
M. Rabeeth
Chikkanna Government Arts College, Tiruppur-602, Tamil Nadu, India
A. Anitha
Nehru Arts and Science College, Coimbatore-105, Tamil Nadu, India
Geetha Srikanth
Amrita Vishwa Vidyapeetham University, Coimbatore, Tamil Nadu, India
Pakistan Journal of Biological Sciences
Year: 2011 | Volume: 14 | Issue: 16 | Page No.: 788-797







ABSTRACT
Streptomyces griseus (MTCC 9723) is a chitinolytic bacterium isolated from prawn cultivated pond soil of Peddapuram Village; East Godavari District was studied in detailed. Chitinase (EC 3.2.1.14) was extracted from the culture filtrate of Streptomyces griseus and purified by ammonium sulfate precipitation, DEAE-cellulose ionexchange chromatography, Sephadex G-100 and Sephadex G-200 gel filtration chromatography. The molecular mass of the purified chitinase was estimated to be 34, 32 kDa by SDS gel electrophoresis and confirmed by activity staining with Calcofluor White M2R. Chitinase was optimally active at pH of 6.0 and at 40°C. The enzyme was stable from pH 5-9 and up to 20-50°C. The chitinase exhibited Km and Vmax values of 400 mg and 180 IU mL-1 for colloidal chitin. Among the metals and inhibitors that were tested, the Hg+, Hg2+ and P-chloromercuribenzoic acid completely inhibited the chitinase activity at 1 mM concentration. The purified chitinase showed high activity on colloidal chitin, chitodiose, and chitooligosaccharide. An in vitro assay proved that the crude chitinase, actively growing cells of S. griseus having antifungal activity against all studied fungal pathogen. This result implies that characteristics of S. griseus producing endochitinase made them suitable for biotechnological purpose such as for degradation of chitin containing waste and it might be a promising biocontrol agent for plant pathogens.
PDF Abstract XML References Citation
Received: July 05, 2011;
Accepted: October 14, 2011;
Published: November 24, 2011
How to cite this article
M. Rabeeth, A. Anitha and Geetha Srikanth, 2011. Purification of an Antifungal Endochitinase from a Potential Biocontrol Agent Streptomyces griseus. Pakistan Journal of Biological Sciences, 14: 788-797.
DOI: 10.3923/pjbs.2011.788.797
URL: https://scialert.net/abstract/?doi=pjbs.2011.788.797
DOI: 10.3923/pjbs.2011.788.797
URL: https://scialert.net/abstract/?doi=pjbs.2011.788.797
INTRODUCTION
Chitin, is a homopolymer of β-(1, 4)-linked N-acetyl-D-glucosamine (GlcNAc), is the second most abundant important sources of nutrients and energy polysaccharide existing in nature after cellulose. Chitin is a major structural component of most biological systems such as insects, crustaceans, fungi, algae, protozoa and marine invertebrates (Gomes et al., 2001). Degradation of chitin is performed by three enzymes categorized as endochitinase, exochitinase and β-N acetylglucosaminidase that constitute the chitinase complex. Chitinase (EC 3.2.1.14) is the first enzyme acts on the insoluble chitin which produces a multimer of N-acetylglucosamine (NAG), the exochitinase which produce a soluble dimmer of NAG and then β-N acetylglucosaminidase (EC 3.2.1.52) produces a single NAG from dimeric unit (Vyas and Deshpande, 1989).
Chitinases are widely distributed in nature and play an important role in the degradation of chitin. Chitinases are present in a wide range of organisms, including organisms that do not contain chitin, such as bacteria including streptomyces, fungi, viruses, higher plants and animals and play important physiological and ecological roles.
Actinomycetes, particularly streptomycetes, which are Gram-positive mycelial bacteria, ubiquitous in soil, are well known as producers of many extracellular enzymes with polymer-degrading properties, including chitinases (Gomes et al., 2000; Gupta et al., 1995). During the last decade, chitinases have received increased attention due to their potential application in biocontrol of phytopathogenic fungi (Taechowisan et al., 2003). The organism could either be used directly in the biological control of microorganisms, or indirectly using purified proteins. This study deals with the extraction and purification of endochitinase from Streptomyces griseus (MTCC - 9723), for their possible use as biocontrol agents against phytopathogenic fungi.
MATERIALS AND METHODS
Chitinase enzyme producing S. griseus (MTCC - 9723) stain was isolated from prawn cultivated pond soil of Peddapuram Village; East Godavari District. Further the purification and in vitro antifungal activity of enzyme chitinase of S. griseus was studied in detail.
Inoculum preparation
Primary inoculum: The isolated and identified Streptomyces griseus (AC11) from prawn cultivated soil was inoculated into 100 mL Yeast Mannitol (YM) broth, incubated in a rotary shaker for 24-48 h at 37°C (Remi-R8C).
Secondary inoculum: One mL of primary inoculum was inoculated into 100 mL of Yeast Mannitol broth, incubated in a rotary shaker for 36-48 h at 37°C.
