
Mahima
Animal Nutrition, U.P. Pt. Deen Dayal Upadhayay Pashu Chikitsa Vigyan Vishvidyalaya Evum Go-Anusandhan Sansthan (DUVASU), Mathura, India
A.K. Garg
Department of Centre of Advanced Studies in Animal Nutrition, Indian Veterinary Research Institute, Izatnagar, India
Vishal Mudgal
Department of Animal Nutrition, Veterinary College, Jabalpur, Madhya Pradesh, India
Pakistan Journal of Biological Sciences
Year: 2012 | Volume: 15 | Issue: 9 | Page No.: 448-453







ABSTRACT
A 70 day experiment on forty guinea pigs (Cavia porcellus) was conducted to find the influence of different level of sodium selenite (inorganic selenium supplementation) on growth, nutrient utilization and selenium uptake. The sodium selenite was supplemented into a basal diet at 0, 0.1, 0.2 and 0.3 ppm, respectively and the basal diet comprised of 25% ground cowpea (Vigna unguiculata) hay, 30% ground maize (Zea mays) grain, 22% ground gram (Cicer arietinum) grain, 9.5% deoiled rice (Oryza sativa) bran, 6% soybean (Glycine max) meal, 6% fish meal, 1.5% mineral mixture (without Se), ascorbic acid (200 mg kg-1) and 0.1 ppm Se to meet their nutrient requirements. Daily feed intake and weekly body weights were recorded. Intake and digestibility of dry matter, organic matter, ether extract, crude fiber and nitrogen-free extract as well as uptake of calcium and phosphorus, total body weight and average daily gain were similar (p>0.05) among the four groups. However, there was a trend of increase in Se absorption of the guinea pigs with the increasing levels of Se, in the groups given 0.2 and 0.3 ppm of Se. It can be concluded that requirement of Se in guinea pigs is 0.1 ppm, as supplementation of ≥0.1 ppm sodium selenite in the diet (having 0.1 ppm Se) did not enhanced their growth rate and nutrient utilization.
PDF Abstract XML References Citation
Received: May 11, 2012;
Accepted: August 06, 2012;
Published: September 07, 2012
How to cite this article
Mahima, A.K. Garg and Vishal Mudgal, 2012. Influence of Sodium Selenite on Growth, Nutrient Utilization and Selenium Uptake in Cavia porcellus. Pakistan Journal of Biological Sciences, 15: 448-453.
DOI: 10.3923/pjbs.2012.448.453
URL: https://scialert.net/abstract/?doi=pjbs.2012.448.453
DOI: 10.3923/pjbs.2012.448.453
URL: https://scialert.net/abstract/?doi=pjbs.2012.448.453
INTRODUCTION
Recently, there has been a growing understanding that marginal trace element deficiency is one of the major factor limiting health and productivity of the animals. This is partly because of important physiological role of most of the trace elements in the body’s disease resistance mechanisms. Along with the quantitative aspects it has also been realized that mineral form is also critical for its function and utilization in the body systems. Since, more than half a century, Selenium (Se) was considered as an essential trace mineral for animals viz., rat (Schwarz and Foltz, 1957) and birds (Patterson et al., 1957) and play important role in various biochemical functions in human as ell as animals like hepatoprotective effect (Ghazi-Khansari et al., 2005; Rahimi et al., 2012), antioxidant defense (Sharma, 2007; Fan et al., 2009; Ozdemir, 2011), immune function, reproduction protein turnover, growth performance of the animals via thyroid metabolism (Jianhua et al., 2000; Sharadamma et al., 2011) as it is a component of enzyme type I deiodinase which is required for the conversion of thyroxine in to more active triiodothyronine (Beckett et al., 1987). It is an important component of antioxidant enzymes such as glutathione peroxidase and thioredoxin reductase, thus regulating the body’s glutathione pool (Tapiero et al., 2003).
