
Wu Yingben
Department of Environmental Microbiology, Hunan Institute of Microbiology, Changsha, Hunan 410009, China
He Yuelin
Department of Environmental Microbiology, Hunan Institute of Microbiology, Changsha, Hunan 410009, China
Yin Hongmei
Department of Environmental Microbiology, Hunan Institute of Microbiology, Changsha, Hunan 410009, China
Chen Wei
Department of Environmental Microbiology, Hunan Institute of Microbiology, Changsha, Hunan 410009, China
Wang Zhen
Department of Environmental Microbiology, Hunan Institute of Microbiology, Changsha, Hunan 410009, China
Xu Lijuan
Department of Environmental Microbiology, Hunan Institute of Microbiology, Changsha, Hunan 410009, China
Zhang Aiqun
Department of Environmental Microbiology, Hunan Institute of Microbiology, Changsha, Hunan 410009, China
Pakistan Journal of Biological Sciences
Year: 2012 | Volume: 15 | Issue: 23 | Page No.: 1144-1151







ABSTRACT
Microorganisms have been obtained to improve the agronomic value of Rock Phosphates (RPs), but the phosphorus solubilizing rate by these approaches is very slow. It is important to explore a high-efficient phosphate-solubilizing approach with a kind of microorganisms. This study aimed to isolate a high-efficient level of phosphate-solubilizing fungus from rhizosphere soil samples phosphate mines (Liuyang County, Hunan province, China) and apply it in solubilization of RPs. The experiments were carried out by the conventional methodology for morphological and biochemical fungus characterization and the analysis of 18s rRNA sequence. Then the effects of time, temperature, initial pH, phosphorus (P) sources, RPs concentration, shaking speed and silver ion on the content of soluble P released by this isolate were investigated. The results showed this isolate was identified as Galactomyces geotrichum P14 (P14) in GeneBank and the maximum amount of soluble P was 1252.13 mg L-1 within 40 h in a modified phosphate growth agar’s medium (without agar) where contained tricalcium phosphate (TCP) as sole phosphate source. At the same time, it could release phosphate and solubilize various rock phosphates. The isolated fungus can convert RPs from insoluble form into plant available form and therefore it hold great potential for biofertilizers to enhance soil fertility and promote plant growth.
PDF Abstract XML References Citation
Received: November 30, 2012;
Accepted: March 04, 2013;
Published: March 09, 2013
How to cite this article
Wu Yingben, He Yuelin, Yin Hongmei, Chen Wei, Wang Zhen, Xu Lijuan and Zhang Aiqun, 2012. Isolation of Phosphate-solubilizing Fungus and Its Application in Solubilization of Rock Phosphates. Pakistan Journal of Biological Sciences, 15: 1144-1151.
DOI: 10.3923/pjbs.2012.1144.1151
URL: https://scialert.net/abstract/?doi=pjbs.2012.1144.1151
DOI: 10.3923/pjbs.2012.1144.1151
URL: https://scialert.net/abstract/?doi=pjbs.2012.1144.1151
INTRODUCTION
Phosphorus (P) is one of the major macronutrients of plants for their growth and development (Vassilev and Vassileva,. 2003; Mittal et al., 2008). Although, soils of the China cropland are often high in total P, only limited quantities are available to crop plants (Zapata and Zaharah, 2002). Thus, it is necessary to develop phosphatic fertilizer with high performance in the soil for optimum crop yield (Sharpley, 1995). Generally, phosphatic fertilizer is produced globally by the mining and chemical processing of Rock Phosphates (RPs), a non-renewable natural resource. Currently, the complete dissolution of the ore by chemical processing usually results in the release of undesirable contaminants with environmental pollution.