Growth and enzyme production: One mL of secondary inoculum of isolated S. griseus (AC11) was cultured in chitinase production medium (MS medium) containing the following (g L-1); 1 g colloidal chitin, 5 g peptone, 5 g Yeast, 0.7 g KH2PO4, 0.3 g K2HPO4, 4 g NaCl, 5 g MgSO4•7H2O, 1 mg FeSO4•7H2O, 0.1 mg ZnSO4•7H2O, 0.1 mg MnSO4•7H2O, 1 g NH3SO4, pH 7.0 and at 37°C for 5-7 days. The culture was harvested, filtered, centrifuged and the supernatant was used for further studies (Kim et al., 2003).
Protein estimation: The protein content of the enzyme source was estimated as per Lowry et al. (1951).
Endo chitinase assay: Colloidal chitin was used as a substrate with reference to Wen et al. (2002). The assay mixture containing 0.3 mL of 1% colloidal chitin in acetate buffer (50 mM; pH 6.0) and 1 mL of enzyme source, incubated at 30°C for 30 min. The hydrolysis reaction was terminated by adding 0.6 mL of Dinitrosalicylic acid (DNS) and kept in a boiled water bath for 15 min and centrifuged at 10,000 rpm for 10 min. The amount of reducing sugar released in the supernatant was measured at 540 nm (Miller, 1959) using N-acetyl D-glucosamine as standard. One unit of enzyme activity was defined as the amount of enzyme that catalyzed the release of 1 μmol of N-acetyl D-glucosamine/mL/min.
Enzyme purification: The purification of chitinase was carried out in four steps. The protein from the crude enzyme was precipitated by ammonium sulphate (75%) at 4°C overnight. The precipitate was collected by centrifugation and resuspended in an acetate buffer (50 mM; pH 5.0). It was dialyzed against same buffer and freeze-dried.
The dialyzed freeze dried sample was reconstituted and loaded on a DEAE-cellulose column (2x20 cm) pre equilibrated with acetate buffer (50 mM; pH 6.0) and washed with same buffer. The enzymes were eluted in a stepwise gradient on NaCl (0.1 – 1.0 M) at a flow rate of 24 mL h-1. Fractions of 3 mL were collected and read at 280 nm. The fractions containing chitinase activity were combined, concentrated by lyophilization and stored at -20°C for further use.
The concentrated sample was passed through Sephadex G-100 column (Hidex Himedia; 60x1.0 cm column) for further purification. The column was packed and equilibrated with acetate buffer (50 mM; pH 6.0) and eluted with the same buffer at flow rate of 15 mL h-1. The fractions of 3 mL were collected, proteins were measured at 280 nm (Systronics; 2101) and chitinase activity was measured at 540 nm. The fractions containing enzyme activity were pooled and freeze dried.
Further purification of freeze dried sample was done by passing through Sephadex G-200 column (Hidex Himedia; 50x2.5 cm). Fractions of 3 mL were collected with a flow rate of 15 mL h-1, proteins were measured at 280 nm. The activity containing fractions were pooled; freeze dried, stored at- 20°C for further use (Shin-Hye and Jung-Hyun, 2000).
Protein profiling by sodium dodecyl sulphate- polyacrylamide gel electrophoresis (SDS-PAGE): From each purification step, protein purity and molecular weight were characterized by SDS-PAGE using 10% gel (Laemmli, 1970).
Zymography analysis: Glycol chitin was synthesized by acetylation of glycol chitosan as per the procedure stated by Trudel and Asselin (1989).
Native PAGE was prepared with 10% separating gel and 5% stacking gel containing 0.05% glycol chitin. After electrophoresis at 4°C, the gel was stained with 0.01% Calcoflour white M2R in Tris-HCl buffer (0.5 M; pH 8.9) for 7 min and destained with water. The lytic zones were photographed under UV-transilluminator (Kim et al., 2003; Gohel et al., 2005). The active bands in the native gel were eluted by homogenization. The contents were centrifuged from the collected supernatant protein content and chitinase activity was measured. The molecular weight of the eluted fractions was determined by SDS-PAGE (Laemmli, 1970).
Effect of various factors on chitinase activity and stability: The parameters such as optimum substrate concentration (Vmax and Km), optimum pH and temperature were measured.
Effect of substrate concentration: Different concentrations of substrate colloidal chitin (0.2-1.4 g) were incubated with purified enzyme preparation. The enzyme activity was measured and Vmax, Km were determined by Line Weaver- Burk’s Plot (Dahiya et al., 2005).
Effect of pH: The effect of pH on chitinase activity was determined by incubating the enzyme chitinase at different pH levels (pH 3-12) under standard assay condition. The buffer system used is as follows: Glycine HCL buffer (50 mM), pH 3.0; Acetate buffer (50 mM), pH 4-5; Phosphate buffer (50 mM), pH 6.0-7.0; Tris -HCl buffer (50 mM), pH 8.0- 9.0 and Glycine - NaOH buffer (50 mM), pH 10.0-12.0 was used. The enzyme stability was determined by pre incubating the enzyme at various pH (pH 3-12) without substrate for 16 h. The residual enzyme activity was measured under standard assay condition.
Effect of temperature: Chitinase activity was determined by performing the standard assay at various temperature ranges from 20 to 80°C. To determine thermal stability, enzyme was incubated for 1 h at the temperature of 20 to 80°C without substrate. At the end, the residual chitinase activity was measured under standard assay condition.
