
Asad Ud-Daula
Helmholtz Zentrum M�nchen, German Research Center for Environmental Health, Institute of Ecological Chemistry, Ingolst�dter Landstrasse 1, D-85764 Neuherberg, Germany
Gerd Pfister
Helmholtz Zentrum M�nchen, German Research Center for Environmental Health, Institute of Ecological Chemistry, Ingolst�dter Landstrasse 1, D-85764 Neuherberg, Germany
Karl-Werner Schramm
Helmholtz Zentrum M�nchen, German Research Center for Environmental Health, Institute of Ecological Chemistry, Ingolst�dter Landstrasse 1, D-85764 Neuherberg, Germany
Pakistan Journal of Biological Sciences
Year: 2012 | Volume: 15 | Issue: 23 | Page No.: 1133-1138







ABSTRACT
Two types dopamine receptor present in the cell membrane of vertebrates. But in this study D1 receptor was identified in the invertebrate ciliates protozoan, Tetrahymena thermophila by use of fluorescent ligands. D1 specific agonist SKF-38393 binds specifically to Tetrahymena. The specific binding of SKF-38393 was encountered by equimolar addition of D1 antagonist thus showed no binding of ligands. In addition, it was also proved that the D1 specific agonist did not cross bind with the D2 type receptor due to the equimolar addition of D2 selective antagonist spiperone. Interestingly this study also showed that the dopamine receptor present in the endoplasmic reticulum and endosomes of Tetrahymena as well as cell membrane which was revealed by laser scanning microscope. Therefore, this evidence supports the existence of a D1 receptor in the ciliate protozoan.
PDF Abstract XML References Citation
Received: September 27, 2012;
Accepted: March 04, 2013;
Published: March 09, 2013
How to cite this article
Asad Ud-Daula, Gerd Pfister and Karl-Werner Schramm, 2012. Identification of Dopamine Receptor in Tetrahymena thermophila by Fluorescent Ligands. Pakistan Journal of Biological Sciences, 15: 1133-1138.
DOI: 10.3923/pjbs.2012.1133.1138
URL: https://scialert.net/abstract/?doi=pjbs.2012.1133.1138
DOI: 10.3923/pjbs.2012.1133.1138
URL: https://scialert.net/abstract/?doi=pjbs.2012.1133.1138
INTRODUCTION
In vertebrates, two distinct populations of dopamine receptors exist in the nervous system as D1 type and D2 type. The physiological effects of dopamine are mediated by at least five G protein-coupled receptor subtypes encoded by different genes referred to as D1-D5 (O’Dowd, 1993). The D1 receptor is positively linked to the activation of the cAMP and D2 receptor is linked to a variety of signal transduction mechanisms (Enjalbert et al., 1986; Lacey et al., 1987). The pharmacological profiles of these two dopaminergic receptor can be discriminated on the basis of their agonist/antagonist specificities (Creese et al., 1983). Fluorescent dopaminergic agonist/antagonist ligands play a vital role to the recognition of cellular localization of these two dopamine receptors (Ariano et al., 1989). SKF-38393 agonist is high affinity D1 receptor selective ligand (Iorio et al., 1983) and SCH 23390 is an antagonist ligand to D1 receptor (Kaiser et al., 1982). Both ligands were chemically derivatized with fluorescent com.
MATERIALS AND METHODS
Chemicals: D1 selective receptor agonist SKF-38393, antagonist SCH 23390 and D2 selective antagonist spiperone were purchased from Sigma-Aldrich Chemie GmbH Laboratory, Germany. Both ligands were derivatized by fluorescent rhodamine-dye (Monsma et al., 1989). The stock solutions of these compounds were prepared in Millipore water according to the solubility instruction of chemicals by manufacturers.
Equipments: Protozoan cell inoculation was maintained inside a sterile bank (Heraeus LaminAir® HBB 2448 S) and incubated in an incubator (Heraeus kelvitro®t). The experiment was conducted in sterile tissue culture plates, 96 W and flat bottom obtained from Greiner bio-one, Germany. Millipore water was prepared by Milli-Q® Ultrapure Water Purification Systems (290 Concord Road Billerica, MA, 01821, USA). The binding of dopamine ligands to the cell was visualized by the two different fluorescence microscope, one was Axioplan 2 and Camera, AxioCam MRm (Zeiss, Germany) and the other was the laser scanning microscope, LSM 510 META (Carl Zeiss, Germany).
