Deng’s degenerate primers), 4: Water control |
 | |
| Fig. 2: | TYLCV infected tomato plant |
The same set of degenerate primers had been used for the detection of Begomoviruses in infected tomato-Banglore and naturally infected weed hosts (Reddy, 2006).
PCR primers that anneal to two highly conserved sequences within the most highly conserved gene of the whitefly transmitted geminivirus subgroup would be useful for broad-spectrum PCR based virus detection. Highly conserved DNA motifs within the capsid gene of subgroup III geminiviruses are potentially suitable as degenerate priming sites (Wyatt and Brown, 1996). The AV 494/AC 1048 primer pair is ideal because of the anticipated conservation and hence their apparently universal nature. DNA sequence of the 550 bp core capsid gene fragment was amplified by another set of degenerate primers (Wyatt and Brown, 1996). The selected sample that is shown typical TYLCD symptoms, amplified with the above degenerate primers, resulting 550 bp PCR products (Fig. 1b). In Turkey, 15 symptomatic tomato samples were tested for Begomovirus infection by using PCR with Wyatt and Brown’s degenerate Begomovirus primers (Koklu et al., 2006). Begomovirus infected bean samples were amplified with Wyatt and Brown’s degenerate primers resulting 550 bp of bands. Infected cassava sample amplified with Deng’s degenerate primers showed the 520 bp band as a reference. The results showed further, the association of Begomovirus in infected tomato plant sample showing typical TYLCD.
DNA sample, which was positive for the presence of Begomovirus using Deng’s and Wyatt and Brown’s degenerate primers, amplified with the TYLCV specific primer (P1V and P4C) resulting 1650 bp PCR product (Fig. 3). P1V and P4C primers anneal with the 6-80 and 2054-2071 positions of the viral genome, respectively.
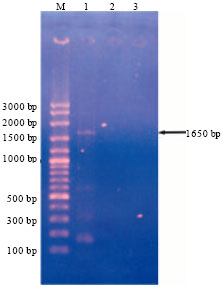 | |
| Fig. 3: | Detection of TYLCV in infected tomato sample using TYLCV specific primers, Lanes-M: 100 bp marker (Vivantis), 1: Infected tomato sample, 2: Healthy tomato sample, 3: Water control |
Navot et al. (1992) reported similar results using total DNA extracted from the TYLCV infected tomato samples in Israel. TYLCV specific primers P1V (corresponds to the viron positive strand) and P4C (corresponding to complementary viron strand) were used for the detection of TYLCV and Immuno-captured (IC) PCR was employed in Palestine (Sawalha, 2012).
Based on the PCR with TYLCV specific primer, it clearly showed the presence of TYLCV in the tomato sample, with typical TYLCD symptoms. However, the healthy sample was not found positive with both Begomovirus specific primes and TYLCV specific primers. This is the first attempt made for molecular detecting of TYLCV in Sri Lanka.
The nucleic acid hybridization with Sri Lankan probe SL 14 confirmed the presence of TYLCV in samples of T-245 and Thilina varieties sent to Taiwan in 2000 (Ariyarathna et al., 2004). Tomato plant samples showing leaf mottling, curling, yellowing, purpling, discoloration, distortion and stunting, spotted on nitrocellulose membranes were sent to Asian Vegetable Research and Development Centre (AVRDC) for DNA hybridization tests using Indian tomato geminivirus probe and found negative results (De Zoysa, 1996).