Activity of enzyme chitinase on different substrates: The enzyme chitinase was incubated separately with different substrates like powdered chitin, colloidal chitin, swollen chitin, chitobiose, chitotetrose and chitopentose for determining the suitable substrate for enzyme chitinase under standard assay method.
Effect of metal ions and inhibitors on enzyme chitinase activity: The effect of metal ions such as HgNO3, HgCl2, CaCl2. 2H2O, MgCl2, AgNO3, FeCl3, MnCl2, CuSO4, FeSO4, CoCl2. 2H2O and SnCl2 and inhibitors such as KCl, SDS, β-mercaptoethanol, P-chloromecuribenzoate (P-CMB) and EDTA on enzyme activity was studied by incorporating these metal ions and inhibitors at the concentration of 1, 5, 10 and 50 mM. The reaction mixture was pre incubated for half an hour at room temperature and subsequently the residual enzyme activity was measured under standard condition (Dahiya et al., 2005).
Analysis of hydrolysis pattern of chitinase in TLC: Hydrolysis products of colloidal chitin and N-acetyl-chitooligosaccharides (dimer to hexamer) were analyzed by Thin Layer Chromatography (TLC). The purified chitinase of the isolate S. griseus (0.5 U) was incubated with 1 mg of colloidal chitin and N-acetylchitiooligosaccharides (Sigma) in 1 mL of citrate phosphate buffer (0.1 M; pH 6.0) at 37°C for 6 h. The hydrolysates were spotted onto a Silica gel (TLC) coated plates and developed in a solution of n-butanol:
Methanol: 25% ammonia solution: water (5:4:2:1). The spots were detected using anilinephthalate reagent (Shin-Hye and Jung-Hyun, 2000).
Antagonistic activity of S. griseus: Antagonistic activity was observed directly on YEMA plates, using modification of the hyphal extension inhibition assay (Fenice et al., 1998). Actively growing S. griseus were streaked at one edge and pathogenic fungal strains on the opposite side of YEMA plate (Srinon et al., 2006). After incubation at 28°C for 5-7 days the growth of S. griseus and pathogenic fungal strains was measured and Percent Inhibition (PI) was calculated as:
where, γ0 is the growth of test pathogen (cm) in the absence of antagonist strain; γ is the growth of test pathogen (cm) in the presence of antagonist strain.
In vitro inhibition of fungal growth by enzyme chitinase: A spore suspension of plant fungal pathogens namely AFC1-Fusarium oxysporum; AFC2-Alternaria alternate; AFC3-Rhizoctonia solani; AFC4-Fusarium solani; AFC5-Fusarium oxysporum; AFC6-Alternaria alternate; AFC7-Rhizoctonia solani; AFC8-Fusarium solani; AFC9-Aspergillus flavus and AFC10-Aspergillus flavus were uniformly swabbed on Potato Dextrose Agar plates. Filter paper discs were laid on the seeded plates, into each disk 50 μL of chitinase of S. griseus at the concentration of 5, 50 and 100 U was added. Control was prepared simultaneously by disc soaked in distilled water. The plates were incubated over 5-7 days at 28°C and inhibition of mycelia growth was measured.
RESULTS
Purification and characterization of the enzyme chitinase: In the present effort, Streptomyces griseus (MTCC 9723) having chitiolytic activity was studied in detailed. The purification, characterization and antifungal assay of endochitinase enzyme extracted from S. griseus was carried out in detailed.
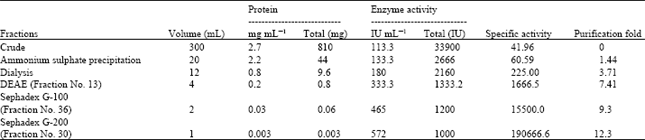
Partial purification of the S. griseus endochitinase enzyme was done by ammonium sulphate precipitation, dialysis, DEAE-Cellulose Ion Exchange Chromatography, Sephadex G-100 and Sephadex G-200 gel filtration chromatographic columns and data’s were illustrated in graphs. Table 1 showed the values specific activity, total protein and purification fold of enzyme chitinase. The selected isolate has shown better growth in MS medium supplemented with colloidal chitin by producing 2.7 mg mL-1 of protein and enzyme activity of 113.3 IU mL-1.
| Table 1: | Partial purification scheme of enzyme chitinase of S. griseus |
 | |
| Values are mean of triplicates | |
The crude enzyme encompasses the specific activity of 41.96 IU mg-1 with purification fold of 0.00. Ammonium sulphate precipitated enzyme showed specific activity of 60.59 IU mg-1, protein content of 2.2 mg mL-1 with purification fold of 1.44. Specific activity of 225, 0.8 mg mL-1 of protein with the purification fold of 3.71 was observed after dialysis. Earlier Kim et al. (2003) revealed that lower specific activity of 14.2 IU mg-1 for chitinase enzyme attained from Streptomyces sp. M-20. In preceding work, lower specific activity of chitinase from Vibrio sp. 98CJ11027(1.3 IU mg-1); Aeromonas schubertii (0.43 U mg-1) and B. subtilis W-118 (2.13 U mg-1) subjected to 30-75% fractional ammonium sulphate precipitation have been reported (Shin-Hye and Jung-Hyun, 2000; Guo et al., 2004; Lan et al., 2006). In contrast, cell free supernatant of Enterobacter sp. NRG4 subjected to 30-75% fractional ammonium sulphate precipitation has exhibited higher chitinase specific activity of 560.5 U mg-1 of protein (71%) with purification fold of 3.18 (Dahiya et al., 2005).