At present, the recommended requirements of Se (0.15 ppm), for guinea pigs (NRC, 1995) were based on the requirement established for the rats, with the suggestion that they can be used till the requirements for guinea pigs are established. However, in a study, the requirement of Se for guinea pigs was recommended as 0.08 ppm (Jensen and Pallauf, 2007). Scanty of information appear to be available in the literature regarding the effect of selenium supplementation (as sodium selenite) on growth rate, feed intake and utilization of nutrients in guinea pigs. In view of these facts, the present study was conducted with the objective to evaluate the effect of supplementation of selenium through inorganic (sodium selenite) sources at different levels i.e., 0.1, 0.2 and 0.3 ppm on the performance of guinea pigs, particularly on their growth rate, nutrient utilization and feed conversion efficiency.
MATERIALS AND METHODS
General: Animal grouping, housing, feeding and management: Forty healthy weaned domestic Guinea pigs (Cavia porcellus) with 144.3±2.04 g mean body weight were divided into four equal groups of ten animals each following completely randomized design. The basal diet was formulated to meet the nutrient requirement as per NRC (1995) (Table 1). Guinea pigs allotted to group I (control) were fed only the basal diet, guinea pigs allotted to groups II, III and IV were additionally supplemented with 0.1, 0.2 and 0.3 ppm of Se as sodium selenite, respectively. Ad libitum water and about 10-20 g of available green fodder (berseem) was offered daily to each animal to meet its vitamin A requirements. Experimental feeding lasted for a period of 70 days including a three day digestibility trial. To find out the daily feed (DM) intake, a weighed amount of the basal diet was offered daily at 10:00 a.m. and residue left after 24 h was collected and quantified.
Body weight gain, digestibility trial and sampling and chemical analysis: Animals were weighed at weekly interval in the morning before offering feed and water throughout the experimental period to assess their growth rate. To find out the influence of Se supplementation on intake and utilization of different nutrients, a 3 day digestibility trial was conducted after 21 days of experimental feeding on six animals randomly selected from each group and they were kept and fed individually in plastic cages having the facility for individual feeding and watering as well as faeces collection. During the digestion trial, amount of feed offered, residue left and faeces voided in 24 h were daily quantified and recorded for all the animals separately and a representative sample of each was dried in a hot air oven at 100±1°C overnight and preserved (Schneider and Flat, 1975) for further analysis. A suitable aliquot of feces was also daily preserved in 20% sulfuric acid for the estimation of nitrogen/Crude Protein (CP). The pooled dried samples were ground in a Willey mill to pass through a 1 mm sieve and analyzed for proximate principles and phosphorous (AOAC, 2000) and calcium (Ca) (Talapatra et al., 1940). Se contents in these samples were estimated from their mineral extracts with the help of atomic absorption spectrophotometer (Model 4141, Electronic Corporation of India Limited, Hydrabad, India) using air acetylene flame.
Statistical analysis: Data generated in the study were statistically analyzed using analysis of variance (ANOVA) technique (Snedecor and Cochran, 1989) and means were compared using Duncan’s multiple-range test (Steel and Torrie, 1980).
RESULTS AND DISCUSSION
The chemical composition of the basal diet offered to the animals in various groups was having organic matter (88.61%), crude protein (22.41%), ether extract (3.93%), total ash (11.39%), crude fibre (11.23%), NFE (51.04%), calcium (1.63%), P (0.54%) and Se (0.1 ppm) on dry matter basis Table 1 which were as per National Research Council recommendations (NRC, 1995). The Se analyses of our diets (Table 2) indicated that the basal diet contained 0.1 ppm of Se. So, the group I (control) was having Se 0.1 ppm while the group II, III and IV were having Se as 0.2, 0.3 and 0.4 ppm, respectively.
