
Mohammed Z. Allouh
Department of Anatomy, Faculty of Medicine, Jordan University of Science and Technology, Irbid, Jordan
Mohammed H. Aldirawi
Department of Anatomy, Faculty of Medicine, Jordan University of Science and Technology, Irbid, Jordan
Pakistan Journal of Biological Sciences
Year: 2012 | Volume: 15 | Issue: 5 | Page No.: 215-223







ABSTRACT
Sustanon is one of the most commonly used anabolic androgenic drugs to increase skeletal muscle mass and strength. This drug is a blend of four esterized testosterone derivatives: Testosterone propionate, testosterone phenylpropionate, testosterone isocaproate and testosterone decanoate. Little is known about the effects of this drug on skeletal muscle at the cellular level. This study aimed to investigate the influence of Sustanon on the morphology of skeletal muscle fibers and the distribution of myogenic stem cells known as Satellite Cells (SCs) during postnatal growth. We hypothesized that Sustanon-induced skeletal muscle hypertrophy is associated with an increase in the number of SCs. Robust immunocytochemical techniques and morphometric analyses were used to calculate the numbers of SCs and myonuclei within the pectoralis muscle of chickens. Also, DNA concentration and Pax7 protein levels were measured to confirm immunocytochemical findings. Sustanon significantly increased pectoralis mass and fiber size. All SC indices and the number of myonuclei increased significantly by Sustanon administration. In addition, greater DNA concentration and Pax7 protein expression were found in Sustanon-treated birds. This study indicates that Sustanon can induce avian skeletal muscle hypertrophy and that this is correlated with increased numbers of SCs and myonuclei.
PDF Abstract XML References Citation
Received: February 19, 2012;
Accepted: May 26, 2012;
Published: June 29, 2012
How to cite this article
Mohammed Z. Allouh and Mohammed H. Aldirawi, 2012. Effects of Sustanon on the Distribution of Satellite Cells and The Morphology of Skeletal Muscle Fibers During Maturation. Pakistan Journal of Biological Sciences, 15: 215-223.
DOI: 10.3923/pjbs.2012.215.223
URL: https://scialert.net/abstract/?doi=pjbs.2012.215.223
DOI: 10.3923/pjbs.2012.215.223
URL: https://scialert.net/abstract/?doi=pjbs.2012.215.223
INTRODUCTION
Satellite cells (SCs) are myogenic stem cells that reside between the basal lamina and plasmalemma of the skeletal muscle fiber (Pophal et al., 2004; Moore et al., 2005; Zammit et al., 2006; Allouh et al., 2008). They are responsible for postnatal muscle growth, repair and regeneration (Zammit et al., 2006; Mohamed et al., 2007). Normally, SCs are quiescent in the skeletal muscle; however, when growth or repair is necessary, SCs become active and start to proliferate (Hawke and Garry, 2001; Morgan and Partridge, 2003). Eventually, they differentiate by fusing to muscle fibers, where their nuclei become new myonuclei (Hawke and Garry, 2001; Pophal et al., 2004; Moore et al., 2005). In a less common scenario, SCs may fuse together to form a new myofiber (Hawke and Garry, 2001; Zammit et al., 2006). Some daughter cells return to quiescence to maintain a reservoir of SCs (Dhawan and Rando, 2005; Wozniak et al., 2005).
SCs express many molecular markers that can be used in their identification, such as paired box transcription factor 7 (Pax7), M-cadherin, c-met and CD34 (Wozniak et al., 2005; Zammit et al., 2006). Pax7 is particularly useful for this purpose because it is expressed by SCs during each of the different stages of quiescence, proliferation and differentiation but is not expressed by myonuclei. In a previous study, we reported specific expression of Pax7 by all resident SCs in the chicken pectoralis muscle (Allouh et al., 2008).
Anabolic Androgenic Steroids (AASs) are synthetic derivatives of the endogenous primarily male steroid hormone, testosterone. It is already established that AASs increase skeletal muscle mass and strength (Bhasin et al., 2001; Pala, 2003; Marjani et al., 2009) and enhance athletic performance (Ebrahimi, 2009). However, the exact mechanisms by which steroids increase lean body mass are still poorly understood. One of the expected mechanisms by which AASs exert their effects on the muscle might be through activation of SCs, since proliferation and differentiation of SCs into new myonuclei is a fundamental process for muscle fiber growth (O’Connor and Pavlath, 2007; Velleman et al., 2010). In fact, Sinha-Hikim et al. (2004) have shown that both myonuclei and Satellite Cell Nuclei (SCN) express androgen receptors.