Microorganisms play a critical role in natural phosphorus cycle (Biswas and Narayanasamy, 2006; Vassilev et al., 2006) and recently, microbially-based approach can improve the agronomic value of RPs to a certain extent. Microbial solubilization of RPs is gaining great attentions in agriculture. Some organisms have been isolated for the purpose (Kucey, 1987; Kumar et al., 2001; Ahuja et al., 2007; Achala et al., 2007; Coutinho et al., 2011; Gupta et al., 2012; Lu et al., 2012). These phosphate solubilizers can be used directly as biofertilizers in the soil or bioreactors for the bio-processing of RPs. This approach not only compensates for higher cost of manufacturing phosphate fertilizer in industry but also reduces environment pollution caused by traditional chemical process.
Under the situation, many phosphate-solubilizing microorganisms have been isolated from different environments and phosphorus availability to plants (Yu et al., 2012) through the inoculation of phosphate-solubilizing microorganisms has been widely studied under pot and field conditions (Duponnois et al., 2005; Valverde et al., 2006). However, in fact, these microorganisms are only a small percentage of the total microbial population and few of them present a high potential to solubilize RPs under natural conditions which seriously restrains the application of this microbially-based technique. Moreover, a main problem, indeed, is that how to remain a high destiny and activity of the introduced microorganisms (Van Veen et al., 1997). Normally, many isolates present high phosphate-solubilizing capability in growth medium, but when they are inoculated to natural environment, they are often in a form characterized by non-growth and/or low phosphate-solubilizing capability (Xiao et al., 2008). Therefore, it is significant and necessary to isolate new and potential phosphate-solubilizing microorganisms.
Here, the experiments were carried out by isolating and characterizing phosphate-solubilizing fungus from rhizosphere soil samples around phosphate mines in Liuyang County and the effects of time, temperature, initial pH, P sources, RPs concentration, shaking speed and silver ion on the content of soluble P released by this isolate were investigated.
MATERIALS AND METHODS
Isolation and identification of phosphate-solubilizing fungus: The fungus strain was isolated from rhizosphere soil samples collected from the various agricultural fields located around phosphate mines in Liuyang County. Various serial dilutions of soil samples with 100 μL aliquots were plated on a Modified Phosphate Growth Agar’s (MPGA) medium (glucose, 10 g; (NH4)2SO4, 0.15 g; KCl, 0.2 g; MgCl2•6H2O, 0.5 g; MgSO4•7H2O, 0.25 g; agar, 20 g; distilled water, 1000 mL) supplemented with ampicillin (50 μg mL-1) and streptomycin (30 μg mL-1) and containing tricalcium phosphate (TCP, Ca3(PO4)2, 10.0 mg mL-1). Here TCP was chosen as sole P source for selectively screening strains which have the capability to solubilize unsoluble-P and release soluble P from TCP. After incubation for 3 days at 28°C, the strain developed clear zones around colonies. Colonies with clear zones were further purified by replanting on MPGA’s medium. The screened phosphate-solubilizing fungus was selected and stored for further work. The isolate was identified on the basis of morphological, physiological and biochemical characterization and sequence of 18S rRNA gene.
Morphological characterization, 18S rRNA gene sequencing and phylogenetic analysis of the isolated fungus: In order to identify the isolated fungus strain for a correct species, the morphology characteristics of the isolate were evaluated according to the methods described by Wei (1979). Physiological and biochemical characteristics were performed as per standard procedures by plate assays.