From the Table 3, it can be seen that there was no significant difference in DM intake among the groups, indicating that Se supplementation as sodium selenite up to 0.4 ppm level in the basal diet (containing 0.1 ppm Se) had no adverse effect on the palatability and feed intake pattern of the animals. Similar to our findings, no adverse effect on feed intake was observed in hamsters (Julius et al., 1983; Birt et al., 1986), poultry (Echevarria et al., 1988; Payne and Southern, 2005a, b), gilts (Mahan and Peters, 2004), piglets (Daza et al., 2000), sheep (White and Somers, 1977), buffalo calves (Mudgal, 2005), lambs (Kumar, 2006) with 0.1- 0.5 ppm Se (as sodium selenite) in basal diet. Contrary to our findings, here are also few studies where supplementation of Se has adversely affected the feed intake of the animals, as was observed in chicks supplemented with 9 mg Se kg-1 diet (Echevarra i et al., 1988).
| Table 1: | Chemical composition of the basal diet offered to guinea pigs |
 | |
| Table 2: | Comparison of supplemental with actual analyzed selenium levels in diets* |
 | |
| *Analyzed Se levels in diets are the total Se levels reported as basal (unsupplemented) level plus supplemental level | |
| Table 3: | Intake and digestibility of nutrients and plane of nutrition in different groups |
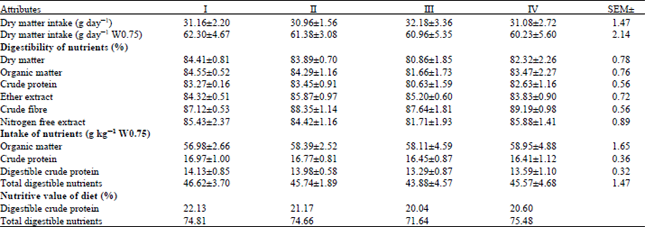 | |
But in that experiment, the level of Se supplementation was too high which may have caused an adverse effect on feed intake of the animals.
DM digestibility among the four groups were found as 84.41, 83.89, 80.86 and 82.32% in groups I, II, III and IV, respectively Table 3 which were statistically non-significant (p>0.05). Similar to our findings, no effect on DM digestibility was reported in merino weathers (White and Somers, 1977), sheep (Nicholson et al., 1991), buffalo calves (Mudgal, 2005) and lambs (Kumar, 2006), on supplementation of Se as sodium selenite. However, contrary to our findings, an improvement in DM digestibility was reported by supplementation of 0.025, 0.050, 0.075 and 0.100 ppm Se (Adkins and Ewan, 1984) and 0.05 ppm Se (Glienke and Ewan, 1977) in pigs. But, the probable reason for this improvement may be due to very low levels of Se in the basal diets (Se deficient diet) used by these workers. Similarly, the digestibility of Organic Matter (OM) was also comparable (p>0.05) among the four groups which is in agreement with the earlier reports in sheep (Nicholson et al., 1991; Kumar, 2006) and buffaloes (Mudgal, 2005; Mudgal et al., 2008). There was no significant difference (p>0.05) in the digestibility of Crude Protein (CP) among the four groups which is in accordance to the earlier studies in male Holstein calves (Nicholson et al., 1991) and buffalo calves (Mudgal et al., 2008). It was further observed that the digestibility of EE, Crude Fiber (CF) and nitrogen-free extract (NFE) was also comparable (p>0.05) among the four groups (Table 3). Similar to our findings, there was no effect on digestibility of EE and different fiber fractions (neutral detergent fiber, acid detergent fiber, hemicellulose and cellulose) in growing lambs (Kumar, 2006) and male buffalo calves (Mudgal et al., 2008) supplemented with 0.15 and 0.3 ppm Se, respectively. In contrast to our observations (Glienke and Ewan, 1977) found an improved digestibility of EE in 0.05 ppm Se supplemented pigs. But, probably it was due to Se deficient basal diet used by these workers, because on supplementation of 0.1 ppm Se, there was no further improvement in the EE digestibility over 0.05 ppm group. However, the basal diet used in our study was having 0.1 ppm Se which might have been sufficient for optimum utilization of EE, CF and NFE and therefore, further supplementation of Se in the diet had no effect on the digestibility of these nutrients in our study. There were no significant difference (p>0.05) in the intake of Organic Matter (OM), Crude Protein (CP), Digestible Crude Protein (DCP), Total Digestible Nutrients (TDN), nutritive value (TDN and DCP) of diet among the different groups (Table 3). The similar findings were reported in earlier studies in which supplementation of 0.15 ppm (Kumar, 2006) and 0.3 ppm (Mudgal et al., 2008) Se had no effect on intake of OM, CP, TDN and DCP and nutritive (TDN and DCP) value of the diets in the lambs and buffalo calves, respectively. It indicated that supplementation of selenium as sodium selenite up to 0.4 ppm level has no adverse effect on plane of nutrition and nutritive values of diets in the guinea pigs.