Sustanon is a popular and commonly used anabolic drug. Its popularity is likely due to the notion that it exhibits superior anabolic effects when compared to other testosterone compounds (Beotra, 2005; Wills, 2005). This drug is a mixture of four testosterone derivatives that have been found to react very positively together. The mixture includes: testosterone propionate, testosterone phenylpropionate, testosterone isocaproate and testosterone decanoate. The effect of these components is time-released, so that Sustanon goes rapidly into the system and remains effective in the body for several weeks (Ahmed Bin Bisher, 2009). Sustanon is effective after 1 day of administration due to the testosterone propionate component (Fitts et al., 2004; Beg et al., 2008) but remains active for 3-4 weeks due to the testosterone decanoate component (Anderson et al., 2002). Sustanon is clinically used to treat many cases of osteoporosis, male hypogonadism, infertility and absence of male libido (Ahmed Bin Bisher, 2009).
Based on the previous facts, this study aimed to examine the effects of Sustanon on the distribution of SCs and the morphology of skeletal muscle fibers during maturation. It tested the hypothesis that Sustanon-induced skeletal muscle hypertrophy is associated with an increase in the number of SCs. Applying immunocytochemical techniques and computer image analyses, the frequency of SCs, number of myonuclei and size of muscle fibers were determined in both control and Sustanon-treated groups. Sustanon significantly increased pectoralis mass and fiber size. In addition, all SC indices and the number of myonuclei increased significantly by Sustanon administration. This study indicates that Sustanon can induce avian skeletal muscle hypertrophy and that this is correlated with increased numbers of SCs and myonuclei.
MATERIALS AND METHODS
Animals: Female broiler chickens (Gallus gallus) were hatched at the same time and raised under proper conditions in the animal care unit at Jordan University of Science and Technology (JUST). All animal care procedures and treatments were conducted with the approval of the JUST committee on animal care and in accordance with the guidelines of the U.S. National Institutes of Health on the use and care of laboratory animals. The birds were fed ad libitum. Initial room temperature was 35°C. Subsequently, temperature was decreased 2.5°C at weekly intervals until 21°C was reached when the birds were 6 weeks old. The birds were separated into two groups (control and Sustanon-treated). Each group contained five birds. At the age of 6 weeks, the injections into the right pectoralis muscle of the birds were begun. The Sustanon-treated group received four injections at weekly intervals (1 injection/week). Each injection contained 30 mg of Sustanon (Organon Int., Oss, The Netherlands). The control group received normal saline injections. Birds were sacrificed by cervical dislocation at the age of 10 weeks post-hatch. The left chicken pectoralis muscle was dissected out, trimmed free of fat and weighed. Chicken pectoralis is a very large muscle which makes it easier to be handled. It consists almost entirely of one type of fibers (Bandman and Rosser, 2000) and has been revealed to respond to AAS treatment (Allouh and Rosser, 2010). In addition to pectoralis, the tibialis anterior, gastrocnemius externus and gracilis muscles were excised and weighed for comparative purpose between control and Sustanon-treated groups.
Tissue preparation and sectioning: Muscle samples were excised from the cranial half of the superficial region of the main part of the left pectoralis muscle of each bird. Each sample was approximately 0.5x0.5x1.0-2.0 cm. The long axis of the sample was parallel to the direction of the muscle fibers. Samples for immunocytochemistry were coated with optimal cutting temperature compound (O.C.T.; Bio-Optica, Milano, Italy) and then frozen in 2-methylbutane cooled via liquid nitrogen. Samples for DNA and protein analyses were immediately frozen in 2-methylbutane cooled via liquid nitrogen. All samples were then stored at -40°C. Serial cross sections of 10 μm thickness were cut from O.C.T. coated samples at -20°C using a cryostat (Leica, CM3050 S, Nussloch, Germany). Each pair of continuous serial sections was picked up on Superfrost Plus microscopic slides (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA). Two sections were collected on each slide to increase the possibility of choosing better fields for imaging. Serial slides bearing sections were numbered and stored at -40°C. Along with these cross sections, some longitudinal sections were obtained in order to measure the lengths of SCN and myonuclei.
Immunocytochemical protocol: Slides were removed from -40°C and air-dried for 15 min. Slides were fixed with 4% formaldehyde in phosphate-buffered saline (PBS) for 4 min and then washed for 5 min in fresh PBS twice. Blocking solution consisting of 5 mM ethylenediaminetetraacetic acid, 5% goat serum and 1% bovine serum albumin in PBS was applied over the sections for 20 min. The blocking solution was then drained from each slide and combined anti-Pax7 and anti-laminin primary antibodies diluted in blocking solution were applied at a volume of 150 μL per slide. Anti-Pax7 (Developmental Studies Hybridoma Bank, Iowa City, IA, USA) is a mouse monoclonal antibody developed against chicken Pax7 and was used at a dilution of 1:100 to detect SCN (Allouh et al., 2008). Anti-laminin (L9393, Sigma Chemical Co., St. Louis, MO, USA) is a rabbit polyclonal antibody developed against the glycoprotein laminin of mouse origin and was used at a dilution of 1:200 to label the basal laminae of skeletal muscle fibers (Allouh, 2011). After applying the primary antibodies, slides were incubated overnight at 4°C.