The DEAE- Cellulose Ion Exchange chromatography disclosed a specific activity of 1666.5 IU mg-1 and 0.2 mg mL-1 of protein with the purification fold of 7.41 (Fig. 1). This activity was found to be higher than that of dialyzed sample. Compared with present study, lower specific activity of 30.0 IU mg-1 and 42 mg of protein mL-1 with the purification fold of 4.1 was revealed by Kim et al. (2003).
The enzyme was then purified using Sephadex G-100 column and purification profile was exemplified in Fig. 2. Among the fractions collected, fraction number 36 showed highest specific activity of 15500.0 IU mg-1, protein of 0.06 mg mL-1 with purification fold of 9.3. Whereas, specific activity of 137.0 IU mg-1, protein content of 6 mg mL-1 and the purification fold of 6 for Sephadex G-100 purified chitinase of Streptomyces M-20 reported by Kim et al. (2003). Likewise chitinase of Streptomyces griseus HUT 6037 showed two peaks (P-l and P-2) on Sephadex G-100 column. P-1 showed specific activity of 13.9 IU mg-1, protein of 50.5 mg mL-1 and the purification fold of 0.82, whereas P-2 showed specific activity of 283 IU mg-1, protein of 29.3 mg mL-1 and the purification fold of 61.5 has recalling the findings of present study (Tanabe et al., 2000).
Further purification of enzyme was done by passing the enzyme through Sephadex G-200 gel filtration chromatograpy column (Fig. 3). From the all collected 80 fractions, fraction number 30 showed highest specific activity of 190666.6 IU mg-1. The protein of 0.003 mg mL-1 with purification fold of 12.3 was obtained. On the other hand, Dahiya et al. (2005) have reported the higher purification fold of 44.12 with specific activity of 7783.3 IU mg-1 for Enterobacter sp. NRG4 chitinase using gel filtration. Similarly, according to the study of Shin-Hye and Jung-Hyun (2000) chitinase enzyme of Vibrio sp. purified by Sephadex G -200 column expressed specific activity of 43.0 IU mg-1, protein content of 1.6 mg mL-1 and the purification fold of 33.1 with 27% yield.
The chitinase purified by sephadex G-200 gel filtration showed molecular weight of 34 and 32 kDa by SDS-PAGE (Fig. 4a). Native gel electrophoresis of crude enzyme yield two major bands in zymography analysis which correspond to chitinase activity (Fig. 4b) suggesting that the chitinase of S. griseus having two isozymes. Similar to this study chitinolytic enzyme of Streptomyces sp. TH-11 showed lower molecular weight of 29 kDa (Hoang et al., 2011). In earlier findings, whereas in contrast culture supernatant of S. griseus HUT 6037, showed molecular mass greater than 44 kDa in SDS-PAGE was reported by Tanabe et al. (2000).
Effect of various factors on chitinase activity and stability: Substrate concentration for maximum chitinase activity was determined in terms of Vmax, Km against colloidal chitin as a substrate. The Vmax value was determined by MM equation (Fig. 5) and confirmed by Line Weaver Burk plot (Fig. 6). Vmax of 1 g (180 IU mL-1) and Km of 400 mg was observed which is comparatively lower than the other reports in literature. Higher Km values of Enterobacter sp. NRG4 chitinase of 1.41 mg mL-1 against colloidal chitin was recorded (Dahiya et al., 2005).
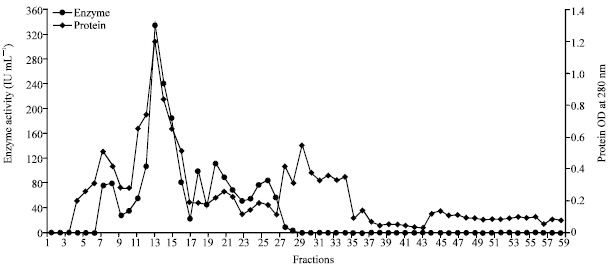 | |
| Fig. 1: | Partial purification of enzyme chitinase of Streptomyces griseus by Ion Exchange Chromatography (DEAE-Cellulose) |
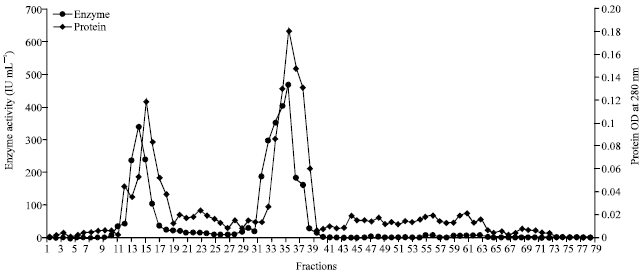 | |
| Fig. 2: | Partial purification of enzyme chitinase of Streptomyces griseus by sephadex G 100 gel chromatography |
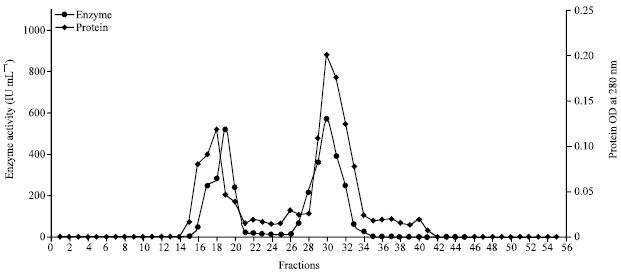 | |
| Fig. 3: | Partial purification of enzyme chitinase of Streptomyces griseus by sephadex G 200 gel chromatography |
Likewise Km of 3.0 mg mL-1 for Alcaligenes xylosoxydans chitinase against colloidal chitin was observed by Vaidya et al. (2003).
