
A.N. Tan
Laboratory of Marine Science and Aquaculture, Institute of Bioscience, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
A. Christianus
Laboratory of Marine Science and Aquaculture, Institute of Bioscience, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
S. Shakibazadeh
Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
P. Hajeb
Center of Excellence for Food Safety Research, Faculty of Food Science and Technology, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
Pakistan Journal of Biological Sciences
Year: 2012 | Volume: 15 | Issue: 13 | Page No.: 610-620







ABSTRACT
Local and regional decline of Asian horseshoe crabs has spurred a study on its spawning population at Balok Beach, Kuantan, Pahang, Malaysia. This location was identified as spawning site due to the occurrence of horseshoe crab spawning pairs and nests. Size-frequency, length-weight relationships, sex ratio and epibiont infestation of Tachypleus gigas were studied. Instar stage was estimated based on prosomal width. Condition of the horseshoe crab carapace was reported. Visual search technique of horseshoe crab was conducted during high tide of new and full moons. Prosomal, opisthosomal and telson length and weight of each horseshoe crab were measured. Largest female was recorded with mean prosomal length and width of 154.4 and 246.9 mm, respectively. About 69.8% of the males belonged to size group of 151-200 mm and 53.3% of females were grouped into 201-250 mm. All individuals were of fourteenth to sixteenth instar stages. Sex ratio varied from 0.313 to 2.5 and attributed to commercial harvest and monsoon season. Sand sediment of study site showed 93% of fine sands with grain size ranged from 120 to 250 μm. Acorn and pedunculate barnacle, conical and flat slipper shells were found on the carapace of the specimens. Most males had damaged eyes and carapaces while females with broken telsons. Body damages of about 19.9% on the specimens were likely due to nearby fishing activities. Lack of satellite male indicated low spawning population. The finding of this study showed that the species is extremely threatened by human activities and coastal development.
PDF Abstract XML References Citation
Received: September 19, 2012;
Accepted: October 19, 2012;
Published: January 23, 2013
How to cite this article
A.N. Tan, A. Christianus, S. Shakibazadeh and P. Hajeb, 2012. Horseshoe Crab, Tachypleus gigas (Müller, 1785) Spawning Population at Balok Beach, Kuantan, Pahang, Malaysia. Pakistan Journal of Biological Sciences, 15: 610-620.
DOI: 10.3923/pjbs.2012.610.620
URL: https://scialert.net/abstract/?doi=pjbs.2012.610.620
DOI: 10.3923/pjbs.2012.610.620
URL: https://scialert.net/abstract/?doi=pjbs.2012.610.620
INTRODUCTION
Three horseshoe crab species namely Tachypleus gigas, Tachypleus tridentatus and Carcinoscorpius rotundicauda are reported in the coastal waters of Malaysia (Kassim et al., 2008; Christianus and Saad, 2009; Zaleha et al., 2010; Srijaya et al., 2010a). The horseshoe crab migrates from offshore continental shelf areas to spawn on estuarine beaches (Brockmann and Smith, 2009) during high tides of new and full moons throughout the year (Sekiguchi et al., 1977; Chatterji et al., 1992; Chatterji, 1994; Hajeb et al., 2005, 2009). Earlier studies showed that T. gigas can be found mostly on sandy beaches with slight influence of mud (Christianus et al., 2005; Zaleha et al., 2011).Most researchers suggest that the Asian horseshoe crabs are declining both locally and regionally. Therefore it is important to consider some mitigation measures to avoid the extinction of this fossil species. Anthropogenic factors are mainly responsible for declining of horseshoe crab population in the United States and Asia (Botton and Haskin, 1984; Shuster, 1982; Itow, 1993; Chatterji, 1994; Michels, 1996; Chiu and Morton, 1999, 2003a; Widener and Barlow, 1999; Botton, 2000; Chen et al., 2002, 2004; Tanacredi, 2002; Burger et al., 2003; Carmichael et al., 2003; Zhou and Morton, 2004; Seino et al., 2004; Almendral and Schoppe, 2005; Lee and Morton, 2005, 2009; Christianus, 2006; Berkson et al., 2009; Cartwright-Taylor et al., 2009; Hsieh and Chen, 2009; Mishra, 2009; Nishida and Koike, 2009; Zadeh et al., 2009; Shin et al., 2009; Yang et al., 2009; Morton and Lee, 2011).
The Asian horseshoe crab does not have multiple utilization comparing to the Atlantic species but it has potential uses in biomedical and eco-tourism (Kassim et al., 2008). Quantitative data on the population of Malaysian horseshoe crabs are sparse. The population structure of juvenile and adult horseshoe crabs at the spawning ground remains unknown. Moreover, the public is unaware of the biological and ecological importance of these horseshoe crabs.
In Asia, horseshoe crab eggs are popularly served as a local delicacy (Morton and Blackmore, 2001). At Balok beach, gravid female horseshoe crabs are harvested to make into soup, curry and salad. Horseshoe crabs are also treated as by-catch and exported to Thailand where it fetches higher price (Christianus and Saad, 2009).