Primers for the 18S rRNA were used to amplify a DNA sequence (primers NS1 [5’- GTAGTCATATGCT TGTCTC -3’] and NS2 [5’- TGCTGGCACCAGACTTG -3’]) by Polymerase Chain Reaction (PCR) (White et al., 1990). Primers were synthesized by GenScript Corporation Ltd. (NanJing, China). PCR amplifications were carried out in 50 μL reaction volumes with a PCR reactor (Eppendorf) with following program: initial denaturation at 94°C for 3 min followed by 30 cycles of denaturation at 94°C for 1 min, annealing at 55°C for 1 min and extension at 72°C for 2 min followed by a final extension phase at 72°C for 10 min. The purified PCR product was sequenced in both the directions. The sequences of 18S rRNA genes were first analyzed using the BLAST searching program at the National Center for Biotechnology Information (NCBI) website:http://www.ncbi.nlm.nih.gov/BLAST/. Sequence alignments were performed using MEGA-5 software. The 18S rRNA sequence of the isolate was aligned with reference sequences and other close relatives from GenBank. A maximum likelihood phylogenetic tree was constructed using MEGA version 5.0. To estimate the confidence of the phylogenetic tree, bootstrapping was performed (Tamura et al., 2007).
Culture medium and solubilizing conditions: P solubilization experiments were carried out in flasks with 100 mL (in 500 mL conical flask) of MPGA’s medium (without agar) and 5.0 g TCP as sole P source (Mittal et al., 2008). Cells suspensions of the isolate were counted by a haemocytometer to adjust the count to approximately 107 cells mL-1. Each flask was inoculated with the cell suspensions at 10% (v/v). Flasks were shaken under 160 rpm at 28°C for 2 days. The pH of the culture medium was measured by pH electrode and periodically adjusted and maintained at 7.0 by adding sterile 2% H2SO4 or NaOH solution. All experiments were performed supplemented with triplicate.
Experimental design
Optimization of time, temperature and initial pH: The effects of the reaction time, temperature and initial pH on P solubilization by the isolate were investigated. Cell suspensions of the isolate were inoculated into 100 mL MPGA’s medium supplemented with 5.0 g TCP. The isolates were incubated at different time (0, 8, 16, 24, 32, 40, 48, 60, 72 and 96 h), temperatures (20, 23, 25, 28, 30, 32, 34 and 37°C) and different initial pH (5.0, 5.5, 6.0, 6.5, 7.0, 7.5 and 8.0) for 2 days. The initial pH of the culture medium was periodically adjusted and maintained by adding sterile 2% (v/v) H2SO4 or NaOH solution.
P sources: Different P sources for RPs solubilization by the isolate were investigated. TCP was replaced with aluminium phosphate (AlPO4), different RPs from Liuyang, Shimen and Yuanling phosphate mines (Hunan Province). The main chemical compositions of Liuyang RP sample are 8.29% P, 23.67% Ca, 9.47%, Si, 10.12% Mg, 1.31% Na, 0.87% Al and 0.47% Fe. Shimen RP sample contains 7.82% P, 24.14% Ca, 10.51% Si, 6.75% Mg, 1.64% Na, 0.47% Al and 1.59% Fe. Yuanling RP sample contains 6.67% P, 25.07% Ca, 13.92% Si, 6.74% Mg, 0.64% Na, 0.99% Al and 1.83% Fe. The sample was comminuted to a particle size of almost 300 meshes (0.05 mm).
RPs concentration: The effect of changing concentration of RPs on its solubilization by the isolate was studied. RPs concentration in the medium was 2.0, 3.0, 4.0, 5.0, 6.0, 7.0 and 8.0 g L-1, respectively.
Shaking speed: The shaking speed on Liuyang RP, Shimen RP and Yuanling RP solubilization by the isolate was optimized. Flasks were shaken under 80, 100, 120, 140, 160, 180 and 200 rpm, respectively for 2 days.
Silver ion: Heavy metals impose high toxicity to microorganisms. The interaction between heavy metals and microorganisms has perhaps attracted great interest to researchers due to its application in metallurgy and decontamination of the environment (White et al., 1990). Here, silver ion (Ag+) was selected as a representative of heavy metals to study the influence of heavy metals on RPs solubilization by the isolate. Different concentrations (0, 50, 100, 150, 200, 250 and 300 mg L-1, respectively) of AgNO3 were added to the culture medium to study the influence of Ag+ on RPs solubilization by the isolate.