The effect of pH on activity and stability of enzyme chitinase of S. griseus was explored. The highest activity of 190±0.00 IU mL-1 was observed at pH 6. The enzyme activity above and below the level of pH 6 was declined. It has been concluded that the optimum pH for enzyme activity is pH 6 (Fig. 7). Many chitinases, including the present one have showed their activity at optimum pH in acidic range.
 | |
| Fig. 4(a-b): | Electrophoretic separation of endochitinase enzyme of S. griseus, Zymographic analysis of protein of S. griseus having endochitinase activity |
The enzyme activity at optimum pH 6 for enzyme chitinase has been reported in the earlier literature (Kim et al., 2003; Yuli et al., 2004).
Enzyme chitinase showed similar activity at the range of 190±0.00 IU mL-1 from pH 5 - pH 9. Further increase in pH, enzyme activity was found to be declined gradually. The above findings confirmed that the stability of enzyme chitinase is between pH of 5 - 9 (Fig. 7). In earlier research, lower pH stability at the range of pH 4-9 for chitinase enzyme preparation obtained from Streptomyces thermoviolaceus OPC-520, Streptomyces viridicans and Streptomyces RC1071 was reported (Tsujibo et al., 1993; Gupta et al., 1995; Gomes et al., 2001). Whereas other bacterial chitinase showed their stability over different rages of pH 6.8-8.0 for Bacillus sp. NCTU2 chitinase (Wen et al., 2002) was scrutinized from the literature.
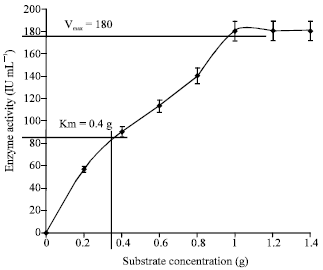 | |
| Fig. 5: | Effect of substrate concentration on activity of enzyme chitinase of S. griseus |
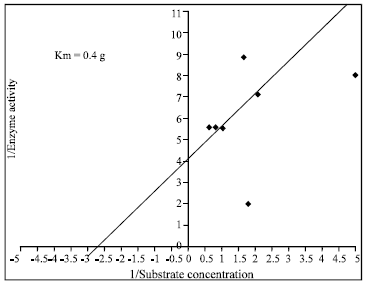 | |
| Fig. 6: | Line Effect of pH on activity and stability of enzyme chitinase of S. griseus |
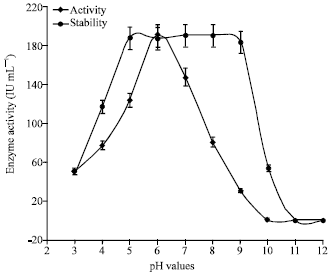 | |
| Fig. 7: | Effect of pH on activity and stability of enzyme chitinase of S. griseus |
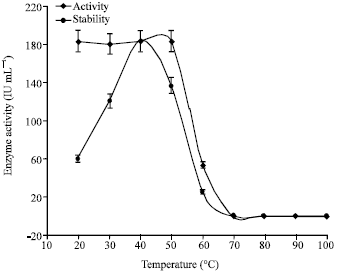 | |
| Fig. 8: | Effect of temperature on activity and stability of enzyme chitinase of S. griseus |
Chitinase of S. griseus showed maximum activity at 40°C (183.3±0.00 IU mL-1). Above and below 40°C the enzyme activity was declined (Fig. 8). Similar activity of 183.3±0.00 IU mL-1 was observed from 20 to 50°C, then activity was declined. This would suggested that the enzyme chitinase from S. griseus is stable upto 50°C (Fig. 8). Wang et al. (2009) observed that optimum temperature of 40°C with stability up to 60°C for chitinase of Bacillus cereus TKU006. Gomes et al. (2001) showed maximum enzyme activity at temperature of 40°C and the stability of 40-70°C for chitinase enzyme from Streptomyces RC1071. In contrast to the present study, Tanabe et al. (2000) have examined the optimum temperature of 60°C and stability upto 40°C for chitinase enzyme of Streptomyces griseus HUT 6037.
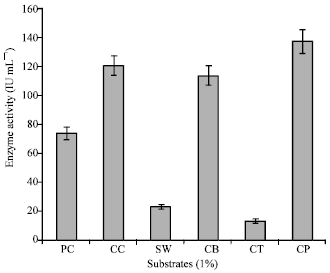 | |
| Fig. 9: | Substrate specificity of enzyme chitinase of S. griseus, PC: Powdered chitin, CC-Colloidal chitin, SC: Swollen Chitin, CB: Chitobiose, CT: Chitotriose, CP: Chitopentose |
More closely associated optimum temperature of 45°C has been exhibited by chitinase enzyme of Vibrio sp. 98CJ11027 (Shin-Hye and Jung-Hyun, 2000). Whereas Jami et al. (2008) reported that chitinase enzyme Paenibacillus chitinolyticus was optimally activity at 37°C.