Commercial harvesting for food and production of Tachypleus Amebocyte Lysate (TAL), habitat and spawning area loss and coastal pollution are the major threats to Asian horseshoe crabs (Chen et al., 2004; Cartwright-Taylor et al., 2009; Hajeb et al., 2009). To date, no conservation practices, legislation or harvest regulations have been implemented to protect the horseshoe crabs in Malaysia as they are not listed in the Malaysian Wildlife Conservation Act 2010 (Act, 716).
This study aims to determine the size frequency distribution, length-weight relationships and sex ratios of horseshoe crab population in Balok Beach, Kuantan, Pahang, which is a spawning site located at the east coast of Peninsular Malaysia. Carapace condition and infestation of epibionts on the horseshoe crabs were observed. This study provides both qualitative and quantitative baseline information on the spawning population of T. gigas in Malaysia, which is crucial for future implementation of conservation measures and management planning.
MATERIALS AND METHODS
Study site: Study was conducted between May 2009 and April 2010, at Balok beach (3°56.257’N, 103°22.568’E; Fig. 1), a sandy estuarine beach in Kuantan, Pahang, located at the east coast of Peninsular Malaysia. Spawning pairs of T. gigas were found in May and June, during high tides of new moon and full moon (Zaleha et al., 2010; Kassim et al., 2008).
Balok beach experiences north-east monsoon between November and February (Chua, 1984) and mix semi-diurnal tides. According to Department of Survey and Mapping Malaysia (JUPEM, 2010), the tidal amplitude is approximately 2.28 m during spring tide and reaches a maximum wave height of 3 m during new and full moon phases.
Water analysis and sediment characteristics of study site
Physicochemical parameters of Balok Beach coastal water: During each sampling, physicochemical parameters of coastal water including water temperature, salinity, dissolved oxygen content and pH were measured in situ at a depth of 10 cm using a water quality monitoring probe (YSI, USA). Sediment grain size of Balok beach: Sand sediments from horseshoe crab nests collected in five cores (5 cm in diameter). Each core consisted of two measures, the surface sand sediment (0 to 5 cm depth) and sediment at a depth of 15 to 20 cm (Pooler et al., 2003). Grain size of sand sediment then analyzed using sieve-pipette method (Gee and Bauder, 1986) and wet-sieving through mesh sizes of 16 to 240 μm. Meanwhile, silt-clay content was determined using pipette method (Hsieh et al., 2004).
Sampling strategy: The horseshoe crabs were sampled for 6 consecutive days during every high tide of new and full moon phases between May 2009 and April 2010. Sampling was conducted during both day and night.
Random quadrate and transect technique was not suitable for this study site due to the small population size of T. gigas. Therefore, visual search technique used was based on sighting of horseshoe crab along the beach at high tide line. Searches were made for approximately 1.5 h prior to each high tide of new and full moons and covering a stretch of 300 m of the beach. Landing location of horseshoe crabs was recorded with a GPS (Global positioning system) device. The same location was re-visited during low tides to determine the presence of eggs nest.
During high tide, the horseshoe crab is partly buried in the sand or rapidly swimming in search for the right spot to nest. The horseshoe crab was gently handpicked out from the sand. Each horseshoe crab individual counted, sexed and measured for size (mm) and weight (g).
Determination of size-frequency distribution, length-weight relationship, age estimation and instar stage of horseshoe crab, sex ratio of horseshoe crab population at Balok beach: Fresh body weight of each individual was measured to the nearest 0.01 kg using weighing apparatus. Prosomal sizes (length and width), opisthosomal length, total length and telson length of each individual were measured according to Yamasaki et al. (1988) to the nearest millimeter (mm) using a measurement tape.
Each horseshoe crab was classified according to prosomal width size classes. The size-frequency distribution was expressed in bar chart with different interval size classes of 0-50, 51-100 mm, 101-150 mm and so on. Prosomal width of horseshoe crab was employed as a morphometric proxy to estimate age and instar stage based on size-age-growth relationship established by Sekiguchi et al. (1988a). Length and weight data analyzed according to Le Cren (1951) using hypothetical formula: W = aLb, where W is weight (g), L is carapace length (mm), parameter ‘a’ is proportionality constant and parameter ‘b’ is the allometric coefficient.
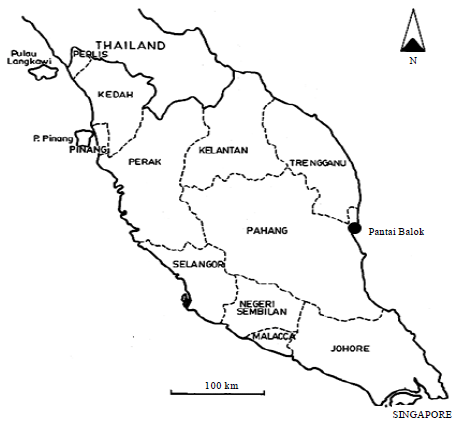 | |
| Fig. 1: | The study site at Balok Beach, Kuantan, Pahang, Malaysia |
The logarithm equation log W = log a + b log L calculated separately for both sexes and a straight line fitted to scatter diagram. Parameters ‘a’ and ‘b’ estimated based on logarithmically transformed expression where weight is the dependent variable and length is the independent variable (Cherif et al., 2008). Sex ratio determined according to morphological characteristics following Debnath (1991) and calculated for each month throughout the study period.