The following day, slides were washed in fresh PBS for 5 min twice and secondary antibodies diluted at 1:200 in PBS were applied over the sections on each slide for 40 min at room temperature. Fluorescein isothiocyanate-conjugated anti-mouse (A-11001, Invitrogen Corp., Carlsbad, CA, USA) and tetramethyl rhodamine isothiocyanate-conjugated anti-rabbit (A-11010, Invitrogen Corp.) secondary antibodies were used to label anti-Pax7 green and anti-laminin red, respectively, when viewed with epifluorescent microscopy. Slides were then washed for 5 min twice in PBS. U ltraCruz Mounting Medium containing DAPI (SC-24941, Santa Cruz Biotechnology Inc.) was then applied with cover slips over the sections.
Image analysis and data collection: Six different fields of view were captured from each immunofluorescent slide using a fluorescence microscope (Nikon Eclipse E600, Tokyo, Japan) equipped with a digital still camera. Three epifluorescent images, each viewed through a different wavelength filter, were acquired from each field. The resultant images show all nuclei in blue, SCNs in green and basal laminae in red (Fig. 1). Images were subsequently uploaded to a computer and the three images of each field of view were superimposed using Adobe Photoshop (Adobe System Inc., San Jose, CA, USA).
The minor axis of the ellipse which is similar to the lesser fiber diameter, was used to assess fiber size (Allouh, 2011). The minor axis was chosen to surmount the distortion in the muscle fiber cross section when it is cut at an angle other than transversely. Minor axes of 200 contiguous fiber cross sections were measured from each animal using the Image J program (Abramoff et al., 2004). Numbers of SCNs and myonuclei were counted for each of the 200 fibers. The frequency of SCs was then calculated for each animal using the formula:
Frequency = (SCN/SCN+MN)x100% |
where, MN is myonuclei.
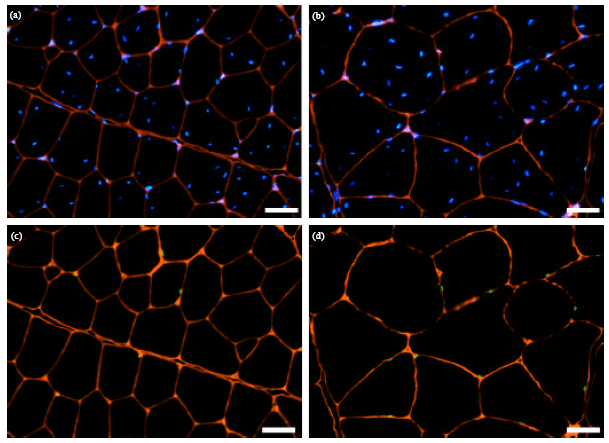 | |
| Fig. 1(a-d): | Immunofluorescent identification of pectoralis muscles of chickens showing DPAPI stained nuclei (blue) and muscle fibre basal laminae (red) in (a) Control and (b) Sustanon-treated birds and anti-Pax7 stained nuclei (green) and basal laminae (red) in (c) Control and (d) Sustanon-treated birds, Scale bars = 50 μm |
The numbers of SCs and myonuclei per unit length of fiber were also calculated for each bird using the formula:
N = A/(Ln+M) |
where, N is the number of cells per unit length of fiber, A is the mean number of nuclei per fiber cross section, Ln is the average length of the nucleus and M is the thickness of the tissue section (Allouh et al., 2008). The lengths of SCNs and myonuclei were measured from the longitudinal sections obtained from different animals in each group. Applying the same protocol used to label SCN and myonuclei within the cross sections, the lengths of 50 SCNs and 100 myonuclei from each animal were measured using the ImageJ program.
The surface area of the sarcolemma per unit length of the fiber was measured using the formula:
S = πxExU |
where, S is the surface area of the sarcolemma per unit length of fiber, E is the ellipse minor axis and U is the unit length of the fiber. The area of muscle fiber sarcolemma per SC was determined by dividing the surface area measurements by the number of SCs for each bird. The myonuclear domain size was calculated for each bird by dividing the volume of sarcoplasm per unit length of fiber by the number of myonuclei in that unit length. The volume of sarcoplasm per unit length of fiber (V) was computed using the formula:
V = π(E/2)2xU |
DNA extraction and quantitation: Total DNA was extracted from preweighted frozen muscle samples (100 mg each) using the DNeasy Blood and Tissue Kit (Qiagen Inc., Valencia, CA, USA) as described previously (Allouh and Aldirawi, 2012). DNA concentrations were determined using the NanoDrop 1000 spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA) at a wavelength of 230 nm.