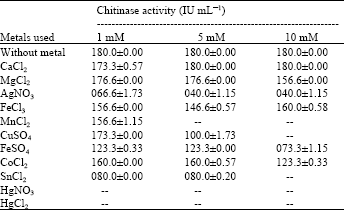
Chitinase activity on different forms of substrates was analyzed. The substrate which is readily hydrolyzed by enzyme chitinase is colloidal chitin (120.3±0.00 IU mL-1) and chitopentose (136.3±0.00). But the enzyme did not hydrolyze chitotriose, swollen chitin and powder chitin (Fig. 9). The above results indicated that chitinase extracted and purified from S. griseus is specific for N-acetyl glucosamine oligomers of more than two units in length. The enzyme chitinase could be classified as endochitinase and N-acetyl-β-glucosaminidase. Like the present result, chitinase enzyme from Aeromonas hydrophila SUWA-9 chitinase has hydrolyzed colloidal chitin, powder chitin and chitosan significantly compared with other substrates (Lan et al., 2006). Similarly, the S. aureofaciens CMUAc130 chitinase showed higher activity against chitooligosaccharides, chitotriose and chitotetraose (Taechowisan et al., 2003). Chitinase activity was strongly inhibited by metal ions of Ag+, Sn+ at 5 mM and completely inhibited by Hg+, Hg2+ at 1 mM, Mn2+ inhibited the enzyme activity at 5 mM where as Sn2+, Cu2+ inhibited the enzyme activity at 10 mM (Table 2). While further tested metal ions did not show any strong inhibitory action on enzyme activity. This indicates the participation of the groups mainly sulphur group present in active site for enzyme activity. Complete inhibition of chitinase activity by Hg+, Hg2+ was reported in many cases (Hiraga et al., 1997; Okazaki et al., 1999; Gomes et al., 2001).
| Table 2: | Effect of metal ions on enzyme chitinase activity of S. griseus |
 | |
| Values are Mean±SE of triplicates | |
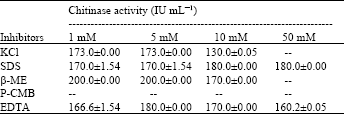
| Table 3: | Effect of chemical compounds and inhibitors on enzyme chitinase activity of S. griseus |
 | |
| Values are Mean±SE of triplicates | |
Besides, Mn2+ and Ca2+ are also shown to inhibit chitinase of Bacillus sp. 13.26 (Yuli et al., 2004) whereas stimulatory effect of Ca2+ (30%) and Mn2+ (20%) at 1 mM concentration on Pseudomonas sp. YHS-A2 chitinase have been suggested by Lee et al. (2000).
The effect of some chemical compounds and inhibitors on chitinase activity was analyzed and showed in Table 3. The increase in chitinase activity of 200±0.00 IU mL-1 at 1 mM of β-mercaptoethanol was observed, whereas P-chloromercuric benzoic acid completely inhibited the enzyme chitinase activity at 1 mM concentration. KCl, SDS and EDTA did not affect the enzyme activity significantly even at 10 mM concentration. Increased activity by β-mercaptoethanol may be indicated by the presence of sulfhydryl groups on active site of the enzyme. There are few reports in the literature on effect of chemical compounds on chitinase activity. Similar inhibition by P-CMB and β-mercaptoethanol augmentation of Streptomyces sp. chitinase was described by Gupta et al. (1995). In contrast to the present work Dahiya et al. (2005) have stated that P-CMB stimulated the chitinase activity of Enterobacter sp. NRG4 and N-bromosuccinamide at 1 mM and iodoacetamide at 10 mM concentration completely inhibited the enzyme activity.
The hydrolysis of colloidal chitin by the enzyme chitinase produced N acetyl glucosamine (GlcNAc) and triacetylchitotriose (GlcNAc3) (Fig. 10a). On treated with N-acetyl-chitooligosaccharides, enzyme chitinase released diacetylchitobiose and tetraacetylchitotetrose (Fig. 10b).
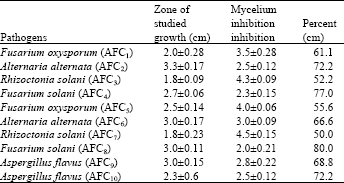
| Table 4: | Anti fungal activity of S. griseus against fungal plant pathogens -Dual culture test |
 | |
| Values are Mean±SE of triplicates | |
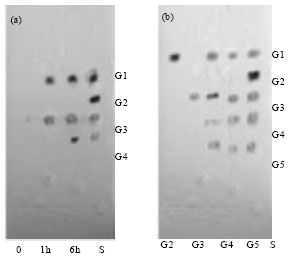 | |
| Fig. 10(a-b): | Tlc of hydrolysis product from (a) colloidal chitin (b) N actyloligosacchaarides of chitinase of S. griseus |
The enzyme reacted on hexa-acetylchitohexose and did not show any reactions on triacetylchitotriose. On the basis of this presumption, S. griseus possessing the enzyme chitinase may be regarded as endochitinase and N-acetyl- β-glucosaminidase and it can be applied in the synthesis of N-acetyl-chitooligosaccharides derivatives. Similar such study was revealed by Li et al. (2002) assumed that the Bacillus brevis No. G1 chitinase having endochitinase activity based on the hydrolysis of a mixture of chitobiose and chitotriose. Jami et al. (2008) despite the fact that, chitooligosaccharides were the predominant products throughout the enzymatic hydrolysis of colloidal chitin showed that the Paenibacillus chitinolyticus chitinase enzyme was an endochitinase.