Epibiont infestation on horseshoe crab: Each horseshoe crab examined individually for the presence of epibionts. Each epibiont was removed using a scalpel and fixed in 10% formalin. The collected epibiont then identified to genus level in the laboratory.
Observation of horseshoe crab carapace condition: The condition of carapace, lateral eyes and telson of each horseshoe crab individual examined and summarized. Upon completion of data collection, all horseshoe crab individuals were return to the water.
Statistical analysis: Mean value for each morphometric parameter was compared between sexes using Student’s t-tests and a significant level of 95% probability was adopted (Chiu and Morton, 2003b). Normal distribution within each prosomal size group examined using Kolmogorov-Smirnov test. Student’s t-test was used to determine the significance at 95% probability level. Covariance analysis was employed to describe difference in the regression of logarithmic value of weight on logarithmic value of carapace length (Debnath and Choudhury, 1988; Srijaya et al., 2010b). Chi-squared test was applied to determine deviation of sex ratios from 1 (Cartwright-Taylor et al., 2009).
RESULTS AND DISCUSSION
Species determination, size-frequency distribution, length-weight relationship, age estimation and instar stage of horseshoe crab, sex ratio of horseshoe crab population at Balok beach: Based on morphologically characteristics following Yamasaki et al. (1988), the horseshoe crab species found at Balok Beach was identified as Tachypleus gigas. In total, 161 T. gigas (86 males and 75 females) were sampled throughout the study period. The total number of horseshoe crabs collected varied every month. Male individuals outnumbered the females because they tend to re-visit the spawning beach more often than the female (Lovel and Botton, 1992). Hence, males were the dominant sex at the spawning site.
| Table 1: | Mean and range (bracketed values) of various morphometric parameters of Tachypleus gigas examined at Balok Beach, Kuantan, Pahang, Malaysia |
 | |
| *Significant differences between the mean values of the two sexes using Student’s t-tests at 95% probability level | |
Most horseshoe crabs came ashore in amplexus pairs. There was occurrence of solitary males and two mating pairs in tandem where two males were coupling with one behind the other male and a female was at front of the males. The satellite male was not observed at this site. This is probably due to the small population size therefore lack of satellite male and group spawning (Botton et al., 1996; Brockmann and Smith, 2009).
The largest prosomal width sampled male and female were 241 and 320 mm, respectively. Sexual dimorphism characteristic of horseshoe crab is obviously on the size difference. Females are prominently bigger than the males (Yamasaki et al., 1988; Key et al., 1996; Brockmann and Smith, 2009). The females were bigger in size than the males with mean prosomal length and width of 154 and 246 mm, respectively (Table 1). Matured females were heavier than the males with mean body weight of 824 g (Table 1) due to greater growth in each molt (Gerhart, 2007) and numerous egg masses in prosomal cavity (Graham et al., 2009). Table 1 showed significant differences (p<0.05) in prosomal size (length and width), telson length, carapace length (prosomal length) and body weight between the two sexes.
Size-frequency distribution of Tachypleus gigas at Balok Beach, Kuantan, Pahang: Size-frequency data indicated size groups of 151-200 mm for males and 201-250 mm for females (Fig. 2). Male T. gigas is smaller than the respective female. Based on prosomal widths, approximately 69% of male horseshoe crabs ranged from 151-200 mm and 53% of females were ranging from 201-250 mm. Mean values obtained from prosomal widths of both sexes showed significant differences (p<0.05). The size-frequency distribution between both sexes was not normally distributed.
Length-weight relationship of horseshoe crab population at Balok Beach, Kuantan, Pahang: The parabolic equations were determined from the length-weight data:
| • | Male: | W = 0.0018 L2.557 |
| • | Female: | W = 0.0005 L2.837 |
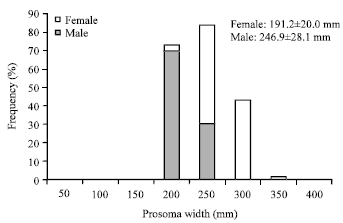 | |
| Fig. 2: | Frequency of size classes of Tachypleus gigas at Balok Beach, Kuantan, Pahang |
The length-weight relationships were linear for both sexes. The equations were shown in the following:
| • | Male: | log W = 2.557 log L-2.750 |
| • | Female: | log W = 2.837 log L-3.303 |
The values of the slopes (b) for both sexes were more than 1 thus exhibited a positive allometry where increment of weight was more rapid as compared to prosomal length. Covariance analysis revealed significant difference between the slopes of both sexes (F = 3085.391 with df 1,160; p<0.05). Weights of males and females increased with exponentials of 2.557 and 2.837, respectively.
Length-weight relationship varies with water temperature, food availability, habitat type and reproductive activities. Fish weight and b-value increase with sufficient feeding and gonad development. The b-value is also an indicator of food intake and growth pattern (Arslan et al., 2004). An animal living in inadequate water temperature and feed insufficiently would demonstrate a negative allometric growth (Arslan et al., 2004).