Western blot analysis: Total muscle protein was isolated by homogenization of preweighted frozen samples (100 mg each) that were crushed into powder and placed in 1.0 mL ice-cold buffer containing 80 mM Tris-HCl (pH 6.8), 100 mM DTT, 70 mM SDS, 1 mM glycerol and freshly added protease inhibitor cocktail (Sigma Chemical Co.). After homogenization the samples were centrifuged for 20 min at 20,000 rpm and the supernatant was transferred to clean microtubes. Total protein concentration was determined using the Bio-Rad Protein Assay (Bio-Rad, Hercules, CA, USA). Sample were then stored at -20°C until Western analysis.
Equal amounts of protein (10 μg) from each sample were loaded on a 10% polyacrylamide mini-gel, along with molecular weight marker (Fermentas Life Sciences, Ontario, Canada) and separated by SDS-PAGE performed with 160 volts at room temperature. Following electrophoresis, the gel was removed and equilibrated in a fresh transfer buffer solution for 10 min at room temperature. The proteins were then transferred to a 0.45 μm nitrocellulose membrane (GE Water and Process Technologies, Trevose, PA, USA) overnight at 200 mA. Following transfer, the membrane was immersed in a blocking solution containing 5% nonfat dry milk (Bio-Rad) dissolved in Tris-buffered saline (pH 7.4) with 0.05% Tween-20 (T-TBS) at room temperature for 1 h. The membrane was then incubated in mouse anti-Pax7 (1:100) (Hybridoma Bank, Iowa City, Iowa) diluted in blocking solution overnight at room temperature. Next day, the membrane was washed 5x10 min in T-TBS and incubated for 2 h in a goat anti-mouse IgG/peroxidase conjugate used at a dilution of 1:500 in T-TBS. After rewashing 3x10 min with T-TBS, the membrane was incubated for 15 min with freshly prepared solution of Diaminobenzidine (DAB) by dissolving 20 mg of DAB, 12 mg nickel chloride in 30 mL carbonate buffer (0.1 M carbonate buffer pH 9.6) containing 24 μL of 30% hydrogen peroxide (H2O2). The membrane was then washed with distilled water, dried, scanned and stored in the dark. Densitometric analysis was performed using Quantity One software (v 4.6.3, Bio-Rad).
Statistical analysis: Samples were divided into two groups (control and Sustanon). Levene’s test for equality of variance was first applied to determine the homogeneity of variance. Data were then evaluated by independent samples t-test at both 5 and 1% levels of significance. All numerical data are expressed as Mean±Standard Error (SE).
RESULTS
Muscle weight: The mean weight of pectoralis muscle±SE was 207.23±7.36 g in the Sustanon-treated group, compared to 134.66±8.97 g in the control group. The wet weight of pectoralis muscle was significantly (p<0.01) greater by approximately 53% in Sustanon-treated birds than in control birds. In addition, the wet weights of tibialis anterior, gastrocnemius externus and Gracilis muscles were significantly (p<0.01) greater in Sustanon-treated birds than in control birds (Table 1).
| Table 1: | Skeletal muscle wet weights (g) in control and Sustanon-treated chickens |
| Values are expressed as mean weight±SE, of n = 5, ** p<0.01 (t-test) | |
| Table 2: | Occurrence of giant fibers in Sustanon-treated chicken pectoralis |
 | |
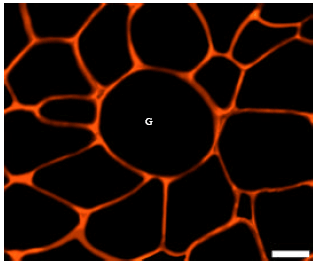 | |
| Fig. 2: | Giant fiber formation in Sustanon-treated birds’ pectoralis muscle, the image shows immun of fluorescent labeling by anti-laminin for the muscle fiber basal laminae in transverse section. The photo shows a giant fiber (G) that appears as exceptionally large fiber with circular cross-sectional area, Scale bar = 50 μm |
Size of muscle fiber: The mean ellipse minor axis±SE in the Sustanon-treated group was 93.78±3.79 μm, compared to 59.42±2.57 μm in the control group. Fibers of the pectoralis muscle were significantly (p<0.01) larger in the Sustanon-treated group than in control group by about 57% (Fig. 1).
Formation of giant fibers: Giant fibers are unusually large fibers that possess circular transverse sectional areas (Fig. 2). These fibers are scattered throughout the muscle as a very small proportion that is usually less than 1% of the total fiber population (Handel and Stickland, 1986). In this study, no giant fibers were found in the control group. However, in Sustanon-treated birds, the mean frequency of giant fibers was 1.00±0.13% (SE). This percent was obtained by examining more than 500 fibers from each bird (Table 2).
Length of SCN and MN: The mean lengths of SCN and myonuclei were determined on longitudinal sections obtained from different animals in each group. There were no significant differences (p>0.05) in the mean lengths of SCN (Table 3) and myonuclei (Table 4) either within or between the groups. These lengths are comparable to those reported in previous studies of the chicken pectoralis (Allouh et al., 2008; Allouh and Rosser, 2010).