In the dual culture test, S. griseus showed an inhibitory effect against growth of all plant pathogenic fungal strains. The plate having the isolate of S. griseus showed zone of inhibition of 2.0±0.28 cm and inhibition of 61.1% against growth of F. oxysporum f. sp. lycopersici (FOL) (AFC1), whereas control plate supported an excellent growth of FOL (Table 4).
| Table 5: | Anti fungal activity of S. griseus enzyme chitinase against fungal plant pathogens |
 | |
| Values are Mean±SE of triplicates | |
The results showed that S. griseus producing higher inhibition could be attributed by antibiotics and other enzyme systems such as chitinases, glucanases which is essential for complete cell wall lysis. Similarly in the dual culture, an inhibitory effect of growing Streptomyces RC1071 against growth of fungal pathogens has been clearly observed by Gomes et al. (2000). Yuan and Crawford (1995) observed the in vitro antagonism of Streptomyces WYEC108 against pathogenic fungi of P. ultimum, Aphanomyces euteiches, F. oxysporum, Rhizoctonia solani and Phymatotrichum omnivorum.
The inhibition of plant pathogenic fungal growth was observed on PDA plates with paper discs loaded with crude enzyme chitinase of S. griseus. Among the different plant pathogenic fungi tested, zone of inhibition was found to be 1.5±0.12 cm against FOL (AFC1) at 100 U enzyme concentration (Table 5). Highest zone of inhibition was observed against Alternaria alternata (AFC6) (3.0±0.00 cm) where the control has no effect on fungal growth. The present investigation indicated that chitinase of endophytic S. griseus can be used as a promising biocontrol agent against plant pathogens. Similarly Taechowisan et al. (2003) have observed that purified chitinase from S. aureofaciens CMUAc130 have the ability to inhibit hyphal extension growth of F. oxysporum. Experimented conducted by Gomes et al. (2000) has recalling the present findings. The endochitinase of Streptomyces RC1071 has produced inhibition of growth of Fusarium solani, Fusarium graminearum, Fusarium sp., Aspergillus parasiticus, Fusarium oxisporum and Colletotrichum gloeosporioides pathogenic fungi. Many works in this line have described that chitinases produced by certain microorganisms could take part in the antagonistic process (Von et al., 2003; Prapagdee et al., 2008).
In the present study, in vitro experiments using growing cells via dual culture, crude extract or the purified endochitinase (disk method) were inhibition of fungal growth could be clearly observed. In conclusion purified endochitinse of Streptomyces griseus, actively growing cells has shown a very pronounced activity against phytopathogenic fungi, suggesting potential as biocontrol agents. Further in vivo experiments would be necessary to confirm the in vitro antagonistic activity including green house and field experiments of Streptomyces griseus against plant pathogenic fungi will now be undertaken.
REFERENCES
- Prapagdee, B., C. Kuekulvong and S. Mongkolsuk, 2008. Antifungal potential of extracellular metabolites produced by Streptomyces hygroscopicus against phytopathogenic fungi. Int. J. Biol. Sci., 4: 330-337.
CrossRefPubMedDirect Link - Wen, C.M., C.S. Tseng, C.Y. Cheng and Y.K. Li, 2002. Purification, characterization and cloning of a chitinase from Bacillus sp. NCTU2. Biotechnol. Applied Biochem., 35: 213-219.
PubMed - Fenice, M., L. Selbmann, R. di Giambattista and F. Federici, 1998. Chitinolytic activity at low temperature of an antarctic strain (A3) of Verticillium lecanii. Res. Microbiol., 149: 289-300.
CrossRef - Gomes, R.C., L.T.A.S. Semedo, R.M.A. Soares, L.F. Linhares, C.J. Ulhoa, C.S. Alviano and R.R.R. Coelho, 2001. Purification of a thermostable endochitinase from Streptomyces RC1071 isolated from a cerrado soil and its antagonism against phytopathogenic fungi. J. Applied Microbiol., 90: 653-661.
CrossRef - Gupta, R., R.K. Saxena, P. Chaturvedi and J.S. Virdi, 1995. Chitinase production by Streptomyces viridificans: Its potential in fungal cell wall lysis. J. Applied Bacteriol., 87: 378-383.
PubMedDirect Link - Hiraga, K., L. Shou, M. Kitazawa, S. Takahashi, M. Shimada, R. Sato and K. Oda, 1997. Isolation and characterization of chitinase from a flake-chitin degrading marine bacterium, Aeromonas hydrophila H-2330. Biosci. Biotechnol. Biochem., 61: 174-176.
CrossRefDirect Link - Hoang, K.C., T.H. Lai, C.S. Lin, Y.T. Chen and C.Y. Liau, 2011. The chitinolytic activities of Streptomyces sp. TH-11. Int. J. Mol. Sci., 12: 56-65.
