
R. Siva Sankar
Department of Ecology and Environmental Sciences, Pondicherry University, 605 014, Puducherry, India
A. Yogamoorthi
Department of Ecology and Environmental Sciences, Pondicherry University, 605 014, Puducherry, India
Pakistan Journal of Biological Sciences
Year: 2012 | Volume: 15 | Issue: 10 | Page No.: 490-495







ABSTRACT
Free amino acid plays an important role in physiological functions. The ghost crab, Ocypode platytarsis caught off the Puducherry sandy beaches, south east coast of India were investigated for the pattern of distribution of free amino acids. The hemolymph and muscle were sampled from male and female crabs and then subjected to free amino acid analysis quantitatively by HPLC. Fourteen free amino acids have been determined in hemolymph and muscle of both sexes in Ocypode platytarsis. The essential amino acids were leucine, lysine, methionine, phenylalanine, threonine, valine and histidine and non-essential amino acids were alanine, arginine, glutamic acid, glycine, serine and tyrosine. The total concentration of free amino acid found in the male and female hemolymph was lower than that in the muscle. Glutamic acid is of higher concentration in the hemolymph whereas histidine found was higher in the muscle of both sexes. The male crab contains a strikingly higher concentration of histidine of about eight times than that of female in muscles. The study revealed that distribution pattern of free amino acids differ in relation to hemolymph and muscle of male and female crabs.
PDF Abstract XML References Citation
Received: July 15, 2012;
Accepted: August 27, 2012;
Published: October 06, 2012
How to cite this article
R. Siva Sankar and A. Yogamoorthi, 2012. Free Amino Acid Composition in Hemolymph and Muscle of the Ghost Crab, Ocypode platytarsis. Pakistan Journal of Biological Sciences, 15: 490-495.
DOI: 10.3923/pjbs.2012.490.495
URL: https://scialert.net/abstract/?doi=pjbs.2012.490.495
DOI: 10.3923/pjbs.2012.490.495
URL: https://scialert.net/abstract/?doi=pjbs.2012.490.495
INTRODUCTION
The concentrations of total amino acids were greater in the marine species than in the fresh water forms (Camien et al., 1951). Free amino acids are one of the most important fractions of non-protein nitrogen in crustaceans (Capillas, 2004). The characteristic of interesting levels of free amino acids with increasing evolutionary development was first observed by (Duchateau and Florkin, 1958). The role played by free amino acids in isosmotic intracellular regulation has been illustrated in several investigations (Schoffeniels and Gilles, 1970a, b; Weber and van Marrewijk, 1972a, b) and it can be an important source of energy producing compounds (Van Marrewijk and Ravestein et al., 1974). In addition, FAAs play important roles in physiological functions such as osmoregulation and buffer capacity in the tissues of aquatic animals (Van Waarde, 1988) and some amino acids are involved in neurotransmission (Anaya and Aniello, 2006). The non-protein amino acids in Crustacean muscle are essentially intracellular (Camien et al., 1951). They may provide an alternative organic energy reserve in nonfeeding stages (Bursell, 1966) as well as play a central role in protein synthetic and degradative processes (Skinner, 1965, 1966; Welinder, 1974, 1975). The objective of the present study mainly focused on free amino acid profile in haemolymph and muscle of Ocypode platytarsis in both sexes.
MATERIALS AND METHODS
Sampling site: The sampling site was along the sandy beaches of chinakalapet (Fig. 1), occupying the highest high tide level of the intertidal zone situated at south east coast of India.
Sampling method: Only larger sizes of healthy crabs were collected through hands by dislodging them from their burrows of 1 m depth during day time with the assistance of fisherman and transported to the laboratory alive in a closed plastic bag. Before the experiment was begun, the crabs were weighed and divided based on their sex.
Collection of haemolymph and muscle: The live animals were allowed to bleed completely and rapidly from each walking leg by cutting with a fine sterile scissor. Haemolymph samples from six individuals were collected and a known quantity of fresh tissue obtained by pooling and blending the abdominal muscle of 10 crabs.
 | |
| Fig. 1: | The location of the sampling site |
Haemolymph and muscle sample preparation and analysis: An equal volume of 5% Sulfosalicylic Acid (SSA) was added to the haemolymph for protein precipitation. Samples were centrifuged at 13,000 xg for 15 min at 1°C, then 100 μL of supernatant was taken for amino acid analysis (Findley and Stichle, 1978).