Satellite cells: There were approximately 22 more SCs per millimeter of fiber in the Sustanon-treated group than in the control group (Table 3). The frequency of SCs was significantly (p<0.01) greater in Sustanon-treated birds than in controls. The mean number of SCs along 1 mm of fiber length was significantly (p<0.01) higher in sustanon treated birds than in control birds. The surface area of sarcolemma per SC in Sustanon-treated group was significantly (p<0.01) smaller than that in the control group. This finding indicates a greater concentration of SCs in Sustanon-treated birds than in control birds.
Myonuclei: There were approximately 98 more myonuclei per millimeter of fiber in the Sustanon-treated group than in the control group (Table 4). The mean number of myonuclei per fiber cross section was significantly (p<0.05) higher in Sustanon-treated birds than in controls. The mean number of myonuclei along 1 mm of fiber length was significantly (p<0.05) higher in Sustanon-treated birds than in control birds. The myonuclear domain size, determined by calculating the average volume of sarcoplasm per myonucleus, was significantly (p<0.01) larger in Sustanon-treated birds than in control birds.
DNA concentration: The mean DNA concentration in Sustanon-treated muscle samples was significantly (p<0.01) greater than the concentration in control muscle samples. DNA content in Sustanon-treated muscles was twice as much as the DNA content in control muscles (Fig. 3).
Pax7 protein expression: Western blot analysis showed darker bands for Pax7 protein in Sustanon-treated group than in control group (Fig. 4a).
| Table 3: | Satellite cell distribution in control and Sustanon-treated chickens |
 | |
| SC: Satellite cell, Values are expressed as mean±SE, of n = 5, ** p<0.01 (t-test) | |
| Table 4: | Myonuclear distribution in control and Sustanon-treated chickens |
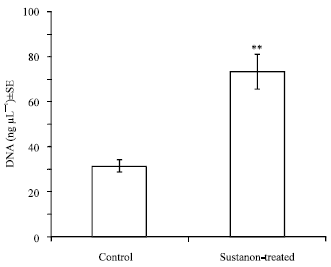 | |
| Fig. 3: | DNA concentration in the pectoralis muscle of control and Sustanon-treated groups. The DNA content was significantly greater in the Sustanon-treated group, **p<0.01, t-test |
The relative density of Pax7 bands were significantly (p<0.05) higher in Sustanon-treated group than in control group (Fig. 4b). This indicates a greater expression of Pax7 protein in the pectoralis muscle of Sustanon-treated chickens compared to control chickens.
DISCUSSION
This study is the first to demonstrate the effects of the drug Sustanon on SC distribution and fiber morphology within avian skeletal muscle. The results reveal that administration of Sustanon over a period of 4 weeks can significantly increase the frequency and concentration of SCs in chicken pectoralis. This is also accompanied by increases in number of myonuclei, DNA content and Pax7 protein level within the muscle.
Sustanon supplementation increases muscle mass and fiber size in the chicken pectoralis without imposed exercise. These findings are in agreement with previous studies that examined the influence of testosterone on human skeletal muscle.
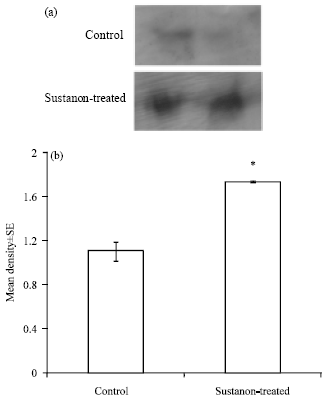 | |
| Fig. 4: | Pax7 protein expression, (a) Western blot was probed with anti-Pax7 antibody against chicken pectoralis extracts from control and Sustanon-treated birds, (b) Densitometric analysis for Pax7 western blot revealed a significant (*p>0.05) difference in Pax7 protein level between control and Sustanon-treated chickens |
Administration of different doses of testosterone enanthate to non-exercising volunteers resulted in a dose-dependent increase in quadriceps muscle volume (Bhasin et al., 2001). Sinha-Hikim et al. (2002) reported that administration of testosterone for 20 weeks to healthy subjects led to muscle hypertrophy without training.
In addition, giant fibers were observed in the pectoralis muscle of birds treated with Sustanon. These are exceptionally large fibers with a circular transverse sectional area that are scattered throughout the muscle in very low numbers. Giant fibers were first discovered by Wohlfart (1949) while studying the sartorius muscle from human fetuses and newborns (Handel and Stickland, 1986). These fibers were originally named B fibers. Giant fibers were subsequently observed in studies of normal stress-resistant and soft pale stress-susceptible porcine skeletal muscles (Cassens et al., 1969; Hendricks et al., 1971; Handel and Stickland, 1986).