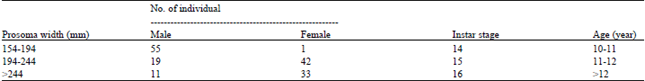
Age estimation of horseshoe crabs at Balok Beach, Kuantan, Pahang: All horseshoe crabs examined at Balok Beach were adult individuals. Based on size-age-growth relationship established by Sekiguchi et al. (1988b), the horseshoe crabs were grouped into fourteenth and fifteenth instar stages, with prosomal width sizes ranged from 154-194 mm and 194-244 m, respectively. Individual classified into fourteenth instar stage was estimated to have survived for 10 to 11 years whereas the fifteenth instar stage group was estimated to have lived for 11 to 12 years. However, individual with prosomal width of more than 244 m was regarded as being in the sixteenth instar stage and presumed to have lived for more than 12 years. Approximately 64% of male individuals with prosomal widths ranged between 154 and 194 m and 55% of female individuals belonged to prosomal width size group ranging from 194 to 244 mm (Table 2).
In previous study conducted by Hajeb et al. (2005), a total of 54 males and 49 females T. gigas were sampled and measured. About 74% of male individuals categorized into prosomal widths group ranging from 154-194 mm hence grouped into fourteenth instar stage and estimated to have lived form 10 to 11 years. Meanwhile, 71% of female individuals in prosomal width size group of 194-244 mm were fifteenth instars and presumed to have lived for 11 to 12 years. There was no record of horseshoe crab individual in size group of more than 244 mm. According to Morton and Lee (2011), locality and salinity affect the increment of prosomal width of horseshoe crab species.
Sex ratio of horseshoe crab population at Balok Beach, Kuantan, Pahang: Fewer horseshoe crabs were sighted during north-east monsoon (November to March) due to occurrence of heavy precipitation, strong surf and flooding at the study site. Strong offshore winds and low water temperature deter the horseshoe crabs from approaching the nesting beach (Gerhart, 2007). Horseshoe crab favors low wave height and swash velocity to increase spawning and hatching success of the eggs (Jackson et al., 2010).
Most horseshoe crabs came ashore at Balok Beach in amplexed pairs. There were two occurrences of tandem amplexus. Neither unattached female nor satellite male was sighted. However, there was sighting of unattached males swimming nearshore. According to Zaleha et al. (2010), horseshoe crab spawning season varies with latitude and spawning peaks reported in May and June at the east coast of Peninsular Malaysia. However, spawning peaks observed in this study for 2009 were in May and September (Table 3). There was no large spawning aggregation at Balok Beach and spawning activity occurred year round except during monsoon season.
As compared to female, higher proportion of returning males were observed. The sex ratio (Table 3) was not significantly different from 1 except in September 2009 and March 2010 (p<0.05). The null hypothesis on the sex ratio was equivalent to 1 per month of the year cannot be rejected except in July, September, December 2009 and January to April 2010 where the sex ratios were not more than 1 when males outnumbered females and thus suggesting an active breeding period. Sex ratio of 1:1 suggests a consequence of low population density (Botton et al., 1996) or a maximized reproduction and a full reproductive capacity of horseshoe crabs (Reyes, 2009).
| Table 2: | Age and instar stage estimation using prosomal widths (mm) of Tachypleus gigas examined at Balok Beach based on size-age-growth relationship established by Sekiguchi et al. (1988b) |
 | |
| Table 3: | Male to female ratios of horseshoe crab Tachypleus gigas sampled at Balok Beach between May 2009 to April 2010 |
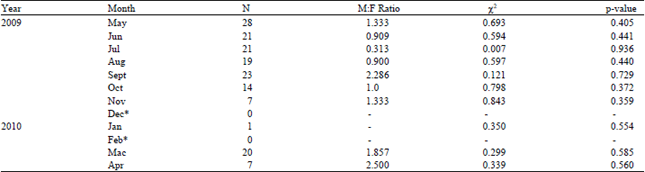 | |
| *No horseshoe crab sighting | |
Fisher’s principle on 1:1 (male to female ratio) is evolutionarily stable but it is not common in horseshoe crab population because male tends to re-visit the nesting beach more often than the female. Sex ratio can be influenced by the harvesting of females horseshoe crab as delicacy by the local community at Balok Beach.
Tachypleus and Carcinoscorpius show insignificant or no reproductive competition. Asian horseshoe crab lacks of satellite male and spawning aggregation (Brockmann and Smith, 2009) thus demonstrates monogamous mating pattern where operational sex ratio is 1:1 (Mattei et al., 2010).
Infestation of epibiont on Tachypleus gigas: A total of 45% of male individuals were fouled. Adult male was prone to epibiotic infestation as compared to female (Table 4) due to more molt frequent of females before reaching maturity (Sekiguchi et al., 1988a) and nesting behavior makes females less vulnerable to epibiont. Only sexually matured adult horseshoe crabs are infested by epibiont as they have stopped molting (Botton, 2009). However, in culture condition, this kind of epibiotic infestation was not observed on larvae and juvenile of T. gigas (Faizul et al., 2011). Horseshoe crabs are generally burrowing shallowly in the sediment substrates when resting, foraging or spawning (Sekiguchi and Shuster, 2009). In amplexus position, the prosoma carapace of female is buried at the level of lateral eyes into the sand during spawning and opisthosoma carapace is covered by the male holding onto the female. Because spawning horseshoe crabs remain amplexed for a considerable time therefore male body is vulnerable to epibiont colonization (Patil and Anil, 2000).