Twenty milliliter of 6% Trichloroacetic Acid (TCA) was added with muscle sample. They were homogenized, sonicated for 3 min (at 50% duty cycle) and centrifuged at 14,000 xg for 20 min at 1% C. After the supernatant was removed, the residue was re-extracted twice with TCA as above. All the supernatant was pooled, added to 30 mL of ether, shaken for 30 sec and the aqueous layer was concentrated in a low pressure rotary evaporator under 40°C to a fairly highly viscous fluid, then diluted with deionized double distilled water for amino acid analysis (Shewbart et al., 1972; McCoid et al., 1984; Via, 1986).
Amino acids analysis was carried out using High Performance Liquid Chromatography (HPLC) as done by Fang et al. (1992).
RESULTS
The concentration of 14 free amino acids in the haemolymph and muscle of both the sexes of crab, O. platytarsis was taken for inference.
Amino acid profile of male haemolymph: Glutamic acid is the compound at highest concentration with (17.5 μM mL-1) followed by histidine (6.4 μM mL-1), lysine (4.4 μM mL-1) and threonine (4.0 μM mL-1). Arginine (0.6 μM mL-1), valine (0.7 μM mL-1), methionine (0.9 μM mL-1) were detected in very lower concentrations. However, free amino acids such as serine, glycine, alanine, tyrosine and phenylalanine occur at relative concentrations between 1.1-1.8 μM mL-1. The concentration of serine was equal to phenylalanine (Fig. 2).
Amino acid profile of female haemolymph: Concentration of glutamic acid (10.8 μM mL-1) was high followed by histidine (9.6 μM mL-1) and threonine (4.1 μM mL-1). Serine (1.3 μM mL-1) and leucine (1.7 μM mL-1) occur at relative concentrations (Fig. 2).
Amino acid profile of male muscle: The most striking finding was the presence of very high concentrations of histidine (301.2 μM mL-1) in the muscle of male crabs. The concentration of threonine (66.3 μM mL-1) is high after histidine followed by arginine (60.7 μM mL-1), alanine (50.2 μM mL-1), glutamic acid (24.0 μM mL-1), glycine (22.2 μM mL-1), tyrosine (15.8 μM mL-1), lysine (9.5 μM mL-1), aspartic acid (6.0 μM mL-1) and valine (1.8 μM mL-1).
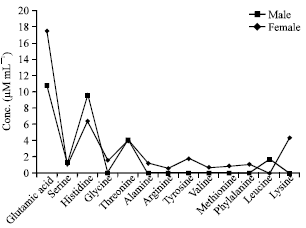 | |
| Fig. 2: | Amino acid contents in male and female hemolymph of O. platytarsis |
The concentration of serine (4.5 μM mL-1) was similar to that of leucine. Among all amino acids, methionine (3.3 μM mL-1) was very much less in concentration (Fig. 3).
Amino acid profile of female muscle: The presence of high concentration of histidine (37.1 μM mL-1) was found, followed by glutamic acid (16.7 μM mL-1), threonine (11.6 μM mL-1) and alanine (4.4 μM mL-1). Equal concentration of serine (1.7 μM mL-1) and leucine was present followed by arginine (1.0 μM mL-1). The lower concentration of tyrosine (0.8 μM mL-1) was detected followed by aspartic acid (0.7 μM mL-1) and lysine (0.6 μM mL-1) and all the three were of relative concentrations (Fig. 3).
Comparison of male and female haemolymph: The enumeration of free amino acids was determined to be nine and five in male and female respectively. Glutamic acid, serine, histidine and threonine were found to be present in male whereas leucine was detected in female (Fig. 2). Glutamic acid and threonine concentrations were high in male and serine and histidine was reported to be on the higher side in female. Threonine, one among the FAA occurs at relative concentrations in both the sex.
Comparison of male and female muscle: There were thirteen amino acids present in the muscle of male crabs whereas ten in the muscle of the female. Amino acids such as glycine, valine and methionine were found only in male and not in female. The concentration of total FAA was estimated to be higher in the muscle of male. With respect to the quantity of amino acids in male, only histidine is noteworthy and is eight times higher in concentration than that of the female (Fig. 3).