There is disagreement in the literature as to whether these giant fibers occur pathologically or not. Wohlfart (1949) stated they are not pathological in nature despite their presence in some forms of muscular atrophy. Also, giant fibers have been observed in normal developing human and guinea pig muscles (Dubowitz, 1965a, b). On the other hand, fibers with histological appearance similar to giant fibers have been found in muscular dystrophy disorders (Schmalbruch, 1982). However, in dystrophy disorders these fibers exist in large groups or sometimes in whole bundles, whereas giant fibers are scattered throughout the muscle and form a small proportion of the total fiber population in non-pathological conditions (Cassens et al., 1969). Also, muscular dystrophy disorders are invariably associated with degenerative changes (Schmalbruch, 1982). Handel and Stickland (1986) have reported no degenerative changes in giant fibers of healthy muscle compared to the large fibers encountered in muscular dystrophies.
The giant fibers observed in Sustanon-treated chickens are likely not associated with any pathological condition. They were scattered throughout the histological sections and represented a very small proportion of the total fiber population. Subsequently, ultra-structural investigation could help determine the exact nature and any potential structural abnormalities of these fibers.
This study shows that Sustanon administration leads to significant increases in all SC indices (frequency, number per millimeter of fiber and concentration) and Pax7 protein expression within avian skeletal muscle. The exact mechanism by which AASs activate SCs is not well known. The action is mainly mediated through androgen receptors (Kadi, 2008). A previous study showed that AASs upregulate androgen receptors within skeletal muscle (Carson et al., 2002). This suggests that Sustanon may directly activate SCs through binding to their androgen receptors. However, it is unclear whether SCs are increased through enhanced replication, inhibition of apoptosis, or both.
Whether SCs are necessary to induce muscle fiber hypertrophy is still a matter of debate. Some studies suggest that activation of SCs is necessary for skeletal muscle hypertrophy since they are the only source for new myonuclei in the muscle (Adams et al., 2002; O'Connor and Pavlath, 2007). For example, ablation of SCs by low doses of gamma irradiation prevented hypertrophy in rodent skeletal muscles (Li et al., 2006). However, other studies have indicated that skeletal muscle is capable of hypertrophy without the need for SCs (Lowe and Alway, 1999; McCarthy et al., 2011). It is speculated that hypertrophy can occur by enhanced transcription of muscle genes within preexisting myonuclei. McCarthy et al. (2011) recently reported robust fiber hypertrophy in SC-depleted mouse skeletal muscle. We assume that, in normal subjects, skeletal muscle fiber hypertrophy occurs coordinately through both SC differentiation into new myonuclei and accelerated transcriptional and translational responses of preexisting myonuclei. However, one of these two mechanisms may predominate according to the influence of certain factors, such as species, age and type of growth stimulus.
Our study showed an increase in the number of myonuclei within hypertrophied muscle fibers treated with Sustanon. It is presumed that myonuclear accretion is necessary to maintain a constant ratio of sarcoplasmic volume per each myonucleus within hypertrophied muscle which is known as myonuclear domain theory (Alway and Siu, 2008). However, this concept has been disputed by others who suggested a loose correlation between sarcoplasmic volume and myonuclear number (Gunderson and Bruusgaard, 2008). Bruusgaard et al. (2006) have reported that the relationship between fiber size and myonuclear number in mouse extensor digitorum longus muscle is lost when the animals mature from young adult to middle age. In this study, despite the increased myonuclear number, the size of myonuclear domain has also significantly increased within hypertrophied muscle fibers. This is comparable to the previous studies that used AASs to induce muscle fiber hypertrophy (Eriksson et al., 2005; Allouh and Rosser, 2010). These findings suggest that the size of the myonuclear domain may not always be correlated with the number of myonuclei within the muscle.
In this study, myonuclei were more easily distinguished because of their more central location within the pectoralis muscle fibers. In mature mammalian muscles, myonuclei are located at the periphery of the muscle fiber and in close proximity to the plasmalemma. Centrally located nuclei in mammalian muscle fibers are considered a sign of muscle pathology (McComas, 1996). As a unique characteristic of a healthy chicken pectoralis muscle, however, myonuclei are normally found deep within the fiber sarcoplasm (Rosser et al., 2002; Allouh et al., 2008).
CONCLUSION
In conclusion, this study supports the concept that Sustanon can induce skeletal muscle hypertrophy in two ways. The first way is through activating SCs which is supported by data demonstrating an increase in the number of these cells. The second way is through enhancing transcription and translation within preexisting myonuclei which is supported by data demonstrating an increase in the size of the myonuclear domain. Future investigations are recommended to determine the differences in fiber type responses to Sustanon and if the increase in SCs by Sustanon is dose-dependent.
ACKNOWLEDGMENTS
This study was funded by a grant from the Deanship of Research at JUST (25/2010). The authors would like to thank the Princess Haya Biotechnology Center for help and support in providing the facility and equipment required for this study. The Pax7 monoclonal antibody developed by A. Kawakami was obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by the University of Iowa, Department of Biology, Iowa City, IA 52242.