Throughout the study period, four epibiont group were found on dorsal and ventral carapaces of Tachypleus gigas namely acorn barnacle Balanus, pedunculate barnacle Octolasmis, conical slipper shell Calyptraea and flat slipper shell Crepidula. Barnacle, polychaete and mollusk are common epibionts infesting horseshoe crabs (Botton, 2009; Tan et al., 2011).
Observation of horseshoe crab carapace condition: The condition of carapaces, lateral eyes and telsons of all horseshoe crab samples were examined and classified accordingly (Table 5). Approximately 8% of males had covered eyes and damaged carapaces and 15% of females with broken telsons (Table 5). Meanwhile, damaged carapace and broken telson was 19%, this is possibly due to collision with boats since the study site is near to boat landing jetty.
At the study site, males with encrusted prosoma and eyes were found attached to females as amplexus pairs. This is in contrast with the report by Brockmann and Penn (1992) that an attached male is less likely to have its prosoma and eye encrusted by fouling organisms. Duffy et al. (2006) also speculated that a male horseshoe crab with deteriorated lateral eyes is not fully excluded in mating. The male is either an attached or satellite individual. There were attached males with severely damaged eyes in present study. Poor eye condition is defined if one or both eyes are rough to be touched and blocked by fouled organisms (Schwab and Brockmann, 2007). In this study, only 9% of males were observed with poor eye condition.
Carcasses of horseshoe crabs were found onshore as the animals are treated as by-catch. The fisherman removes the horseshoe crabs entangled in fishing net and tosses the animals onshore. The animals usually die due to desiccation after prolong exposure to high temperature and fail to overturn. Male in good condition is more likely to right itself using telson and at low risk to be stranded when the tide recedes (Brockmann and Smith, 2009).
| Table 4: | Number and percentage of horseshoe crab individuals infested by epibiont |
 | |
| Table 5: | Individual horseshoe crab Tachypleus gigas with notable physical condition |
 | |
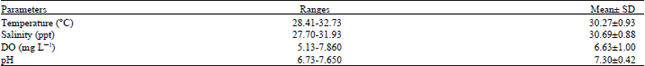
| Table 6: | Mean and standard deviation and range of physicochemical water parameters at Balok Beach, Kuantan, Pahang, Malaysia |
 | |
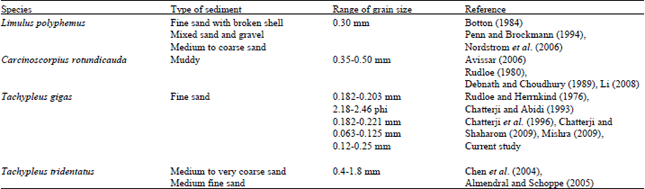
| Table 7: | Comparison between type of sediment and grain size of spawning sites of different horseshoe crab species |
 | |
As described by Botton and Loveland (1989), a female horseshoe crab sampled at study site was found with a stubby telson and a male has a curved telson. Horseshoe crab with telson abnormality is at risk of become stranded once overturned by waves.
A female horseshoe crab was found with damaged fifth walking appendage. The walking appendages have higher tendency of damage because horseshoe crab uses the appendages to push itself into the sand sediment (Duffy et al., 2006).
Physicochemical parameters of Balok Beach coastal water: Physicochemical parameters of water including temperature, salinity, dissolved oxygen content and pH were measured in situ at Balok coastal water during May 2009 and April 2010. Table 6 showed the measured physicochemical parameters at the study area.
Sediment grain size of Balok Beach: The sand sediment of study site comprises 93.78% of fine sands with mean grain size ranged from 120 to 250 μm and silt-clay content is 5.58%. Each horseshoe crab species prefers a specific range of sediment grain size for nesting site (Table 7). Medium grain size is most ideal for the aeration of fertilized eggs (Rudloe and Herrnkind, 1976). It is reported that the moderate sorted sediment promotes hatching and survival of larvae (Chiu and Morton, 2003c). Shuster (1985) suggested sediment size and sorting affect both water infiltration and exfiltration through the beach. While, Rudloe and Herrnkind (1976) noted that an increase in the grain size does not favor horseshoe crab nesting.
CONCLUSION
The sand sediment of Pantai Balok mainly is made of fine sands with mean grain size ranged from 120 to 250 μm. Horseshoe crab nests can be found at study site after the tide recedes.
Out of the 86 males and 75 female horseshoe crabs T. gigas sampled, 49 were amplexus pairs. The size of the largest male and female measured were 241 and 320 mm, respectively.
More males were found ashore hence the sex ratio was male biased. In comparison to the Atlantic horseshoe crab, occurrence of satellite male was not observed. Local community harvests the female horseshoe crabs for two purposes, own consumption and income source (sold to middleman for exports to Thailand). Based on prosomal width size classes, the horseshoe crabs were found to be fourteenth to sixteenth instars.