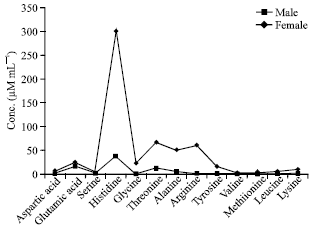 | |
| Fig. 3: | Amino acid contents in male and female muscle of O. platytarsis |
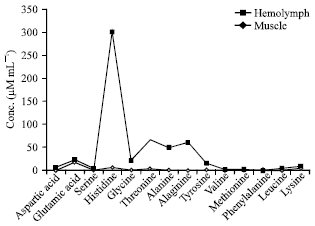 | |
| Fig. 4: | Amino acid contents in hemolymph and muscle of male in O. platytarsis |
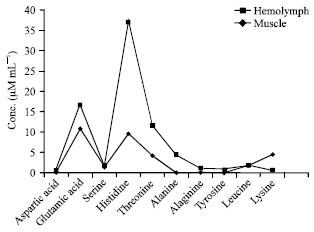 | |
| Fig. 5: | Amino acid contents in hemolymph and muscle of female in O. platytarsis |
Comparison of haemolymph and muscle of male: Thirteen free amino acids were determined in muscle of male and twelve in haemolymph. Aspartic acid and leucine were found only in the muscle part and phenylalanine was determined only in haemolymph and not in the muscle (Fig. 4). The most striking finding was the presence of histidine level in muscle, nearly fifty times higher than that of haemolymph. The total FAA concentrations found in the muscle was comparatively higher than haemolymph.
Comparison of haemolymph and muscle of female: Five amino acids from haemolymph and ten amino acids from the muscle were reported. Among these, aspartic acid, alanine, arginine and tyrosine were only present in the muscle. Level of serine from haemolymph and muscle occur at relative concentrations. The concentration of leucine from haemolymph and muscle were equal (Fig. 5). The total FAA concentrations found in the muscle were higher than that of haemolymph.
DISCUSSION
Fourteen free amino acids have been determined in hemolymph and muscle of both sexes of O. platytarsis. Among these, the essential amino acids were leucine, lysine, methionine, phenylalanine, threonine, valine and histidine and while the non-essential amino acids were alanine, arginine, glutamic acid, glycine, serine and tyrosine, respectively. The nonessential amino acids play a more important part than the essential ones in the regulation of cellular osmotic pressure as investigated in the tissues of Callinectes sapidus (Gerard and Gilles, 1972). The results obtained with the male and female, in that the lowest free amino acid concentration is found in the female, while the male have the highest concentrations.
Moreover, individual amino acids show considerable variation in proportions from male to female. To be precise, males tended to have higher free amino acid content in the hemolymph than the females as similar to fresh water field crab, Paratelphusa sp. (Padmanabhanaidu and Ramamurthy, 1961). The total concentration of amino acids found in the hemolymph of male and female (41.3 μM mL-1, male; 27.5 μM mL-1, female) was lower than that in the muscle (570 μM mL-1, male; 76.3 μM mL-1, female) as similar to Maia squinado, Eriocheir sinensis, Callinectes sapidus and Penaeus monodon (Camien et al., 1951; Gerard and Gilles, 1972; Fang et al., 1992). The pattern of blood amino acids might play a significant role in keeping the osmotic pressure of males at a higher level than that of females (Padmanabhanaidu and Ramamurthy, 1961). Gerard and Gilles (1972) reported that the level of free intracellular amino acids of various species varies with changes in the osmotic pressure of the surrounding fluid.
In our study, arginine was found in both sexes. It probably resulted from the hydrolysis of arginine phosphate, a well-known phosphagen in invertebrates (Simpson et al., 1959). It can be the function of arginine, phosphate, which takes over the role of creatine phosphate in muscle contraction in crustacean (Meister, 1965). The post-ecdysial reduction in glycine, aspartic and glutamic acids (which are the three predominant constituents of cuticle) may be attributed to cuticular deposition (Welinder, 1974). Glycine, arginine, glutamic acid and alanine were the amino acids found higher in males than females, these are the important substances for the regulation of osmotic pressure in Crustacean muscles (Camien et al., 1951).