REFERENCES
- Abramoff, M.D., P.J. Magelhaes and S.J. Ram, 2004. Image processing with image. Biophotonics Int., 11: 36-42.
Direct Link - Adams, G.R., V.J. Caiozzo, F. Haddad and K.M. Baldwin, 2002. Cellular and molecular responses to increased skeletal muscle loading after irradiation. Am. J. Physiol. Cell Physiol., 283: C1182-C1195.
Direct Link - Allouh, M.Z., 2011. Effect of Ferula hermonis root extract on rat skeletal muscle adaptation to exercise. Exp. Biol. Med., 236: 1373-1378.
CrossRefPubMedDirect Link - Allouh, M.Z. and B.W. Rosser, 2010. Nandrolone decanoate increases satellite cell numbers in the chicken pectoralis muscle. Histol. Histopathol., 25: 133-140.
PubMedDirect Link - Allouh, M.Z. and M.H. Aldirawi, 2012. Influence of Mesterolone on satellite cell distribution and fiber morphology within maturing chicken pectoralis muscle. Anatom. Rec., 295: 792-799.
CrossRefDirect Link - Allouh, M.Z., Z. Yablonka-Reuveni and B.W. Rosser, 2008. Pax7 reveals a greater frequency and concentration of satellite cells at the ends of growing skeletal muscle fibers. J. Histoch. Cytochem., 56: 77-87.
CrossRefPubMedDirect Link - Alway, S.E. and P.M. Siu, 2008. Nuclear apoptosis contributes to sarcopenia. Exerc. Sport Sci. Rev., 36: 51-57.
PubMed - Bandman, E. and B.W. Rosser, 2000. Evolutionary significance of myosin heavy chain heterogeneity in birds. Microsc. Res. Tech., 50: 473-491.
CrossRefPubMedDirect Link - Beg, T., Y.H. Siddique, G. Ara, J. Gupta and M. Afzal, 2008. Antioxidant effect of ECG on testosterone propionate induced chromosome damage. Int. J. Pharmacol., 4: 258-263.
CrossRefDirect Link - Bhasin, S., L. Woodhouse and T.W. Storer, 2001. Proof of the effect of testosterone on skeletal muscle. J. Endocrinol., 170: 27-38.
CrossRefDirect Link - Ahmed Bin Bisher, A.S., 2009. Histopathological evidences of the nephritic pathological alterations induced by the anabolic androgenic drug (Sustanon) in male guinea pigs (Cavia porcellus). J. Biol. Sci., 9: 514-523.
CrossRefDirect Link - Carson, J.A., W.J. Lee, J. McClung and J.A. Hand, 2002. Steroid receptor concentration in age rat hindlimb muscle: Effect of anabolic steroid administration. J. Applied Physiol., 93: 242-250.
PubMed - Cassens, R.G., C.C. Cooper and E.J. Briskey, 1969. The occurrence and histochemical characterization of giant fibers in the muscle of growing and adult animals. Acta Neuropathol., 12: 300-304.
CrossRef - Dhawan, J. and T.A. Rando, 2005. Stem cells in postnatal myogenesis: Molecular mechanisms of satellite cell quiescence, activation and replenishment. Trends Cell Biol., 15: 666-673.
PubMed - Dubowitz, V., 1965. Enzyme histochemistry of skeletal muscle. Part I. Developing animal muscle. J. Neurol. Neurosurg. Psychiat., 28: 516-519.
Direct Link - Ebrahimi, M., 2009. Anabolic steroid exposure of athletes of Qom, Iran, through performance enhancing drugs use. Res. J. Environ. Toxicol., 3: 50-55.
CrossRefDirect Link - Eriksson, A., F. Kadi, C. Malm and L.E. Thornell, 2005. Skeletal muscle morphology in power-lifters with and without anabolic steroids. Histochem. Cell Biol., 124: 167-175.
PubMed - Fitts, J.M., M.R. Klein and C.A. Powers, 2004. Comparison of tamoxifen and testosterone propionate in male rats: Differential prevention of orchidectomy effects on sex organs, bone mass, growth and the growth hormone-IGF-I axis. J. Androl., 25: 523-534.
PubMed - Gunderson, K. and J.C. Bruusgaard, 2008. Nuclear domains during muscle atrophy: nuclei lost or paradigm lost?. J. Physiol., 586: 2675-2681.
CrossRef - Handel, S.E. and N.C. Stickland, 1986. Giant muscle fibers in skeletal muscle of normal pigs. J. Comp. Pathol., 96: 447-457.
PubMed - Hawke, T.J. and D.J. Garry, 2001. Myogenic satellite cells: Physiology to molecular biology. J. Applied Physiol., 91: 534-551.
Direct Link - Hendricks, H.B., D.T. Lafferty, E.D. Aberle, M.D. Judge and J.C. Forrest, 1971. Relation of porcine muscle fiber type and size to postmortem shortening. J. Animal Sci., 32: 57-61.
