
A. Zoghlami Khelil
Laboratoire des Productions Animales et Fourrageres, INRAT-2049-Ariana, Tunisie
H. Hassen et S. Benyoussef
Laboratoire des Productions Animales et Fourrageres, INRAT-2049-Ariana, Tunisie
Pakistan Journal of Biological Sciences
Year: 2011 | Volume: 14 | Issue: 12 | Page No.: 693-697







ABSTRACT
Seed germination of two pasture legumes was tested under dark conditions at three alternating temperatures of 5-20, 15-20, 20-35°C and constant 20°C. Duration of the alternating temperatures was 12 h. The seedling counts were checked every day during four weeks. Species differed significantly in germination response to temperature. A. hamosus was more sensitive to temperature than C. scorpioides. The temperature of 20-35°C was more favourable for the germination of A. hamosus than constant 20°C which markedly reduced its germination. The interaction between accessions and temperatures for total germination was significant in both species. In A. hamosus, almost 50% of germination was achieved by all accessions by the first week at alternating 20-35°C temperature while in C. scorpioides, average total germination was significantly low (4%) even by the 28th day. A total rate of 11 and 9% of germination were recorded by two accessions from Ouesslatia (210) and Agareb (19), respectively. A considerable variation for germination was observed among accessions of both species. Temperature requirements for seed germination of the studied species suggest why they are relatively easy or difficult to establish in the field. A. hamosus, with very small seeds required relatively high alternating temperature for germination than C. scorpioides which was insensitive to temperature and has relatively bigger seeds.
PDF Abstract XML References Citation
Received: June 02, 2011;
Accepted: September 12, 2011;
Published: November 17, 2011
How to cite this article
A. Zoghlami Khelil and H. Hassen et S. Benyoussef, 2011. Germination of Astragalus hamosus L. and Coronilla scorpioides (L.) as Influenced by Temperature. Pakistan Journal of Biological Sciences, 14: 693-697.
DOI: 10.3923/pjbs.2011.693.697
URL: https://scialert.net/abstract/?doi=pjbs.2011.693.697
DOI: 10.3923/pjbs.2011.693.697
URL: https://scialert.net/abstract/?doi=pjbs.2011.693.697
INTRODUCTION
Milkvetch (A. hamosus, L.) and Scorpion vetch (C. scorpioides L.) are widely distributed in Tunisia and in other parts of the world (Zoghlami and Zouaghi, 2003). Both these annual legumes are neither well documented nor used as cultivated crops, although their potential for pasture use was recommended since more than fifty years ago (Gounot, 1958). They are often found growing together. A. hamosus is a hard drought-tolerant plant potentially useful for soil protection in Mediterranean areas (Patane and Gresta, 2006).
Legume seeds are known for their rigid seed coat (Kozuharova et al., 2010). In most commercial forage and pasture legumes, seed germination has been a problem for years. Temperature appears to be an important factor for seed germination (Brar et al., 1991). Species which germinate readily over a range of temperatures should be easier to establish than those with highly specific temperature requirements (Towsend and McGinnies, 1972).
In our forage and pasture genetic resources programme we are evaluating species for which there is little information on many characteristics, the most important being temperature requirement for seed germination. The purpose of this study is to determine the response of seed germination of local populations of A. hamosus and C. scorpioides under different temperature treatments, in order to promote a more successful use of these species in the improvement of natural pasture of Tunisia, as well as their integration in crop rotation system.
MATERIALS AND METHODS
Seed germination of A. hamosus and C. scorpioides populations was tested under dark conditions at four temperature treatments: three alternating temperatures of 5-20, 15-25 and 20-35°C and one constant temperature of 20°C. The 20-35°C treatment was not tested on C. scorpioides because of insufficient seeds. Duration of each alternating temperature was 12 h and the regime of moisture was maintained constant between 70 and 80%. A. hamosus and C. scorpioides were represented by 10 and 5 populations, respectively seeds of both species were originated from previous collecting missions conducted conjointly between INRAT, ICARDA and CLIMA/Australia in 1992 and 1994 in order to conserve the local Tunisian forage and pasture legume germplasm.
| Table 1: | Code and characteristics of sites of origin of A. hamosus and C. coronilla populations used in the experiment |
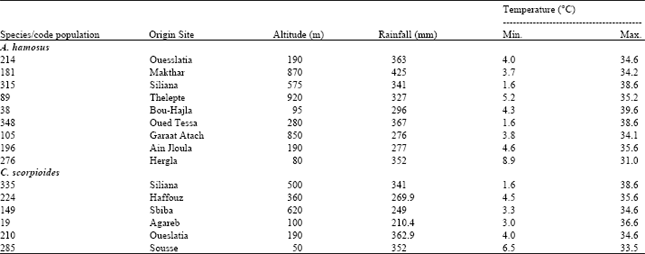 | |
A seed increase programme was then initiated for all germplasm in the subsequent years. However, seeds of tested species were approximately the same age and were stored in room conditions for several years followed by cold storage more recently. The characteristics of the collection sites of the studied populations are presented in Table 1.