Fewer horseshoe crabs were sighted during north-east monsoon due to the occurrence of heavy precipitation, strong surf and flooding at the study site. Spawning peaks were in May and September. Males were more prone to epibiont infestation than females. This is basically due to its nesting behavior, whereby males carapace were exposed to the environment of epibiont. Four epibiont species found on the carapaces of these horseshoe crabs were acorn barnacle Balanus, pedunculate barnacle Octolasmis, conical slipper shells Calyptraea and flat slipper shells Crepidula. Considering the study site is near to fish landing jetty, it is not a surprise to discover horseshoe crabs with damaged eyes, carapaces and telsons, possibly due to collision with boats.
ACKNOWLEDGMENT
Authors would like to thank the Department of Aquaculture and Institute of Bioscience, Universiti Putra Malaysia for providing sampling apparatus, laboratorial equipment and facilities for this study.
REFERENCES
- Almendral, M.A. and S. Schoppe, 2005. Population structure of Tachypleus tridentatus (Chelicerata: Merostomata) at a nursery beach in Puerto Princesa City, Palawan, Philippines. J. Nat. Hist., 39: 2319-2329.
CrossRef - Avissar, N.G., 2006. Modeling potential impacts of beach replenishment on horseshoe crab nesting habitat sustainability. Coast. Manage., 34: 427-441.
CrossRef - Botton, M.L., 1984. Diet and food preferences of adult horseshoe crab Limulus polyphemus in Delaware Bay, New Jersey, USA. Mar. Biol., 81: 199-207.
CrossRef - Botton, M.L., 2000. Toxicity of cadmium and mercury to horseshoe crab (Limulus polyphemus), embryos and larvae. Bull. Environ. Contam. Toxicol., 64: 137-143.
CrossRef - Botton, M.L. and R.E. Loveland, 1989. Reproductive risk: High mortality associated with spawning by horseshoe crabs (Limulus polyphemus) in Delaware Bay, USA. Mar. Biol., 101: 143-151.
CrossRef - Botton, M.L., C.N. Shuster, K. Sekiguchi and H. Sugita, 1996. Amplexus and mating behavior in the Japanese horseshoe crab, Tachypleus tridentatus. Zool. Sci., 13: 151-159.
CrossRef - Brockmann, H.J. and D. Penn, 1992. Male mating tactics in horseshoe crab, Limulus polyphemus. Anim. Behav., 44: 653-665.
CrossRef - Burger, J., C. Dixon, T. Shukla, N. Tsipoura and H. Jensen et al., 2003. Metals in horseshoe crabs from Delaware Bay. Arch. Environ. Contam. Toxicol., 44: 36-42.
CrossRef - Carmichael, R.H., D. Rutecki and I. Valiela, 2003. Abundance and population structure of the Atlantic horseshoe crab Limulus polyphemus in Pleasant Bay, Cape Cod. Mar. Ecol. Prog. Ser., 246: 225-239.
CrossRef - Cartwright-Taylor, L., J. Lee and C.C. Hsu, 2009. Population structure and breeding pattern of the mangrove horseshoe crab Carcinoscorpius rotundicauda in Singapore. Aquat. Biol., 8: 61-69.
CrossRefDirect Link - Chatterji, A., A.H. Parulekar and S.Z. Qasim, 1996. Nesting Behavior of the Indian Horseshoe Crab, Tachypleus gigas (Muller) (Xiphosura). In: India's Exclusive Economic Zone: Resources, Exploitation, Management, Qasim, S.Z. and G.S. Roonwal (Eds.). Omega Scientific Publishers, New Delhi, India, pp: 142-155.
- Chatterji, A. and F. Shaharom, 2009. Unusual spawning behavior of the horseshoe crab (Tachypleus gigas, Muller) after the Tsunami along Orissa Coast, India. Pertanika J. Sci. Tech., 17: 263-268.
Direct Link - Chen, C.P., H.Y. Yeh and P.F. Lin, 2004. Conservation of the horseshoe crab at Kinmen Taiwan: Strategies and practices. Biodiversity Conservat., 13: 1889-1904.
Direct Link - Cherif, M., R. Zarrad, H. Gharbi, H. Missaoui and O. Jarboui, 2008. Length-weight relationships for 11 fish species from the Gulf of Tunis (SW Mediterranean Sea, Tunisia). Pan-Am. J. Aquatic Sci., 3: 1-5.
Direct Link - Chiu, H.M.C. and B. Morton, 2003. The morphological differentiation of two horseshoe crab species, Tachypleus tridentatus and Carcinoscorpius rotundicauda (Xiphosura), in Hong Kong with a regional Asian comparison. J. Nat. Hist., 37: 2369-2382.
CrossRef - Duffy, E.E., J.P. Dustin, M.L. Botton, H.J. Brockmann and R.E. Loveland, 2006. Eye and clasper damage influence male mating tactics in the horseshoe crabs, Limulus polyphemus. J. Ethol., 24: 67-74.
CrossRef - Graham, L.J., M.L. Botton, D. Hata, R.E. Loveland and B.R. Murphy, 2009. Prosomal-width-to-weight relationships in American horseshoe crabs (Limulus polyphemus): Examining conversion factors used to estimate landings. Fish. Bull., 107: 235-243.