Protozoan cell and culture medium: The ciliated unicellular protozoan Tetrahymena thermophila SB210 was received from Tetrahymena Stock Center, C5 155 Veterinary Medical Center, Cornell University, Ithaca, New York 14853. It was grown axenically at 28°C in a nutrient Proteose Peptone Yeast (PPY) extract medium (Ud-Daula et al., 2008).
Exposure of dopamine ligands to the cell: The experiment was conducted in 96 well plates and 250 μL inoculums was taken in each well of the plate and consequently incubated at 28°C. Initial concentration of the cells for the experiment was maintained at 1x104 cells mL-1 according to OECD guide lines. Ligands were added in the well plates at molar concentration where the selected five mounted concentrations were 5, 10, 20, 40 and 80 μM. The rhodamine-labeled dopamine receptor agonists SKF-38393 was exposed to the Tetrahymena cells and incubated for two different times, 3 and 6 h. After incubation, 200 μL cell-suspension was taken into eppendorf tubes followed by fixation with 10% formaldehyde and washed twice with Phosphate Buffer Saline (PBS). Then 6 μL cell pellets were used to prepare slides and subsequently the fluorescent signal was examined employing fluorescence microscopy. Just prior to examination, the slides were covered with cover slips and a drop of immersion oil was added on the top. Then, cells were observed using the fluorescence optics of a Zeiss photomicroscope-2 with 546/590 nm excitation/emission filters for the rhodamine fluorophore. Two-dimensional photomicrographs were captured by an auto camera with 40xmagnification. Six hours incubation of ligands elicited the higher fluorescent activity. The specific binding of the fluorescent probe to dopamine receptors was assessed with competition between agonist SKF-38393 and D1 specific antagonist SCH 23390. Both ligands were labelled with rhodamine and added together to the cells at the same five nominal concentrations; 5 μM, 10, 20, 40 and 80 μM and incubated for 6 hours. The cross binding of SKF-38393 to the receptors was faithfully checked by spiperone, a D2 antagonist. These two ligands were exposed to the cells at same concentrations as above mentioned and incubated for 6 h. Afterwards the binding of SKF-38393 to D1 receptor was scrutinized to determine competitive cross-reactivity of ligands binding in the cells.
The 40 μM concentration of SKF-38393 exhibited the higher binding sensitivity to D1 receptor after 6 h incubation. This binding of SKF-38393 was used for further assessment employing Laser Scanning Microscope (LSM) where argon ion laser 488 nm excitation and Band Pass (BP) filter 500-550 nm emission were used. Image was magnified 40 times by C-Apochromat Water Immersion Lens. (Helium Neon Laser 543 nm excitation and LP 560 emission). The LSM capture photograph from a different plane; thus indicates the binding location of agonist in the cell. Slide preparation carried out as above mentioned.
RESULTS AND DISCUSSION
Rhodamine SKF-38393 agonist and rhodamine SCH-23390 antagonist fluorescent probes demonstrated faithfully the cellular localization of the D1 dopamine receptor subtype. The investigation of D1 type receptor in Tetrahymena thermophila exerted photographic representation of ligands binding to the receptor. Figure 1 illustrates the photograph of control cells. The control cells were also observed by 546/590 nm excitation/emission for the rhodamine fluorophore. They did not show fluorescent signals because they were not exposed with SKF-38393.
In this experiment, the exposure of cells to fluorescent agonist SKF-38393 caused the emission of fluorescent signals. The cells were treated with several concentrations of the agonist at two different incubation times. 3 h incubation demonstrated comparatively weaker signals than 6 h incubation as illustrated in Fig. 2. Longer incubation causes saturation of receptor binding. But, more than 8 h exposure of ligand again caused the decrease of receptor binding as well as decrease of fluorescent signals. The specific binding of the ligands was investigated with exposure to two ligands; one was agonist SKF-38393 and another was antagonist SCH-38393. Both ligands are specific to D1 receptors and were added together at the same concentration. The exposed cells were incubated for 6 h and viewed by employing fluorescence microscopy.