For each species and population, one hundred seeds (replicated 3 times) were scarified with sandpaper in order to enhance germination and then placed in Petri dishes on imbibed filter paper. The dishes were placed in a dark germinator at the desired temperature.
Germination counts were made daily over four weeks. For alternating temperatures, heating phase starts each day early in the morning (6 AM) and the cooling phase commences early in the afternoon (18 PM) so as to ensure a duration period of 12 h for each phase.
For both species and for each species separately, analysis of variance was carried out on the angles resulting from the arcsin transformation of the percentages using the SAS version 8 (SAS, 1998). Differences between mean values were evaluated for significance using the Duncan test at 0.05 levels.
RESULTS
For both species together, all main effects and interactions among species, temperatures and accessions were highly significant (p<0.001).
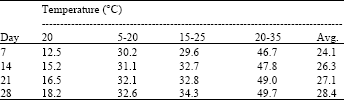
| Table 2: | Seed germination dynamics of A. hamosus populations. Percentage of all germinated seeds during each week at each temperature regime |
 | |
For each species separately, a high significant effect of temperature, accession and their interaction on total germination was observed (p<0.001).
For A. hamosus, the effect of temperature was higher than that of accession or the interaction accession x temperature. However, alternating temperatures (mainly that of 20-35°C) was the most favourable treatment for total germination of this species compared to the constant 20°C which markedly reduced its germination (Table 2). Whatever the temperature, the seed dynamic germination increased with time. The interaction between accession and temperature for total germination was large: 8 accessions were sensitive to the temperature treatment of 20-35° while 2 accessions were sensitive to the temperature treatment of 15-25°C. Forty percent of accessions exceeded 50% of germination at only 20-35°C treatment by the first week. Variation among populations of A. hamosus for total germination was high at all regimes especially 35-20°C. There was only one accession (315) which did not germinate at 20°C and only one accession (348) which did not germinate at 20-35°C regime. Since the germination occurred at all regimes and increased with increasing temperature, A. hamosus can be considered as temperature-tolerant species (Fig. 1a-f). Furthermore, almost all populations germinated better at 20-35°C regime, except 348 and 335 which did not germinate accidently or germinated weakly.
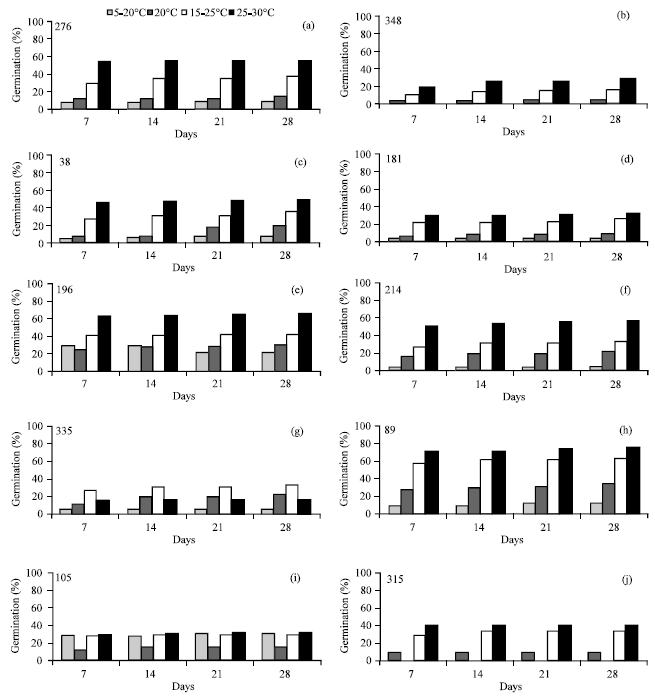 | |
| Fig. 1 (a-j): | Germination of 10 populations (38-89-105-181-196-214-276-315-335-348) of A. hamosus at four time periods and four temperature regimes |
| Table 3: | Average seed germination percentage of C. scorpioides populations at four time periods and four temperature regimes |
 | |
More than 60% of seed germination was recorded at 20-35°C for populations 89 and 196 and slightly for 214. The temperature regime of 5-20°C seemed to be unfavorable for all populations except 105 (Fig. 1). This result did not agree with Brar et al. (1991) who found that 20°C was the recommended temperature for all the tested forage legumes except Mount Barker subterranean clover.