It may be inferred, therefore, the total osmotic pressure of male crabs is higher than that of females at any given body weight (Gilbert, 1 959a-c). Glutamic acid exhibited the both sexes, reemphasizing its central role in amino acid metabolism (Wheatly, 1985). Moreover, free amino acids such as alanine, glutamic acid and glycine, are responsible for flavour and taste. Alanine and glycine have sweet tastes and glutamic acid has the umami taste typical of crustaceans (Yamanaka and Shimada, 1996). The very smaller concentration of aspartic acid was noticed and would be expected these amino acids were present largely either as their amides or as peptides (Camien et al., 1951).
Further, It may be found in the nerve might, therefore, be explained in the light of its possible role in the cation-anion balance of the tissue (Gerard and Gilles, 1972). The less concentration of alanine was also observed in our study. These three amino acids (Glu, Asp and Ala) are closely linked to the citric acid cycle and their concentration could be easily regulated by the reactions of the cycle (Simpson et al., 1959). Keto precursors of the dicarboxylic amino acids (glutamic acid and aspartic acid) at ecdysis may be synthesized from 3C fragments of glycolysis if intermediary metabolism becomes geared towards more anaerobic metabolism (Schoffeniels and Gilles, 1970b). Alternatively, there may be a second metabolic shunt of the Krebs cycle leading to their formation which is unique to crustaceans (Schoffeniels and Gilles, 1970a).
The minor contributions of valine and phenylalanine were found only in male. The reduction in nonpolar species (valine and phenylalanine) can subsequently be correlated with hardening which occurs in postmolt stage B (Stevenson, 1972). Several investigators (Camien et al., 1951; Stevens et al., 1961; Thompson and Farragut, 1966; Evans, 1972; Gerard and Gilles, 1972; Wheatly, 1985; Naczk et al., 2004; Kaya et al., 2009) detected only small amount of histidine in different species of crab, in our experiments histidine was shown to be the most abundant free amino acid. Free histidine has been proposed to be involved in a mechanism for zinc transport (Baslow, 1965, 1966, 1967; Baslow and Lenney, 1967; Erspamer et al., 1965). Some amino acids (tryptophan, asparagine and glutamine) were destroyed by acid hydrolysis and were not determined as investigated by Martinez et al. (2007). This may also be a strong reason that these amino acids were not exhibited in our study.
ACKNOWLEDGMENT
We express our sincere gratitude to Dr. N. Angayarkanni, for her help extended in analysing the amino acid profile at Department of Biochemistry and Cell Biology, Sankara Nethralaya.
REFERENCES
- Anaya, J.P. and S. D'Aniello, 2006. Free amino acids in the nervous system of the amphioxus Branchiostoma lanceolatum. Int. J. Biol. Sci., 2: 87-92.
CrossRef - Baslow, M.H., 1967. N-acetyl-L-histidine metabolism in the fish eye: Evidence for ocular fluid lens L-histidine recycling. Exp. Eye Res., 6: 336-342.
PubMed - Bursell, E., 1966. Aspects of the flight metabolism of tsetse flies (Glossina). Comp. Biochem. Physiol., 19: 809-818.
CrossRef - Camien, M.N., H. Sarlet, G. Duchateau and M. Florkin, 1951. Non-protein amino acids in muscle and blood of marine and fresh-water Crustacea. J. Biol. Chem., 193: 881-885.
Direct Link - Capillas, C.R., 2004. Free amino acids in muscle of Norway lobster Nephrops novergicus (L.) in controlled and modified atmospheres during chilled storage. Food Chem., 86: 85-91.
CrossRef - Duchateau, G. and M. Florkin, 1958. A survey of aminoacidemias with special reference to the high concentration of free amino acids in insect hemolymph. Arch. Intern. Physiol Biochem., 66: 573-591.
PubMed - Erspamer, V., M. Roseghini and A. Anastasi, 1965. Occurrence and distribution of N-acetylhistidine in brain and extra cerebral tissues of poikilothermal vertebrates. J. Neurochem., 12: 123-130.
CrossRef - Evans, P.D., 1972. The free amino acid pool of the haemocytes of Carcinus maenas (L.). J. Exp. Biol., 56: 501-507.
Direct Link - Findley, A.M and W.B. Stichle, 1978. Effects of salinity fluctuation on the hemolymph composition of the blue crab Callinectes sapidus. Mar. Biol., 46: 9-15.