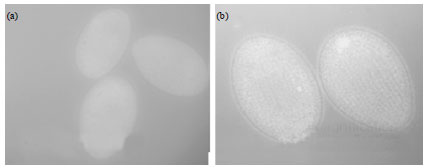 | |
| Fig. 1(a-b): | Control Tetrahymena thermophila cell (a) 40xMagnification (b) 100xMagnification |
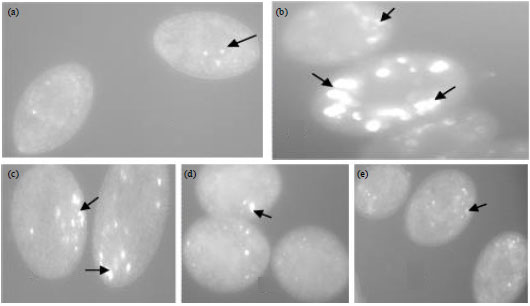 | |
| Fig. 2(a-e): | Fluorescence localization of D1 dopamine receptor binding of the D1 specific agonist fluorescent probe, rhodamine- SKF 38393 in Tetrahymena thermophila. The entire photograph shows total binding of receptor after 3 h exposure of ligand. This time period causes saturation binding of ligands and reveals stronger signals. Ligand concentration, (a) 5 μM, (b) 10 μM, (c) 20 μM, (d) 40 μM and (e) 80 MM arrows: Binding of SKF-38393 |
 | |
| Fig. 3(a-e): | Experiment for specific binding of agonist SKF 38393 to D1 receptor in the Tetrahymena cell. SKF 38393 and D1 specific fluorescent probe antagonist SCH 23390 were applied together to the cells at equal five different nominal concentrations and incubated for 3 h. Cells did not show fluorescence signals of receptor binding for neither agonist nor antagonist. Thus prove the specific binding of SKF 38393. Ligand concentrations of (a) 5 μM, (b) 10 μM, (c) 20 μM, (d) 40 μM and (e) 80 μM, respectively |
The affinities of the two ligands were encountered by each other and exhibited no fluorescence signals. Photographs are presented in the Fig. 3. Interestingly the same result was found when, that the antagonist of one type dopamine receptor could SKF-38393 and SCH-23390 were used together in guinea-pig not prevent the binding of another type dopamine agonist (Furukawa and Morishita, 1997; Zhu et al., 2000).
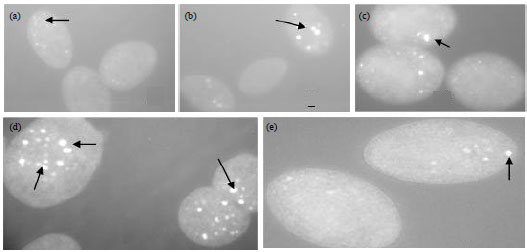 | |
| Fig. 4(a-e): | Assay for the cross binding to D1 receptors. SKF 38393 and non-fluorescent D2 specific antagonist, spiperone were added together to the cells at the equal five different nominal concentrations and incubated for 3 h. Spiperone competes only to binding to D2 tape receptors. Cells exhibited fluorescent signals of receptor binding. So, spiperone did not inhibit agonist binding. This indicates SKF-38393 agonist binding specifically to D1 receptors but not to D2 receptors. Ligand concentrations unit with each No. of (a) 5 μM, (b) 10 μM, (c) 20 μM, (d) 40 μM and (e) 80 μM, respectively. Arrows: Binding of SKF -38393 |
It has also been previously reported that two types of dopamine receptors exists in dopaminergic cells. Therefore, the assessment of cross-affinity of receptor binding is also important. That is why we have selected the D2 specific receptor antagonist spiperone which compete binding to D2 receptor due to its ligand properties (Amenta et al., 1999). SKF-38393 and spiperone were added to the cells together with the five different concentrations of ligand again and incubated for 6 hours. Spiperone did not inhibit SKF-38393 binding to the D1 receptor; photographs are shown in Fig. 4.
It has been previously reported that the affinity of SKF-38393 D1 receptor agonist is inhibited by the selective D1 antagonist SCH-23390, but not by the D2 specific antagonist (Ongini et al., 1985). The fluorescent agonist SKF-38393 binds specifically to D1 type receptors in Tetrahymena thermophila. Its binding to the receptor increases with the exposure time, thus saturating the binding capacity of the D1 receptor and increasing the fluorescence signal. But, longer exposure time of ligands over 8 h causes down regulation of the number of receptors and consequently reduces fluorescence signal. Because prolonged or repeated exposure to a stimulus often results in decreased responsiveness of that receptor to a stimulus thus effecting receptor desensitization and altered affinity for the ligand (Sun et al., 2003). Five different nominal concentrations of the agonist were applied for binding to the D1 receptor. Six hours incubation for all of the applied concentrations of the agonist revealed stronger signals wherein 20 and 40 μM concentrations displayed bright signals. The affinity of agonist binding to the D1 receptors was blocked or damped due to the use of antagonist SCH-23390 at same molar concentrations. Both ligands undergo binding competition for the D1 receptor and antagonize their affinity by each other; as a result the cells did not show ligand binding. Many publications reported that the antagonist of one type of dopamine receptor can not prevent the binding of another type of dopamine agonist. Thus, the D2 specific receptor antagonist spiperone was used to resolve the dispute of cross-binding of SKF-38393 agonist to the D1 receptor. When spiperone and SKF-38393 together were exposed to the cells then the agonist still binds to the D1 receptor. Therefore, D2 receptor antagonist do not compete binding with D1 receptor.