For C. scorpioides, total germination was relatively very low at all regimes and the effect of temperature was not significant (p>0.001) (Table 3).
Variation among accessions was relatively low; the highest total germination didn’t exceed 10% for population’s 210 (Fig. 2a-e).
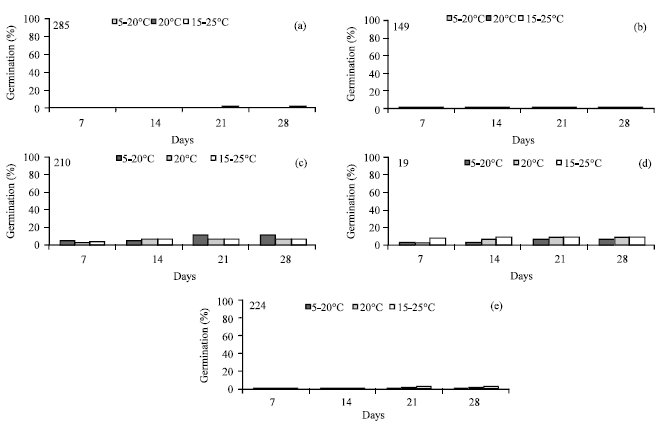 | |
| Fig. 2 (a-e): | Germination of 5 populations (19-149-210-224-285) of C. scorpioides after four time periods and four temperature regimes |
DISCUSSION
Seed germination at fluctuating temperature treatments, 5-20, 15-20, 20-35 and constant 20°C temperature was tested for two annual pasture legumes from the genera Astragalus and Coronilla. Both species are widely distributed in Tunisia, mainly in arid and semiarid areas where they exhibit a large morphological variation and high seed yields (Zoghlami and Zouaghi, 2005, 2003). The ability of both species to survive over a range of climatic conditions suggests that hard seededness (seed coat impermeability) is an important germination regulating mechanism.
Temperature requirements for seed germination of some species explain why they are relatively easy or difficult to establish in field conditions. According to Towsend and McGinnies (1972) and Revell et al. (1998), species with very small seeds-A. hamosus in our case-germinate well at most temperatures. These species are often difficult to establish although this behaviour will greatly increase their chance of survival for the long term.
In the present study, temperature regime is a principal factor affecting the seed germination of A. hamosus. Alternating temperature treatment of 20-35°C was the most favourable temperature on total germination. This result agrees with that of Quinlivan (1961) who demonstrated that seeds of Astragalus sinicus became permeable under the influence of temperature fluctuation between 20 and 40°C. Germination of almost all populations at 15-25 and 20-35°C was completed by the 7th day (Fig. 1). Early high germination across a large range of temperatures may be beneficial for rapid establishment of the pasture crop in semiarid warm regions where soil moisture in the upper soil surface is available for only a short period (Brar et al., 1991). In our conditions, this temperature range can be encountered in early autumn (September-October) where sporadic and intense rain may occur frequently.
Populations of A. hamosus which germinated well at most temperatures will be easier to establish in the field than those which required specific temperatures. The difficulty in establishing this species could be related to its small seed size rather than to temperature requirements (Towsend and McGinnies, 1972).
For C. scorpioides, germination was very low and was not affected by temperature. Eighty percent of seeds were rotten and then died.
In conclusion, temperature had a strong effect on germination of A. hamosus populations and genetic variation is probably important. We think that higher temperature than 20-35°C could be tested for improving germination of this species.
REFERENCES
- Brar, G.S., J.F. Gomez, B.L. McMichael, A.G. Matches and H.M. Taylor, 1991. Germination of twenty forage legumes as influenced by temperature. Agron. J., 83: 173-175.
Direct Link - Patane, C. and F. Gresta, 2006. Germination of Astragalus hamosus and Medicago orbicularis as affected by seed-coat dormancy breaks techniques. J. Arid Environ., 67: 165-173.
Direct Link - Quinlivan, B.J., 1961. The effect of constant and fluctuating temperatures on the permeability Aust. J. Agric. Res., 12: 1009-1022.
Direct Link - Revell, C.K., G.B. Taylor and P.S. Cocks, 1998. Long-term softening of surface hard seeds of yellow serradella (Ornithopus compressus L.) in a range of environments. Aust. J. Agric. Res., 49: 673-686.
Direct Link - Zoghlami, A. and M. Zouaghi, 2003. Morphological variation in Astragalus hamosus L. and Coronilla scorpioides L. populations of Tunisia. Euphytica, 134: 137-147.
CrossRefDirect Link - Zoghlami, K.A. and M. Zouaghi, 2005. Repartition en Tunisie d'Astragalus hamosus L. et de Coronilla scorpioides L. Koch en fonction de quelques facteurs du milieu. Revue des. Regions Arides, 16: 51-67.
Direct Link