CrossRefDirect Link - Kim, K.J., Y.J. Yang and J.G. Kim, 2003. Purification and characterization of chitinase from Streptomyces sp. M-20. J. Biochem. Mol. Biol., 36: 185-189.
PubMed - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lee, H.S., D.S. Han, S.J. Choi, S.W. Choi, D.S. Kim, D.H. Bai and J.H. Yu, 2000. Purification, characterization and primary structure of a chitinase from Pseudomonas sp. YHS-A2. Applied Microbiol. Biotechnol., 54: 397-405.
PubMed - Li, S., Z.A. Zhao, M. Li, Z.R. Gu, C. Bai and W.D. Huang, 2002. Purification and characterization of a novel chitinase from Bacillus brevis. Acta Biochimica Biophysica Sinca, 34: 690-696.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Dahiya, N., R. Tewari, R.P. Tiwari and G.S. Hoondal, 2005. Chitinase from Enterobacter sp. NRG4: Its purification, characterization and reaction pattern. Electron. J. Biotechnol., 8: 134-145.
Direct Link - Okazaki, K., T. Kawabata, M. Nakano and S. Hayakawa, 1999. Purification and properties of chitinase from Arthrobacter sp. NHB-10. Biosci. Biotechnol. Biochem., 63: 1644-1646.
Direct Link - Wang, S.L., C.H. Chao, T.W. Liang and C.C. Chen, 2009. Purification and characterization of protease and chitinase from Bacillus cereus TKU006 and conversion of marine wastes by these enzymes. Mar. Biotechnol., 11: 334-344.
CrossRef - Guo, S.H., J.K. Chen and W.C. Lee, 2004. Purification and characterization of extracellular chitinase from Aeromonas schubertii. Enzyme Microb. Technol., 35: 550-556.
Direct Link - Shin-Hye, P. and L. Jung-Hyun, 2000. Purification and characterization of chitinase from a marine bacterium. Vibrio sp. 98CJ11027. J. Microbiol., 38: 224-229.
Direct Link - Srinon, W., K. Chuncheen, K. Jirattiwarutkul, K. Soytong and S. Kanokmedhakul, 2006. Efficacies of antagonistic fungi against Fusarium wilt disease of cucumber and tomato and the assay of its enzyme activity. J. Agric. Technol., 2: 191-201.
Direct Link - Taechowisan, T. J.F. Peberdy and S. Lumyong, 2003. Chitinase production by endophytic Streptomyces aureofaciens CMUAc130 and its antagonism against phytopathogenic fungi. Ann. Microbiol., 53: 447-461.
Direct Link - Tanabe, T., T. Kawase, T. Watanabe, Y. Uchida and M. Mitsutomi, 2000. Purification and characterization of a 49-kDa chitinase from Streptomyces griseus HUT 6037. J. Biosci. Bioeng., 89: 27-32.
CrossRef - Tsujibo, H., K. Minoura, K. Miyamoto, H. Endo, M. Moriwaki and Y. Inamori, 1993. Purification and properties of a thermostable chitinase from Streptomyces thermoviolaceus OPC-520. Applied Environ. Microbiol., 59: 620-622.
PubMed - Vaidya, R.J., S.L.A. Macmil, P.R. Vyas and H.S. Chhatpar, 2003. The novel method for isolating chitinolytic bacteria and its application in screening for hyperchitinase producing utant of Alcaligenes xylosoxydans. Lett. Applied Microbiol., 36: 129-134.
CrossRefDirect Link - Gohel, V., P. Vyas and H.S. Chhatpar, 2005. Activity staining method of chitinase on chitin agar plate through polyacrylamide gel electrophoresis. Afr. J. Biotechnol., 4: 87-90.
Direct Link - Von Der Weid, I., D.S. Alviano, A.L.S. Santos, R.M.A. Soares, C.S. Alviano and L. Seldin, 2003. Antimicrobial activity of Paenibacillus peoriae strain NRRL BD-62 against a broad spectrum of phytopathogenic bacteria and fungi. J. Applied Microbiol., 95: 1143-1151.
CrossRef - Vyas, P. and M.V. Deshpande, 1989. Chitinase production by Myrothecium verrucaria and its significant for fungal mycelia degradation. J. Gen. Applied Microbiol., 35: 343-350.
Direct Link - Yuan, W.M. and D.L. Crawford, 1995. Characterization of Streptomyces lydicus WYEC108 as a potential biocontrol agent against fungal root and seed rots. Applied Environ. Microbiol., 8: 3119-3128.
PubMed - Lan, X., X. Zhang, J. Hu and M. Shimosaka, 2006. Cloning, expression and characterization of chitinase from the chitinolytic bacterium Aeromonas hydrophila strain SUWA-9. Biosci. Biotechnol. Biochem., 70: 2437-2442.
PubMed - Jami Al Ahmadi, K., M. Tabatabaei Yazdi, M. Fathi Najafi, A.R. Shahverdi, M.A. Faramarzi, Gh. Zarrini and J. Behravan, 2008. Isolation and characterization of a chitinolytic enzyme-producing microorganism, Paenibacillus chitinolyticus JK2 from Iran. Res. J. Microbiol., 3: 395-404.
CrossRefDirect Link