Direct Link - Kadi, F., 2008. Cellular and molecular mechanisms responsible for the action of testosterone on human skeletal muscle. A basis for illegal performance enhancement. Br. J. Pharmacol., 154: 522-528.
CrossRefPubMedDirect Link - Li, P., T. Akimoto, M. Zhang, R.S. Williams and Z. Yan, 2006. Resident stem cells are not required for exercise-induced fiber-type switching and angiogenesis but are necessary for activity-dependent muscle growth. Am. J. Physiol. Cell Physiol., 290: C1461-C1468.
CrossRefDirect Link - Lowe, D.A. and S.E. Alway, 1999. Stretch-induced myogenin, MyoD and MRF4 expression and acute hypertrophy in quail slow-tonic muscle are not dependent upon satellite cell proliferation. Cell Tissue Res., 296: 531-539.
CrossRefDirect Link - Marjani, M., S. Jamili, P.G. Mostafavi, M. Ramin and A. Mashinchian, 2009. Influence of 17-alpha methyl testosterone on masculinization and growth in Tilapia (Oreochromis mossambicus). J. Fish. Aquat. Sci., 4: 71-74.
CrossRefDirect Link - McCarthy, J.J., J. Mula, M. Miyazaki, R. Erfani and K. Garrison et al., 2011. Effective fiber hypertrophy in satellite cell-depleted skeletal muscle. Development, 138: 3657-3666.
CrossRefPubMedDirect Link - Mohamed, R.A., H.M. El Aasar, L.A. Mohamed and A.M. Abbas, 2007. Morphological features of normal human skeletal muscle in different age groups: A histological and ultrastructural study. J. Medical Sci., 7: 161-169.
CrossRefDirect Link - Moore, D.T., P.R. Ferket and P.E. Mozdziak, 2005. Muscle development in the late embryonic and early post-hatch poult. Int. J. Poult. Sci., 4: 138-142.
CrossRefDirect Link - Morgan, J.E. and T.A. Partridge, 2003. Muscle satellite cells. Int. J. Biochem. Cell Biol., 35: 1151-1156.
CrossRefDirect Link - O'Connor, R.S. and G.K. Pavlath, 2007. Point:Counterpoint: Satellite cell addition is/is not obligatory for skeletal muscle hypertrophy. J. Applied Physiol., 103: 1099-1100.
CrossRefDirect Link - Pala, A., 2003. Effects of agouti related protein and androgens on growth. J. Applied Sci., 3: 197-202.
CrossRefDirect Link - Pophal, S., P.E. Mozdziak and S.L. Vieira, 2004. Satellite cell mitotic activity of broilers fed differing levels of lysine. Int. J. Poult. Sci., 3: 758-763.
CrossRefDirect Link - Rosser, B.W.C., M.S. Dean and E. Bandman, 2002. Myonuclear domain size varies along the lengths of maturing skeletal muscle fibers. Int. J. Dev. Biol., 46: 747-754.
Direct Link - Sinha-Hikim, I., J. Artaza, L. Woodhouse, N. Gonzalez-Cadavid and A.B. Singh et al., 2002. Testosterone-induced increase in muscle size in healthy young men is associated with muscle fiber hypertrophy. Am. J. Physiol. Endocrinol. Metab., 283: E154-E164.
Direct Link - Sinha-Hikim, I., W.E. Taylor, N.F. Gonzalez-Cadavid, W. Zheng and S. Bhasin, 2004. Androgen receptor in human skeletal muscle and cultured muscle satellite cells: Up-regulation by androgen treatment. J. Clin. Endocrinol. Metab., 89: 5245-5255.
CrossRefDirect Link - Velleman, S.G., K.E. Nestor, C.S. Coy, I. Harford and N.B. Anthony, 2010. Effect of posthatch feed restriction on broiler breast muscle development and muscle transcriptional regulatory factor gene and heparan sulfate proteoglycan expression. Int. J. Poult. Sci., 9: 417-425.
CrossRefDirect Link - Wohlfart, G., 1949. Muscular atrophy in diseases of the lower motor neuron. Arch. Neurol. Psychiatry, 61: 599-620.
Direct Link - Wozniak, A.C., J. Kong, E. Bock, O. Pilipowicz and J.E. Anderson, 2005. Signaling satellite-cell activation in skeletal muscle: Markers, models, stretch and potential alternate pathways. Muscle Nerve, 31: 283-300.
CrossRefPubMedDirect Link - Zammit, P.S., T.A. Partridge and Z. Yablonka-Reuveni, 2006. The skeletal muscle satellite cell: The stem cell that came in from the cold. J. Histochem. Cytochem., 54: 1177-1191.
CrossRefDirect Link
cheap steroids uk Reply
I bought testosterone boosters to help my muscles, increase my energy, and metabolism how do i block estrogen?