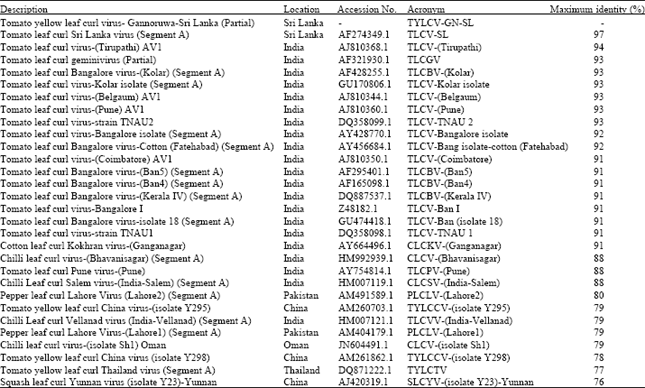
Molecular characterization of TYLCV in tomato: FASTA format of the sequence, denoted as TYLCV-GN-SL (Tomato yellow leaf curl virus-Gannoruwa-Sri Lanka) was used for analysis. Other background nucleotide fragments were removed from the direct sequence data that compared with several other TYLCV sequences.
| Table 2: | FASTA format of sequenced data compared with 29 isolates of Begomoviruses |
 | |
| Presence of TYLCV in major tomato growing districts in Sri Lanka | |
It has resulted 491 nt fragment as a partial sequence of TYLCV-GN-SL. This type of contaminations is common in direct sequencing process. However, sequencing after cloning with a suitable vector could be much more reliable. FASTA format of sequenced data was compared with 29 isolates of other Begomoviruses (Table 2) of highly similar sequences and a phylogenetic tree was made using MEGA 4.0 software. The CP region of the virus displayed different similarity percentages with other related viruses (Fig. 4).
TYLCV-GN-SL isolate was grouped together with Tomato Leaf Curl Sri Lanka Virus (TLCV-SL: AF274349.1) of 97%, tomato leaf curl geminivirus (TLCGV: AF321930.1) of 93% as a group in a small cluster, they were found to be very much genetically similar. Further, many of the related Begomoviruses denoted as ToLCV from India have been form a separate cluster but related to a small cluster, where TYLCV-GN-SL isolate included.
TYLCV-GN-SL isolate has been proved to be TYLCV using specific primers suggested by Navot et al. (1992). Therefore, information generated through this study seem sufficient for confirmation of Tomato leaf curl Sri Lanka virus-(Sri Lanka: Bandarawela: 1997) (ToLCSLV- LK: Ban: 97; AF274349) shown high similarity with TYLCV-GN-SL proved as a TYLCV, the nomenclature of the ToLCSLV may be changed as TYLCV.
Our isolate being genetically similar with most of the geminiviruses from tomatoes in India, it can be suggested that Sri Lanka TYLCV is a strain of ToLCV in Southern India. As a whole, the Indian ToLCV isolates are different from the tomato geminiviruses found in other regions of East and Southeast Asia and Australia (Muniyappa et al., 2000). Point mutation in one or several nucleotides can lead to the genetic diversity of viruses. Such genetical events in viral genome can result in many isolated strains that are genetically very much similar. Genomic recombination in geminiviruses, not only between the variants of the same virus but also between species and even between genera, has resulted in rapid diversification. Due to the rapidly increasing number of geminiviruses that are being isolated, there has been an urgent need for an improved system of classification and nomenclature (Fauquet and Stanley, 2003; Mehrotra and Aggarwal, 2003; Varma and Malathi, 2003).
The <89% of nucleotide identity threshold between full-length of DNA-A component nucleotide sequences for Begomovirus species as a species demarcation criteria is very important.
 | |
| Fig. 4: | Phylogenetic tree showing the genetic relationship of TYLCV- GN- SL isolated from Sri Lanka to other related and begomoviruses. The numbers appearing at each node indicate the percentage of supporting bootstrap samples (1,000 replicates) |
Proposed 89% threshold is not an absolute criterion and it is possible for viruses to share more than 89% identity yet be classified as distinct species on the strength of their biological properties and vice versa (Fauquet and Stanley, 2003).
Geographical location of Sri Lanka being very close to India can be a risk of trans-boundary-migration of virus infected plants, viruliferous whitefly vectors etc. Genetic similarity of TYLCV-GN-SL with related Indian isolates implies this type of trans boundary migration.