Analytical methods: After shaking for 2 days, a certain volume of fresh MPGA’s medium was added to each flask and the culture medium was fixed to 100 mL by a volumetric flask. The 100 mL culture medium was centrifuged at 9000 rpm for 20 min and the supernatant was collected. The content of soluble P in a form of TCP remaining in the supernatant was determined by phosphomolybdate method (Xiao et al., 2008) with a UV-VIS 8500 spectrophotometer at 420 nm.
Statistical analysis: Capability of RPs solubilization was determined by the content of soluble P in the supernatant liquid. Values are given as means±SD. for triplicate samples. Data were analyzed by analysis of variance (ANOVA) and the means were compared with Duncan’s Multiple Range Test (DMRT) at p<0.05 level.
RESULTS AND DISCUSSION
Most of natural RPs is low-grade in China. Thus they do not be directly utilized efficiently and are usually thrown away. However, they have been recognized as a valuable alternative for phosphate fertilizer. In view of environmental concerns and current developments in sustainability, researcher’s efforts concentrating on elaboration of techniques that involve the use of less expensive and bio-available sources such as natural RPs has attracted widespread attentions (Whitelaw, 2000). The advantage of using natural phosphate mines soil to isolate over the genetically manipulation is its easier adaptation and succession when inoculated into the medium containing RPs.
Identification of the isolates and phylogenetic tree analysis: The phosphate-solubilizing fungus was white cylindrical fungus. The isolate was firstly identified based on morphological characteristics (Fig. 1). The full results of morphology, physiological and biochemical characteristics were summarized (Table 1). On MPGA’s medium the color of the colony was always white. Reverse of the colony was light yellow. Conidiophores and Phialides were observed. Conidiophores were white, smooth walled with verticillate branches. Phialides were hyaline, cylindrical and were arranged in line.
Then, the 18S rRNA gene sequences of the isolate was determined and a phylogenetic tree was constructed (Fig. 2). The 914 bp nucleotide sequence of the strain has been deposited in the GenBank nucleotide sequence data library under the following accession number JX110709. The 18S rRNA gene sequences comparison revealed that the strain had 83% similarity with Galactomyces geotrichum. Based on their 18S rRNA gene sequences and phylogenetic positions, the isolate was designated as Galactomyces geotrichum P14.
Optimum time, temperature and initial pH for P solubilization: The effect of incubation time, temperature and initial pH on P solubilization was investigated based on the method described in experimental design.
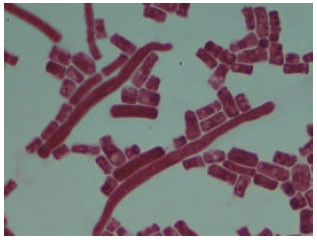 | |