The daily intake, amount absorbed and absorption coefficient of Ca and P were similar (p>0.05) among the four groups (Table 4) which were in agreement with Kumar (2006) and Mudgal et al. (2008), who also reported no effect on Ca and P metabolism with supplementation of 0.15 and 0.3 ppm Se in the lambs and buffalo calves, respectively. The results indicated that supplementation of Se up to 0.4 ppm level has no effect on Ca and P metabolism in the guinea pigs.
The daily Se intake among the four groups were found as 2.96, 6.14, 9.2 and 12.77 μg in groups I, II, III and IV, respectively (Table 4). The daily Se intake was significantly (p<0.05) increasing with the increase in Se supplementation in diet which was obviously due to increasing levels of Se supplementation in these groups.
| Table 4: | Intake and adsorption of Ca and P and Se in different groups |
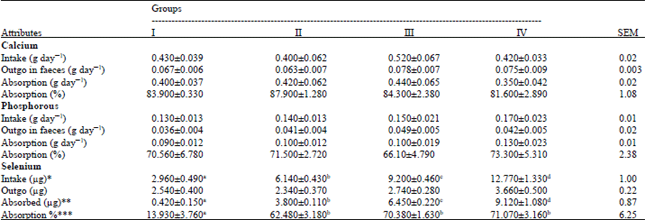 | |
| Means followed by different letters in a row differ significantly at *p<0.05, **p<0.01, ***p<0.001 | |
| Table 5: | Growth rate and feed conversion efficiency in different groups |
 | |
It was further observed that despite of significantly higher intake of Se in groups II, III and IV, excretion of Se through feces was similar (p>0.05) among the four groups, with the result, the amount of Se absorbed and absorption coefficient were highest (p<0.05) in group IV, followed by group III and group II and lowest in group I (control). Similar to our observations, at low levels of Se supplementation, apparent Se digestibility was found to increase with the increasing levels of dietary Se supplementation from 0, 0.25, 1.0 and 2.0 ppm in growing lambs (McClure and Mahan, 1988) and with 5 ppm Se than with 0.3 ppm Se diets in pigs (Lowry et al., 1985). Contrarily, Se concentration increased significantly in feces in camels given an oral supplementation of 0, 2 and 4 mg Se for 3 months but there was no difference between groups receiving 2 and 4 mg Se (Seboussi et al., 2008). However, it may be due to very high levels of Se supplementation, as compared to our experiment as ADG 2.88±0.24, 2.72±0.23, 3.00±0.17 and 2.99±0.21 g day-1.
The weekly body weights and growth performance of the guinea pigs in different groups have been shown in Table 5. The Average Daily Gain (ADG) among the four groups were found as 2.88, 2.72, 3.00 and 2.99 g day-1 in groups I, II, III and IV, respectively. The total body weight, Average Daily Gain (ADG) were similar (p>0.05), among the four groups. Similar to our results, no effect was observed on body weights of broilers fed 0.1 and 0.25 ppm of organic and inorganic Se (Choct et al., 2004). Similarly, there was no difference in body weight gains in broilers fed diets containing 0, 0.05 and 0.15 ppm Se from sodium selenite or selenomethionine (Spears et al., 2003), in ducks with 0.1 and 1.0 ppm Se supplementation (Dean and Combs Jr., 1981) and in lambs supplemented with 0.3 and 0.45 ppm Se (Vignola et al., 2009).