It is important to sequence of the full genome of the TYLCV in Sri Lanka for the obtaining of more information for further classification. In several cases, the CP of TYLCV was more homologous to the CP of other whitefly-transmitted geminiviruses occurring in the same region than to TYLCV isolates from other regions, a fact that may point to adaptation of the geminivirus to its vector (Czosnek and Laterrot, 1997).
Tomato samples collected from Badulla, Nuwara-Eliya, Kandy and Matale districts showed characteristic symptoms of TYLCV identified (Table 3).
| Table 3: | Visual diagnosis and molecular detection of TYLCV from different districts |
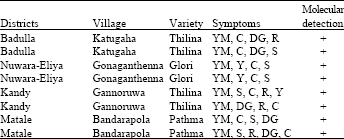 | |
| YM: Yellow color leaf margin area, Y: Yellowing of leaflets, DG: Dark green veinal area, C: Curling, S: Shrinking of leaflets, R: Leaf size and growth reduction, +: Positive results with degenerate primers (Deng et al., 1994) | |
The PCR studies using universal primers showed that all samples were infected with Begomovirus and resulted 520 bp bands (Fig. 5). It suggested that TYLCD is one of the major constraints in major tomato growing districts.
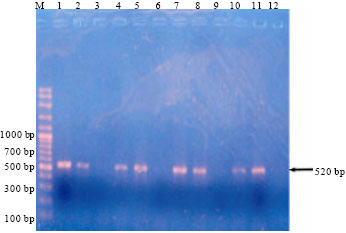 | |
| Fig. 5: | Molecular detection of TYLCV from major tomato growing districts, Lanes-M: 100 bp marker (Vivantis), 1, 2: Infected, 3: Healthy tomato samples (Badulla), 4, 5: Infected, 6: Healthy tomato samples (Nuwara: Eliya), 7, 8: Infected, 9: Healthy tomato samples (Kandy), 10, 11: Infected, 12: Healthy tomato samples (Matale) |
TYLCD is the most important and common virus disease of tomato in Sri Lanka and it is reported in all tomato-growing areas. In 1998, 40% TYLCV infection of variety T- 146 was reported in Katugastota while 90% of TYLCV infection in variety Caribo was reported at Marassana (Ariyarathna et al., 2004). According to the survey conducted, TYLCD incidence was high in Kandy, Matale and Nuwara- Eliya districts. It affected over 25% of the tomato cultivation and disease severity in some areas has gone up to 75% (Anonymous, 2004). Yield losses in tomato due to TYLCV usually ranged from 50-75% and even up to 100% but may be as high as 100% making tomato production unprofitable (Green and Kalloo, 1994). It is important to take the control measures in the beginning of tomato cultivation. In addition, concerning of the epidemiology of TYLCV is important within major tomato growing areas due to the presence of TYLCV. The TYLCV management practices should be concerned with destroying of even single tomato plant infected with TYLCV to control spreading of virus.
ACKNOWLEDGMENTS
We thank all the staff members of the Horticultural Crops Research and Development Institute (HORDI), Gannoruwa, Peradeniya and Department of Plant Sciences, Faculty of Agriculture, Rajarata University of Sri Lanka.
REFERENCES
- Ber, R., N. Navot, D. Zamir, Y. Antignus, S. Cohen and H. Czosnek, 1990. Infection of tomato by the tomato yellow leaf curl virus: Susceptibility to infection, symptom development and accumulation of viral DNA. Arch. Virol., 112: 169-180.
CrossRef - Briddon, R.W. and P.G. Markham, 1995. Use of PCR in the detection and characterization of geminiviruses. EPPO Bull., 25: 315-320.
CrossRef - Brown, J.K and M.R. Nelson, 1988. Transmission, host range and virus-vector relationships of Chino del tomato virus (CdTV), a whitefly-transmitted geminivirus from Sinaloa, Mexico. Plant Dis., 72: 866-869.