| Fig. 1: | Morphology of the Galactomyces geotrichum P14 |
| Table 1: | Morphological, physiological and biochemical characteristics of the isolate |
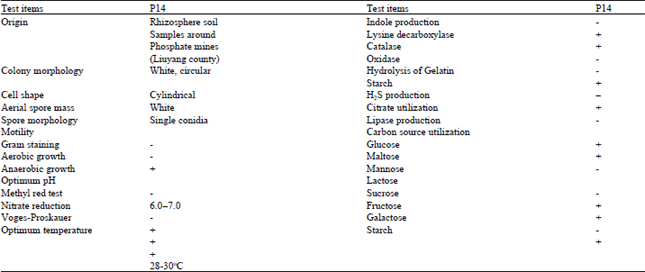 | |
| +: Means positive result, –: Means negative result | |
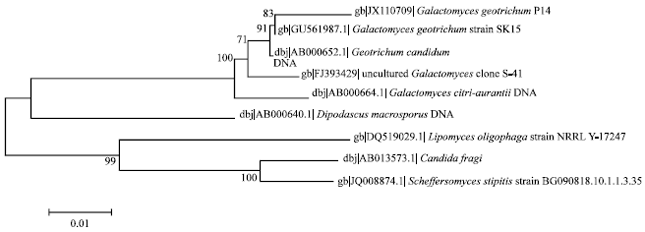 | |
| Fig. 2: | 18S rRNA-based phylogenetic relationship between the isolate and representatives of other related taxa (GenBank accession numbers in parentheses). The numbers at the nodes indicate the levels of bootstrap support based on data for 1,000 replicates; values inferred greater than 50% are only presented. The scale bar indicates 0.01 substitutions per nucleotide position |
Above-mentioned factors had significant influence on P solubilization. The isolate demonstrated high efficiency of phosphate-solubilizing capability within 40 hours, the maximum amount of soluble P was 1252.13 mg L-1 (Fig. 3) released by Galactomyces geotrichum P14, the effect of powerful and efficient phosphate-solubilizing capability was not reported previously (Keyes, 1990; Gleddie et al., 1993; Achala et al., 2007; Collavino et al., 2010; Coutinho et al., 2011; Gupta et al., 2012; Lu et al., 2012). Longer or less than the optimal time, the amount of soluble P decreased. Otherwise, the maximum amount of soluble P was recorded at 30°C which was 1047.34 mg L-1 (Fig. 4). Then, initial pH is concerned. The maximum amount of soluble P was recorded at 1034.64 mg L-1 (Fig. 5) released by the P14 at initial pH 7.0. Higher or lower than the optimal temperature and initial pH, the amount of soluble P decreased correspondingly.
Generally, it might be due to the increasing of the production of organic acid metabolites by different phosphate-solubilizing microorganisms and these metabolites can convert the insoluble RP into soluble P (Tributsch, 2001; Illmer and Schinner, 1995; Rashid et al., 2004; Kim et al., 2005). However, the solubilizing mechanism by the P14 should be further studies.
Solubilization of different P sources: The isolate was capable of solubilizing five kinds of RPs, however, the solubilization capability of the RPs varied with different P sources (Fig. 6).
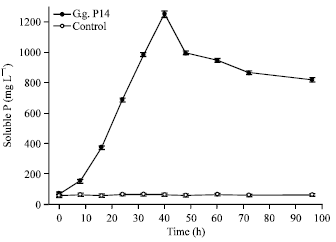 | |
| Fig. 3: | Evolution of the amount of soluble P along with time |
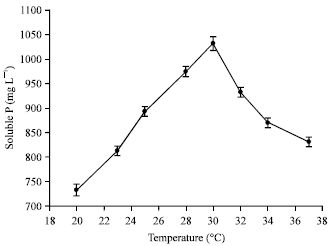 | |
| Fig. 4: | Effect of temperature on the P solubilization |
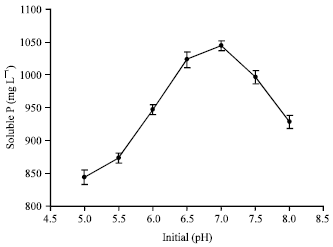 | |
| Fig. 5: | Effect of initial pH on the P solubilization |
The results revealed that the maximum amount of soluble P released by the isolate averaged 1003.57 mg L-1 in presence of TCP, followed by aluminium phosphate (average 771.65 mg L-1), Liuyang RP (average 511.43 mg L-1), Shimen RP (average 441.74 mg L-1) and Yuanling RP (average 406.02 mg L-1), respectively.