Furthermore, the binding of SKF-38393 to D1 type receptor in T. thermophila cell was clearly demonstrated by capturing photograph with employing Laser Scanning Microscope (LSM). Only one suitable concentration, 40 μM of SKF-38393 was used for this study. The transparent picture (Fig. 5) of the cell exhibited a brighter signal in the cytoplasm than in the membrane site. Therefore, it is revealed that the receptor is densely organized and located in the cytosol. This photograph also showed that then D1 receptors are abundant in endosome and rough Endoplasmic Reticulum (ER). Rough ER remains bounded to the ribosome where the proteins are synthesized and migrated to Golgi bodies through transport vesicule.
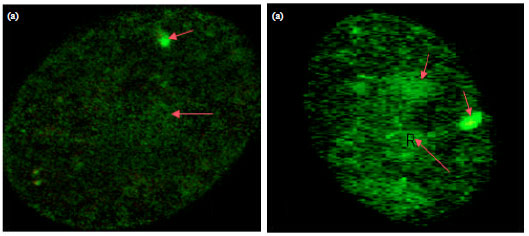 | |
| Fig. 5(a-b): | Both of the figures are captured by LSM. (a) Normal and (b) Transparent picture of same cell. The transparent picture indicated that D1 receptor is abundant in the endosome and rough ER rather than cell membrane. This receptor is internalized in the endosome may be caused by the agonist stimulation. R: Ribosomal location of D1 type receptor, Arrows: Binding of SKF-38393 both in the membrane and endosome |
G protein Coupled Receptor (GPCR) mediates the role of D1 receptors. According to the current ideas about GPCR trafficking in general (Dohlman et al., 1991), those dopamine receptors in particular are internalized in early endosome from which they may be recycled to the plasma membrane or targeted for turnover in late endosome (Ng et al., 1994). A number of transfection studies of D1 and D2 have shown a predominantly cytoplasmic localization for D2 (Ng et al., 1994; Vickery and von Zastrow, 1999; Prou et al., 2001). Surprisingly, agonist stimulation, results in internalization of D1 (Ng et al., 1994; Vickery and von Zastrow, 1999). Internalization of the receptor following ligands binding is known to occur for a number of GPCRs, including the D1 type receptors and this leads to a lowering of surface receptor density (Sun et al., 2003) which are not visible in this observation. However, this observation demonstrated that D1receptor is more visible in the other area of the cell e.g. rough ER. This cell organelle contains ribosome which produces all kind of proteins including D1 receptor protein. The observation was also claimed that D1 receptor present around the rough ER in addition with transport vesicles. This investigation indicated only a D1 type receptor. But dopaminergic cells contain both D1 and D2 type receptors together. Therefore, D2 receptor may also be present in the Tetrahymena cell.
CONCLUSION
D1 type dopamine receptor is identified in the ciliate protozoan Tetrahymena thermophila using fluorescent ligands employing microscopic studies. Both fluorescent and laser scanning microscope were demonstrated that D1 receptor is present in the cell membrane, rough endoplasmic reticulum and endosome. The model invertebrate organisms such as Caenorhabditis elegans, Drosophila melanogaster and Apis mellifera contains D1nd D2 type receptor and similarly single cell ciliates, Tetrahymena thermophila showed D1 receptor and may also D2 receptor. Therefore, Tetrahymena thermophila may join to these groups of invertebrates as a model organism for dopamine receptor research as well as for the understanding of other neurotransmitters receptors mechanisms.
ACKNOWLEDGMENT
This research was completely done and financed by German Research Center for Environmental Health, D-85764 Neuherberg, Germany.