CrossRef - Gilbert, A.B., 1959. The composition of blood of the shore crab Carcinus moenas Pennant in relation to sex and size. I. Blood conductivity and freezing point depressions. J. Exp. Biol., 36: 113-119.
Direct Link - Kaya, Y., H. Turan and M.E. Erdem, 2009. Determination of nutritional quality of warty crab (Eriphia verrucosa forsskal, 1775). J. Anim. Vet. Adv., 8: 120-124.
Direct Link - Martinez, M.V., J. L. Hernandez and M.A. Lage-Yusty, 2007. Protein and amino acid contents in the crab, Chionoecetes opilio. Food Chem., 103: 1330-1336.
CrossRef - McCoid, V., R. Miget and G. Finne, 1984. Effect of environmental salinity of the free amino acid composition and concentration in penaeid shrimp. J. Food. Sci., 49: 327-330.
CrossRef - Naczk, M., J. Williams, K. Brennan, C. Liyanapathirana and F. Shahidi, 2004. Compositional characteristics of green crab (Carcinus maenas). Food Chem., 88: 429-434.
CrossRefDirect Link - Shewbart, K.L., W.L. Mies and P.D. Ludwig, 1972. Identification and quantitative analysis of the amino acids present in protein of the brown shrimp Penaeus aztecus. Mar. Biol., 16: 64-67.
CrossRefDirect Link - Simpson, J.W., K. Allen and J. Awapara, 1959. Free amino acids in some aquatic invertebrates. Biol. Bull., 117: 371-381.
Direct Link - Skinner, D.M., 1965. Amino acid incorporation into protein during the molt cycle of the land crab, Gecarcinus lateralis. J. Exp. Zool., 160: 225-234.
CrossRefDirect Link - Skinner, D.M., 1966. Breakdown and reformation of somatic muscle during molt cycle of the land crab, Gecarcinus lateralis. J. Exp. Zool., 163: 115-124.
CrossRefDirect Link - Stevens, T.M., C.E. Howard and R.W. Schlesinger, 1961. Free amino acids in sera of the marine invertebrates, Cancer Irroratus, Limulus polyphemus and Homarus Americanus. Comp. Biochem. Physiol., 3: 310-314.
PubMed - Stevenson, J.R., 1972. Changing activities of the crustacean epidermis during the molting cycle. Am. Zool., 12: 373-379.
CrossRefDirect Link - Thompson, M.H. and R.N. Farragut, 1982. Amino acid composition of the Chesapeaeke bay blue crab Callinectes sapidus. Comp. Biochem. Physiol., 17: 1065-1078.
CrossRefDirect Link - Van Waarde, A., 1988. Biochemistry of non-protein nitrogenous compounds in fish including the use of amino acids for anaerobic energy production. Comp. Biochem. Physiol., 91: 207-228.
CrossRefDirect Link - Weber, R.E. and W.J.A. van Marrewijk, 1972. Free amino acids and isosmotic intracellular regulation in the shrimp Crangon crangon. Life Sci., 11: 589-595.
Direct Link - Weber, R.E. and W.J.A. van Marrewijk, 1972. Free amino acids in the shrimp Crangon crangon and their osmoregulatory significance. Neth. J. Sea. Res., 5: 391-415.
CrossRefDirect Link - Welinder, B.S., 1974. The crustacean cuticle. I. Studies on the composition of the cuticle. Comp. Biochem. Physiol., 47: 779-787.
CrossRefDirect Link - Welinder, B.S., 1975. The crustacean cuticle. II. Deposition of organic and inorganic material in the cuticle of Astacus fluviatilis in the period after moulting. Comp. Biochem. Physiol., 51: 409-416.
CrossRefDirect Link - Wheatly, M.G., 1985. Free amino acid and inorganic ion regulation in the whole muscle and hemolymph of the blue crab Callinectes sapidus rathbun in relation to the molting cycle. J. Crust. Biol., 5: 223-233.
Direct Link - Van Marrewijk, W.J.A. and H.J.L. Ravestein, 1974. Amino acid metabolism of Astacus leptodactylus esch-I. Composition of the free and protein-bound amino acids in different organs of the crayfish. Comp. Biochem. Physiol., 47: 531-542.
CrossRefDirect Link - Yamanaka, H. and R. Shimada, 1996. Post mortem biochemical changes in the muscle of Japanese spiny lobster during storage. Fish. Sci., 62: 821-824.
Direct Link