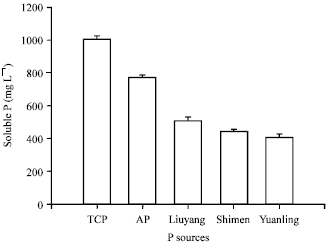 | |
| Fig. 6: | Effect of P sources on P solubilization. (Note: TCP, tricalcium phosphate; AP, aluminium phosphate; Liuyang, Liuyang rock phosphate; Shimen, Shimen rock phosphate; Yuanling, Yuanling rock phosphate) |
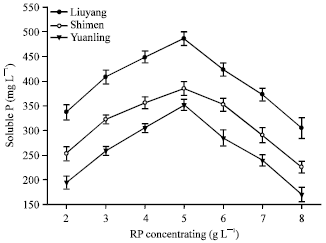 | |
| Fig. 7: | Effect of RP concentration on content of soluble P |
Interestingly, the total P content of RPs was also similar in order. It might be concluded that the capability of RPs solubilization was positively correlated with the grade of RPs. The higher the grade of RPs, the higher the content of soluble P released.
Optimum of RP concentration for RPs solubilization: The optimum RP concentration for RPs solubilization was investigated based on the method described in experimental design. The isolate presented different soluble P releasing ability in the medium with various RPs concentrations (Fig. 7). The maximum amount of soluble P was recorded at 485.83 mg L-1 when added 5.0 g L-1 Liuyang RP was added in the medium inoculated with the P14, followed by Shimen RP (385.34 mg L-1) and Yuanling RP (351.86 mg L-1).
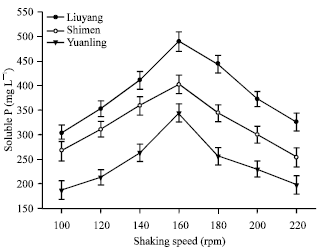 | |
| Fig. 8: | Effect of shaking speed on content of soluble P |
It was also observed that as the concentration of added RPs decreased from 5-2 g L-1, there was a slight increase in RPs solubilization capability as the content of soluble P increased. But as the concentration of added RPs increased from 5-8 g L-1, there was no increase but a visible decrease in the content of soluble P.
Effect of shaking speed on RPs solubilization: The amount of soluble P was released by the P14 at different shaking speeds ranging from 80-200 rpm (Fig. 8). The amount of soluble P increased with the increase of shaking speed from 80-160 rpm. However, the amount of soluble P was decreased, when the shaking speed increased from 160-200 rpm. The reason was the growth of the isolate was often weakened by the shear stress resulting from the strong stirring (Xiao et al., 2008). The maximum content of soluble P released by the P14 was recorded at 488.43 mg L-1 (Liuyang RP) when shaking was done at 160 rpm, followed by Shimen RP (401.74 mg L-1) and Yuanling RP (344.02 mg L-1). However, further studies are required to determine their efficiency in solubilizing RPs under natural conditions (Reyes et al., 1999).
Effect of silver ion on RPs solubilization: Toxic effects of heavy metals include blocking of functional groups biologically and denaturation of enzymes. The effects of different concentrations of silver ion (Ag+) on the content of soluble P released by the isolate are shown in Fig. 9. The concentrations used in these experiments were considered to be toxic to this fungus. The isolate was shown to have sensitivity and intolerance with increasing concentration of Ag+ which was consistent with that reported in previous studies (Xiao et al., 2008).
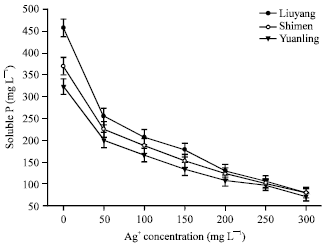 | |
| Fig. 9: | Effect of concentration of Ag+ on content of soluble P |
With the increasing of Ag+ concentration, the content of soluble P decreased sharply. Among all the RPs, the maximal decrease of 44.26% content of soluble P was Liuyang RP and followed by Shimen RP (decreased 39.05%) and Yuanling RP (decreased 37.97%). Moreover, the additive sliver ion could form a small quantity of white flocculent precipitation in the culture medium.