Direct Link - Hajeb, P., A. Christianus, A. Ismail, S. ShakibaZadeh and C.R. Saad, 2009. Heavy Metal Concentration in Horseshoe Crab (Carcinoscorpius Rotundicauda and Tachypleus Gigas) Eggs from Malaysian Coastline. In: Biology and Conservation of Horseshoe Crabs, Tanacredi, J.T., M.L. Botton and D.R. Smith (Eds.). Springer, New York, USA., pp: 455-464.
- Hajeb, P., S. ShakibaZadeh, A. Christianus, A. Arshad and C.R. Saad, 2005. Growth stages and age of the horseshoe crabs, Carcinoscorpius rotundicauda and Tachypleus gigas, in Malaysia. Proceedings of KUSTEM 4th Annual Seminar on Sustainability Science and Management, May 2-5, 2005, Malaysia, pp: 185-190.
- Hsieh, H.L. and C.P. Chen, 2009. Conservation Program for the Asian Horseshoe crab Tachypleus Tridentatus in Taiwan: Characterizing the Microhabitat of Nursery Grounds and Restoring Spawning Grounds. In: Biology and Conservation of Horseshoe Crabs, Tanacredi, J.T., M.L. Botton and D.R. Smith (Eds.). Springer, New York, USA., pp: 417-438.
- Jackson, N.L., K.F. Nordstrom and D.R. Smith, 2010. Armoring of Estuarine Shorelines and Implications for Horseshoe Crabs on Developed Shorelines in Delaware Bay. In: Puget Sound Shorelines and the Impacts of Armoring, Shipman, H., M.N. Dethier, G. Gelfenbaum, K.L. Fresh and R.S. Dinicola (Eds.). U.S. Department of the Interior, U.S. Geological Survey, USA., pp: 195-202.
- Zaleha, K., I. Hazwani, H.S. Hamidah, B.Y. Kamaruzzaman and K.C.A. Jalal, 2011. Effect of salinity on the egg hatching and early larvae of horseshoe crab Tachypleus gigas (Muller, 1785) in laboratory culture. J. Applied Sci., 11: 2620-2626.
CrossRefDirect Link - Zaleha, K., B.Y. Kamaruzzaman, B.A. John and M.C. Ong, 2010. Cd, Cu and Pb concentration levels in horseshoe crab nesting grounds of pahang coast, Malaysia. J. Biol. Sci., 10: 790-794.
CrossRef - Key Jr., M.M., W.B. Jeffries, H.K. Voris and C.M. Yang, 1996. Epizoic bryozoans, horseshoe crabs and other mobile benthic substrates. Bull. Mar. Sci., 58: 368-384.
Direct Link - Le Cren, E.D., 1951. The length-weight relationship and seasonal cycle in gonad weight and condition in the perch (Perca fluviatilis). J. Anim. Ecol., 20: 201-219.
CrossRefDirect Link - Lee, C.N. and B. Morton, 2005. Experimentally derived estimates of growth by juvenile Tachypleus tridentatus and Carcinoscorpius rotundicauda (Xiphosura) from nursery beaches in Hong Kong. J. Exp. Mar. Biol. Ecol., 318: 39-49.
CrossRef - Lee, C.N. and B. Morton, 2009. Emergence Behavior of Juvenile Tachypleus tridentatus under Simulated Tidal Conditions in the Laboratory and at two different Sediment Temperatures. In: Biology and Conservation of Horseshoe Crabs, Tanacredi, J.T., M.L. Botton and D.R. Smith (Eds.). Springer, New York, USA., pp: 275-283.
- Lovel, R.E. and M.L. Botton, 1992. Size dimorphism and the mating system in horseshoe crabs, Limulus polyphemus L. Anim. Behav., 44: 907-916.
Direct Link - Mattei, J.H., M.A. Beekey, A. Rudman and A. Woronik, 2010. Reproductive behavior in horseshoe crabs: Does density matter? Curr. Zool., 56: 634-642.
Direct Link - Faizul, M.M.I., A. Christianus, S.M.N. Amin, S. Shafeeqa and T. Rishzuan, 2011. Evaluation of growth performance of Tachypelus gigas (Muller, 1785) under two different culture system. J. Fish. Aquatic Sci., 6: 788-794.
CrossRefDirect Link - Morton, B. and C.N. Lee, 2011. Spatial and temporal distribution of juvenile horseshoe crabs (Arthropoda: Chelicerata) approaching extirpation along the northwestern shoreline of the New Territories of Hong Kong SAR, China. J. Nat. History, 45: 227-251.
CrossRef - Nishida, S. and H. Koike, 2009. Genetic Structure of Japanese Populations of Tachypleus tridentatus by mtDNA AT-Rich Region Sequence Analysis. In: Biology and Conservation of Horseshoe Crabs, Tanacredi, J.T., M.L. Botton and D. Smith (Eds.). Springer US. New York, ISBN-13: 978-0-387-89958-9, pp:183-196.
- Nordstrom, K.F., N.L. Jackson, D.R. Smith and R.G. Weber, 2006. Transport of horseshoe crab eggs by waves and swash on an estuarine beach: Implications for foraging shorebirds. Estuarine Coastal Shelf Sci., 70: 438-448.