CONCLUSION
It can be concluded that requirement of Se in guinea pigs is 0.1 ppm, as supplementation of ≥0.1 ppm sodium selenite in the diet (having 0.1 ppm Se) did not enhanced their growth rate and nutrient utilization. However, supplementation of Se up to 4 ppm had no adverse effect on palatability, feed intake, plane of nutrition, nutritive value of diet, Ca and P metabolism, body weight and average daily gain in guinea pigs.
REFERENCES
- Adkins, R.S. and R.C. Ewan, 1984. Effect of supplemental selenium on pancreatic function and nutrient digestibility in the pig. J. Anim. Sci., 58: 351-355.
PubMedDirect Link - Julius, A.D., M.H. Davies and D.F. Birt, 1983. Toxic effects of dietary selenium in syrian hamster. Ann. Nutr. Metab., 27: 296-305.
PubMedDirect Link - Beckett, G.J., S.E. Beddows, P.C. Morrice, F. Nicol and J.R. Arthur, 1987. Inhibition of hepatic deiodination of thyroxine is caused by selenium deficiency in rats. Biochem. J., 248: 443-447.
PubMedDirect Link - Choct, M., A.J. Naylor and N. Reinke, 2004. Selenium supplementation affects broiler growth performance, meat yield and feather coverage. Br. Poult. Sci., 45: 677-683.
CrossRefPubMedDirect Link - Daza, A., S. Salado, J.F. Galvez and M. Gutierrez-Barquin, 2000. Effects of vitamin E and selenium supplementation on the immune system, hematological parameters and the performance of weaned pigs. Invest. Agaria Produccion-y-Sanidad-Anim., 15: 21-30.
Direct Link - Dean, W.F. and G.F. Combs Jr., 1981. Influence of dietary selenium on performance, tissue selenium content and plasma concentrations of selenium-dependent glutathione peroxidase, vitamin E and ascorbic acid in ducklings. Poult. Sci., 60: 2655-2663.
PubMedDirect Link - Birt, D.F., A.D. Julius and C.E. Runice, 1986. Tolerance of low and high dietary selenium throughout the life span of Syrian hamsters. Ann. Nutr. Metab., 30: 233-240.
CrossRefDirect Link - Echevarria, M.G., P.R. Henry, C.B. Ammerman, P.V. Rao and R.D. Miles, 1988. Estimation of the relative bioavailability of inorganic selenium sources for poultry. 2. Tissue uptake of selenium from high dietary selenium concentrations. Poult. Sci., 67: 1585-1592.
PubMedDirect Link - Glienke, L.R. and R.C. Ewan, 1977. Selenium deficiency in the young pig. J. Anim. Sci., 45: 1334-1340.
Direct Link - Jensen, C. and J. Pallauf, 2007. Estimation of the selenium requirement of growing guinea pigs (Cavia porcellus). J. Anim. Physiol. Anim. Nutr., 92: 481-491.
CrossRef - Jianhua, H., A. Ohtsuka and K. Hayashi, 2000. Selenium influences growth via thyroid hormone status in broiler chickens. Br. J. Nutr., 84: 727-732.
Direct Link - Lowry, K.R., D.C. Mahan and J.R. Corley, 1985. Effect of dietary calcium on selenium retention in postweaning swine. J. Anim. Sci., 60: 1429-1437.
PubMedDirect Link - Mahan, D.C. and J.C. Peters, 2004. Long-term effects of dietary organic and inorganic selenium sources and levels on reproducing sows and their progeny. J. Anim. Sci., 82: 1343-1358.
Direct Link - McClure, K.E. and D.C. Mahan, 1988. Effect of dietary selenium source of retention, digestibility and wool accumulation of selenium in growing lambs. Nutr. Rep. Int., 37: 839-846.
Direct Link - Mudgal, V., A.K. Garg, R.S. Dass and V.P. Varshney, 2008. Effect of selenium and copper supplementation on blood metabolic profile in male buffalo (Bubalus bubalis) calves. Biol. Trace Elem. Res., 121: 31-38.