CONCLUSION
A high-efficient level of phosphate-solubilizing fungus was isolated from rhizosphere soil samples phosphate mines, identified as Galactomyces geotrichum P14 in GeneBank. The study also demonstrated the capability of solubilizing of RPs obviously varies from the resource of RPs. In addition, the P14 is the powerful RPs solubilizer in MPGA’s medium, suggesting a high soluble P releasing ability and good adaptation. The optimum conditions for the p14 releasing soluble P from RPs were at temperature, 30°C; initial pH, 7.0; shaking speed, 160 rpm; RPs concentration, 5.0 g L-1, respectively and approximately close to the growth conditions of this fungus. The maximal content of soluble P can be achieved when TCP as the sole P source in the medium inoculated with the p14. Moreover, the isolate showed intolerance with increasing concentration of silver ion (Ag+).
The action of microorganisms leading to solubilization of minerals is recognized as direct and indirect action, however, the solubilizing mechanism by the p14 should be further studies.
ACKNOWLEDGMENTS
This research was financially supported by the projects of the China Hunan Provincial Agriculture Department ([2009]76) and the China Hunan Provincial Science and Technology Department (2009FJ1005). The authors are grateful to Hunan University, Changsha and Gene Bank, USA.
REFERENCES
- Achala, V., V.V. Savantb and M.S. Reddya, 2007. Phosphate solubilization by a wild type strain and UV-induced mutants of Aspergillus tubingensis. Soil Biol. Biochem., 39: 695-699.
CrossRefDirect Link - Ahuja, A., S.B. Ghosh and S.F. D'Souza, 2007. Isolation of a starch utilizing, phosphate solubilizing fungus on buffered medium and its characterization. Bioresour. Technol., 98: 3408-3411.
PubMed - Sharpley, A.N., 1995. Soil phosphorus dynamics: Agronomic and environmental impacts. Ecol. Eng., 5: 261-279.
CrossRef - Biswas, D.R. and G. Narayanasamy, 2006. Rock phosphate enriched compost: An approach to improve low-grade Indian rock phosphate. Bioresour. Technol., 97: 2243-2251.
PubMed - Collavino, M.M., P.A. Sansberro, L.A. Mroginski and O.M. Aguilar, 2010. Comparison of in vitro solubilization activity of diverse phosphate-solubilizing bacteria native to acid soil and their ability to promote Phaseolus vulgaris growth. Biol. Fertil. Soils, 46: 727-738.
CrossRefDirect Link - Coutinho, F.P., M.A.Q. Cavalcanti and A.M. Yano-Melo, 2011. Phosphate-solubilizing fungi isolated from a semiarid area cultivated with melon (Cucumis melo L. cv. gold mine). Acta Bot. Bras., 25: 929-931.
CrossRef - Duponnois, R., A. Colombet, V. Hien and J. Thioulouse, 2005. The mycorrhizal fungus Glomus intraradices and rock phosphate amendment influence plant growth and microbial activity in the rhizosphere of Acacia holosericea. Soil Biol. Biochem., 37: 1460-1468.
Direct Link - Gupta, M., S. Kiran, A. Gulati, B. Singh and R. Tewari, 2012. Isolation and identification of phosphate solubilizing bacteria able to enhance the growth and aloin-A biosynthesis of Aloe barbadensis Miller. Microbiol. Res., 167: 358-363.
CrossRef - Illmer, P. and F. Schinner, 1995. Solubilization of inorganic calcium phosphates-solublization mechanisms. Soil Biol. Biochem., 27: 257-263.
CrossRefDirect Link - Kim, Y.H., B. Bae and Y.K. Choung, 2005. Optimization of biological phosphorus removal from contaminated sediments with phosphate solubilizing microorganisms. J. Biosci. Bioeng., 1: 23-29.
CrossRefDirect Link - Kucey, R.M.N., 1987. Increased phosphorus uptake by wheat and field beans inoculated with a phosphorus-solubilizing Penicillium bilaji strain and with vesicular-arbuscular mycorrhizal fungi. Applied Environ. Microbiol., 53: 2699-2703.