CrossRef - Patil, J.S. and A.C. Anil, 2000. Epibiotic community of the horseshoe crab Tachypleus gigas. Mar. Biol., 136: 699-713.
CrossRefDirect Link - Penn, D. and H.J. Brockmann, 1994. Nest-site selection in the horseshoe crab, Limulus polyphemus. Biol. Bull., 187: 373-384.
Direct Link - Pooler, P.S., D.R. Smith, R.E. Loveland, M.L. Botton and S.F. Michels, 2003. Assessment of sampling methods to estimate horseshoe crab (Limulus polyphemus L.) egg density in Delaware Bay. Fish. Bull., 101: 698-703.
Direct Link - Rudloe, A. and W.F. Herrnkind, 1976. Orientation of Limulus polyphemus in the vicinity of breeding beaches. Mar. Behav. Physiol., 4: 75-89.
CrossRef - Schwab, R.L. and H.J. Brockmann, 2007. The role of visual and chemical cues in the mating decisions of satellite male horseshoe crabs, Limulus polyphemus. Anim. Behav., 74: 837-846.
CrossRefDirect Link - Sekiguchi, K., H. Seshimo and H. Sugita, 1988. Post-embryonic development of the horseshoe crab. Biol. Bull., 174: 337-345.
Direct Link - Sekiguchi, K. and C.N. Shuster, 2009. Limits on the Global Distribution of Horseshoe Crabs (Limulacea): Lessons Learned from Two Lifetimes of Observations: Asia and America. In: Biology and Conservation of Horseshoe Crabs, Tanacredi, J.T., M.L. Botton and D. Smith (Eds.). Springer US, New York, ISBN-13: 978-0-387-89958-9, pp: 5-24.
- Seino, S., T. Uda, Y. Tsuchiya and K. Tsuchiya, 2004. Conservation history of horseshoe crab Tachypleus tridentatus and its spawning ground: A designated natural monument in Kasaoka Bay in Okayama Prefecture. Proceedings of the 2nd International Conference Asian and Pacific Coast, February 29- March 4, 2004, Makuhari, Japan, pp:1-14.
- Zadeh, S.S., A. Christianus, C.R. Saad, P. Hajeb and M.S. Kamarudin, 2009. Comparisons in Prosomal Width and Body Weight Among Early Instar Stages of Malaysian Horseshoe Crabs Carcinoscorpius rotundicauda and Tachypleus gigas in the Laboratory. In: Biology and Conservation of Horseshoe Crabs, Tanacredi, J.T., M.L. Botton and D. Smith (Eds.). Springer US, New York, ISBN-13: 978-0-387-89958-9, pp: 267-274.
- Shuster, C.N., 1982. A Pictorial Review of the Natural History and Ecology of the Horseshoe Crab Limulus Polyphemus, with Reference to other Limulidae. In: Physiology and Biology of Horseshoe Crabs, Bonaventura, J., C. Bonaventura and S. Tesh, (Eds.). Progress in Clinical and Biological Research, New York, USA., pp: 1-53.
- Shuster, Jr. C.N., 1985. Introductory Remarks on the Distribution and Abundance of the American Horseshoe Crabs, Limulus Polyphemus, Spawning in the Chesapeake Bay Area. In: The Chesapeake: Prologue to the Future: Proceedings from the Chesapeake Bay Symposium, Chase, V. (Ed.). National Aquarium Baltimore, Baltimore, Maryland, pp: 34-38.
- Srijaya, T.C., P.J. Pradeep, S. Mithun, A. Hassan, F. Shaharom and A. Chatterji, 2010. A new record on the morphometric variations in the populations of horseshoe crab (Carcinoscorpius rotundicauda Latreille) obtained from two different ecological habitats of Peninsular Malaysia. Our Nat., 8: 204-211.
CrossRefDirect Link - Srijaya, T.C., P.J. Pradeep, S. Mithun, A. Hassan, F. Shaharom and A. Chatterji, 2010. Variabilities in different body measurements of the horseshoe crab, Carcinoscorpius rotundicauda (Latreille) collected from Setiu and Gelang Patah habitats in Peninsular Malaysia. J. Bombay Nat. Hist. Soc., 107: 130-134.
Direct Link - Widener, J.W. and R.B. Barlow, 1999. Decline of a horseshoe crab population on cape cod. Biol. Bull., 197: 300-302.
Direct Link - Yang, M.C., C.P. Chen, H.L. Hsieh, H. Huang and A.C.L. Chen, 2009. Phylogeography, Demographic History and Reserves Network of Horseshoe Crab, Tachypleus Tridentatus, in the South and East China Seabeds. In: Biology and Conservation of Horseshoe Crabs, Tanacredi, J.T., M.L. Botton and D.R. Smith (Eds.). Springer, New York, USA., pp: 163-181.
- Zhou, H. and B. Morton, 2004. The diets of juvenile horseshoe crabs, Tachypleus tridentatus and Carcinoscorpius rotundicauda (Xiphosura), from nursery beaches proposed for conservation in Hong Kong. J. Nat. History, 38: 1915-1925.
CrossRef