REFERENCES
- Amenta, F., E. Bronzetti, L. Felici, A. Ricci and S.K. Tayebati, 1999. Dopamine D2-like receptors on human peripheral blood lymphocytes: A radioligand binding assay and immunocytochemical study. J. Auton. Pharmacol., 19: 151-159.
PubMed - Ariano, M.A., F.J. Jr Monsma, A.C. Barton, H.C. Kang, R.P. Haugland and D.R. Sibley, 1989. Direct visualization and cellular localization of D1 and D2 dopamine receptors in rat forebrain by use of fluorescent ligands. Proc. Nat. Acad. Sci., 86: 8570-8574.
PubMed - Creese, I., D.R. Sibley, M.W. Hamblin and S.E. Leff, 1983. The classification of dopamine receptors: Relationship to radioligand binding. Annu. Rev. Neurosci., 6: 43-71.
PubMed - Dohlman, H.G., J. Thorner, M.G. Caron and R.J. Lefkowitz, 1991. Model systems for the study of seven transmembrane-segment receptors. Annu. Rev. Biochem., 60: 653-688.
PubMed - Enjalbert, A., F. Sladczed, G. Guillon, P. Bertrand and C. Shu et al., 1986. Angiotensin II and dopamine modulate both cAMP and inositol phosphate productions in anterior pituitary cells, involvement in prolactin secretion. J. Biol. Chem., 261: 4071-4075.
PubMed - Furukawa, T. and H. Morishita, 1997. Existence of dopamine D1 receptor on the sympathetic nerve endings in the guinea-pig vas deferens. Eur. J. Pharmacol., 328: 229-234.
PubMed - Iorio, L.C., A. Barnett, F.H. Leitz, V.P. Houser and C.A. Korduba, 1983. SCH 23390 a potential benzazepine antipsychotic with unique interactions on dopaminergic systems. J. Pharmacol. Exp. Ther., 226: 462-468.
Direct Link - Kaiser, C., P.A. Dandridge, E. Garvey, R.A. Hahn and H.M. Sarau et al., 1982. Absolute stereochemistry and dopaminergic activity of enantiomers of 2,3,4,5-tetrahydro-7,8-dihydroxy-1-phenyl-1H-benzazepine. J. Med. Chem., 25: 697-703.
PubMed - Lacey, M.G., N.B. Mercuri and R.A. North, 1987. Dopamine acts on D2 receptors to increase potassium conductance in neurones of the rat substantia nigra zona compacta. J. Physiol., 392: 397-416.
PubMed - Monsma, F.J., A.C. Barton, H.C. Kang, D.L. Brassard, R.P. Haugland and D.R. Sibley, 1989. Characterization of novel fluorescent ligands with high affinity for D1 and D2 dopaminergic receptors. J. Neurochem., 52: 1641-1644.
PubMed - Ng, G.Y., B. Mouillac, S.R. George, M. Caron, M. Dennis, M. Bouvier and B.F. O'Dowd, 1994. Desensitization, phosphorylation and palmitoylation of the human dopamine D1 receptor. Eur. J. Pharmacol., 267: 7-19.
PubMed - Ongini, E., M.G. Caporali and M. Massotti, 1985. Stimulation of dopamine D-1 receptors by SKF 38393 induces EEG desynchronization and behavioral arousal. Life Sci., 37: 2327-2333.
PubMed - Prou, D., W.J. Gu, L.S. Crom, J.D. Vincent, J. Salamero and P. Vernier, 2001. Intracellular retention of the two isoforms of the D(2) dopamine receptor promotes endoplasmic reticulum disruption. J. Cell Sci., 114: 3517-3527.
PubMed - Sun, W., N. Ginovart, F. Ko, P. Seeman and S. Kapur, 2003. In vivo evidence for dopamine mediated internalization of D2-receptors after amphetamine: Differential findings with [3H] raclopride versus [3H]spiperone. Mol. Pharmacol., 63: 456-462.
PubMed - Ud-Daula, A., G. Pfister and K.W. Schramm, 2008. Growth inhibition and biodegradation of catecholamines in the ciliated protozoan Tetrahymena pyriformis. J. Environ. Sci. Health Tox Hazard Subst. Environ. Eng., 43: 1610-1617.
PubMed - Zhu, Z.T., Y. Fu, G.Y. Hu and G.Z. Jin, 2000. Electrophysiological study on biphasic firing activity elicited by D(1) agonistic-D(2) antagonistic action of (-)-stepholidine in nucleus accumbens. Sheng Li Xue Bao., 52: 123-130.
PubMed