Direct Link - Kumar, V., R.K. Behl and N. Narula, 2001. Establishment of phosphate-solubilizing strains of Azotobacter chroococcum in the rhizosphere and their effect on wheat cultivars under greenhouse conditions. Microbiol. Res., 156: 87-93.
Direct Link - Lu, J., X. Gao, Z. Dong and L. An, 2012. Expression of mitochondrial malate dehydrogenase in Escherichia coli improves phosphate solubilization. Ann. Microbiol., 62: 607-614.
CrossRef - Mittal, V., O. Singh, H. Nayyar, J. Kaur and R. Tewari, 2008. Stimulatory effect of phosphate-solubilizing fungal strains (Aspergillus awamori and Penicillium citrinum) on the yield of chickpea (Cicer arietinum L. cv. GPF2). Soil Biol. Biochem., 40: 718-727.
CrossRef - Rashid, M., S. Khalil, N. Ayub, S. Alam and F. Latif, 2004. Organic acids production and phosphate solubilization by Phosphate Solubilizing Microorganisms (PSM) under in vitro conditions. Pak. J. Biol. Sci., 7: 187-196.
CrossRefDirect Link - Reyes, I., L. Bernier, R.R. Simard and H. Antoun, 1999. Effect of nitrogen source on solubilization of different inorganic phosphates by an isolate of Pencillium rugulosum and two UV-induced mutants. FEMS Microbiol. Ecol., 28: 281-290.
CrossRefDirect Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Valverde, A., A. Burgos, T. Fiscella, R. Rivas, E. Velazquez, C. Rodriguez and J.M. Igual, 2006. Differential effects of co inoculations with Pseudomonas jessenii PS06 (a phosphate solubilizing bacterium) and Mesorhizobium ciceri c-2/2 strains on the growth and seed yield of chickpea under greenhouse and field conditions. Plant Soil, 287: 43-50.
Direct Link - Van Veen, J.A., L.S. Van Overbeek and J.D. Van Elsas, 1997. Fate and activity of microorganisms introduced into soil. Microbiol. Mol. Biol. Rev., 61: 121-135.
Direct Link - Vassilev, N. and M. Vassileva, 2003. Biotechnological solubilization of rock phosphate on media containing agroindustrial wastes. Appl. Microbiol. Biotechnol., 61: 435-440.
CrossRefDirect Link - Vassilev, N., A. Medina, R. Azcon and M. Vasslev, 2006. Microbial solubilization of rock phosphate media containing agro-industrial wastes and effect of the resulting products on plant growth and P uptake. Plant Soil, 287: 77-84.
Direct Link - White, T.J., T.D. Bruns, S.B. Lee and J.W. Taylor, 1990. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In: PCR Protocols: A Guide to Methods and Applications, Innis, M.A., D.H. Gelfand, J.J. Sninsky and T.J. White (Eds.), Academic Press, San Diego, CA, USA, ISBN-13: 9780123721808, pp: 315-322.
CrossRefDirect Link - Xiao, C.Q., R.A. Chi, X.H. Huang, W.X. Zhang, G.Z. Qiu and D.Z. Wang, 2008. Optimization for rock phosphate solubilization by phosphate-solubilizing fungi isolated from phosphate mines. Ecol. Eng., 33: 187-193.
CrossRef - Yu, X., X. Liu, T.H. Zhu, G.H. Liu and C. Mao, 2012. Co-inoculation with phosphate-solubilzing and nitrogen-fixing bacteria on solubilization of rock phosphate and their effect on growth promotion and nutrient uptake by walnut. Eur. J. Soil Biol., 50: 112-117.
CrossRef - Zapata, F. and A.R. Zaharah, 2002. Phosphorus availability from phosphate rock and sewage sludge as influenced by the addition of water soluble phosphate fertilizer. Nutr. Cycling Agroecosyst., 63: 43-48.
CrossRef