CrossRef - Nicholson, J.W.G., R.E. McQueen and R.S. Bush, 1991. Response of growing cattle to supplementation with organically bound or inorganic sources of selenium or yeast cultures. Can. J. Anim. Sci., 71: 803-811.
CrossRefDirect Link - Payne, R.L. and L.L. Southern, 2005. Comparison of inorganic and organic selenium sources for broilers. Poult. Sci., 84: 898-902.
CrossRefDirect Link - Payne, R.L. and L.L. Southern, 2005. Changes in glutathione peroxidase and tissue selenium concentrations of broilers after consuming a diet adequate in selenium. Poult. Sci., 84: 1268-1276.
Direct Link - Schwarz, K. and C.M. Foltz, 1957. Selenium as integral part of factor-3 against dietary necrotic liver degeneration. J. Am. Chem. Soc., 79: 3292-3293.
Direct Link - Seboussi, R., B. Faye, G. Alhadrami, M. Askar, W. Ibrahim, K. Hassan and B. Mahjoub, 2008. Effect of different selenium supplementation levels on selenium status in camel. Biol. Trace Elem. Res., 123: 124-138.
PubMed - Snedecor, G.W. and W.G. Cochran, 1989. Statistical Methods. 8th Edn., Iowa State University Press, Ames, Iowa, ISBN-13: 9780813815619 Pages: 524.
Direct Link - Talapatra, S.K., S.C. Roy and K.C. Sen, 1940. The analysis of mineral constituents in biological materials. 1. Estimation of phosphorus, chlorine, calcium, magnesium, sodium and potassium in food-stuffs. Indian J. Vet. Sci., 10: 243-258.
Direct Link - Vignola, G., L. Lambertini, G. Mazzone, M. Giammarco, M. Tassinari, G. Martelli and G. Bertin, 2009. Effects of selenium source and level of supplementation on the performance and meat quality of lambs. Meat Sci., 81: 678-685.
PubMed - White, C.L. and M. Somers, 1977. Sulphur-selenium studies in sheep I. The effect of varying dietary sulphate and selenomethionine on sulphur, nitrogen and selenium metabolism in sheep. Aust. J. Biol. Sci., 30: 47-56.
Direct Link - Fan, C., B. Yu and D. Chen, 2009. Effects of different sources and levels of selenium on performance, thyroid function and antioxidant status in stressed broiler chickens. Int. J. Poult. Sci., 8: 583-587.
CrossRefDirect Link - Ozdemir, E., 2011. Physiopathological role of selenium and selenoprotein in neuropsychiatric disease. J. Med. Sci., 11: 11-18.
CrossRefDirect Link - Ghazi-Khansari, M., M. Karami, M. Rezayat, M. Abdollahi, B. Minaie and O. Sabzevari, 2005. The protective effects of antioxidants and propranolol on hepatotoxicity of TCDD during isolated rat liver perfusion. Int. J. Pharmacol., 1: 336-341.
CrossRefDirect Link - Rahimi, H.R., M. Gholami, H.R. Khorram-Khorshid, F. Gharibdoost and M. Abdollahi, 2012. On the protective effects of IMOD and silymarin combination in a rat model of acute hepatic failure through anti oxidative stress mechanisms. Asian. J. Anim. Vet. Adv., 7: 38-45.
CrossRefDirect Link - Sharadamma, K.C., B. Purushotham, P.M. Radhakrishna, P.M. Abhilekha and H.M. Vagdevi, 2011. Role of selenium in pets health and nutrition: A review. Asian J. Animal Sci., 5: 64-70.
CrossRefDirect Link - Sharma, N., 2007. Alternative approach to control intramammary infection in dairy cows: A review. Asian J. Anim. Vet. Adv., 2: 50-62.
CrossRefDirect Link - Tapiero, H., D.M. Townsend and K.D. Tew, 2003. The antioxidant role of selenium and seleno-compounds. Biomed. Pharmacother., 57: 134-144.
Direct Link