CrossRefDirect Link - Czosnek, H. and H. Laterrot, 1997. A worldwide survey of tomato yellow leaf curl viruses. Arch. Virol., 142: 1391-1406.
CrossRefPubMedDirect Link - Deng, D., P.F. McGrath, D.J. Robinson and B.D. Harrison, 1994. Detection and differentiation of whitefly-transmitted geminiviruses in plants and vector insects by the polymerase chain reaction with degenerate primers. Ann. Applied Biol., 125: 327-336.
CrossRefDirect Link - Fauquet, C.M. and J. Stanley, 2003. Geminivirus classification and nomenclature: Progress and problems. Ann. Applied Biol., 142: 165-189.
CrossRefDirect Link - Fauquet, C.M., R.W. Briddon, J.K. Brown, E. Moriones, J. Stanley, M. Zerbini and X. Zhou, 2008. Geminivirus strain demarcation and nomenclature. Arch. Virol., 153: 783-821.
CrossRefPubMedDirect Link - Gafni, Y., 2003. Tomato yellow leaf curl virus, the intracellular dynamics of a plant DNA virus. Mol. Plant Pathol., 4: 9-15.
CrossRef - Koklu, G., A. Rojas and A. Kvarnheden, 2006. Molecular identification and the complete nucleotide sequence of a tomato yellow leaf curl virus isolate from Turkey. J. Plant Pathol., 88: 61-66.
Direct Link - Muniyappa, V., H.M. Venkatesh, H.K. Ramappa, R.S. Kulkarni and M. Zeidan et al., 2000. Tomato leaf curl virus from Bangalore (ToLCV-Ban4): Sequence comparison with Indian ToLCV isolates, detection in plants and insects and vector relationships. Arch. Virol., 145: 1583-1598.
CrossRefDirect Link - Navot, N., M. Zeidan, E. Pichersky, D. Zamir and H. Czosnek, 1992. Use of the polymerase chain reaction to amplify tomato yellow leaf curl virus DNA from infected plants and viruliferous whiteflies. Phytopathology, 82: 1199-1202.
CrossRef - Noris, E., A.M. Vaira, P. Caciagli, V. Masenga, B. Gronenborn and G.P. Accotto, 1998. Amino acids in the capsid protein of tomato yellow leaf curl virus that are crucial for systemic infection, particle formation and insect transmission. J. Virol., 72: 10050-10057.
Direct Link - Roye, M.E., M.E. Wernecke, W.A. McLaughlin, M.K. Nakhla and D.P. Maxwell, 1999. Tomato dwarf leaf curl virus, a new bipartite Geminivirus associated with tomatoes and peppers in Jamaica and mixed infection with tomato yellow leaf curl virus. Plant Pathol., 48: 370-378.
CrossRefDirect Link - Sawalha, H., 2012. Epidemiology of tomato yellow leaf curl virus in the Northern regions of the West Bank, Palestine. Adv. Life Sci. Appl., 1: 6-12.
Direct Link - Scholthof, K.B.G., S. Adkins, H. Czosnek, P. Palukaitis and E. Jacquot et al., 2011. Top 10 plant viruses in molecular plant pathology. Mol. Plant Pathol., 12: 938-954.
CrossRefDirect Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Varma, A. and V.G. Malathi, 2003. Emerging geminivirus problems: A serious threat to crop production. Ann. Applied Biol., 142: 145-164.
CrossRefDirect Link - Wyatt, S.D. and J.K. Brown, 1996. Detection of subgroup III geminivirus isolates in leaf extracts by degenerate primers and polymerase chain reaction. Phytopathology, 86: 1288-1293.
CrossRefDirect Link - Lodhi, M.A., G.N. Ye, N.F. Weeden and B.I. Reisch, 1994. A simple and efficient method DNA extraction from grapevine caltivars and vitis species. Plant Mol. Biol. Rep., 12: 6-13.
Direct Link