
Hisham Y. Al-Matubsi
Faculty of Pharmacy, Applied Science University-Amman, Jordan
Nagham A. Nasrat
Faculty of Pharmacy and Medical Sciences, Al-Ahliyya Amman University-Amman, Jordan
Ghaleb A. Oriquat
Faculty of Pharmacy and Medical Sciences, Al-Ahliyya Amman University-Amman, Jordan
Mahmoud Abu-Samak
Al-Ghad College for Health Sciences Abha., Abha, Saudi Arabia
Khtan A. Al-Mzain
Faculty of Pharmacy and Medical Sciences, Al-Ahliyya Amman University-Amman, Jordan
Maher Salim
Faculty of Pharmacy and Medical Sciences, Al-Ahliyya Amman University-Amman, Jordan
Pakistan Journal of Biological Sciences
Year: 2011 | Volume: 14 | Issue: 7 | Page No.: 425-432







ABSTRACT
The current study investigated the effect of natural diosgenin extracted from fenugreek seeds, in comparison with the pure standard diosgenin, plus chromium chloride (CrCl3) supplementation on high-cholesterol fed Japanese quails. Quails were randomly divided into four groups. Group one (n = 25) fed experimental diet only (control; contained basal diet supplemented with 1% cholesterol), group 2 (n = 20) fed experimental diet supplemented with CrCl3 only (400 μg kg-1 of body weight), groups 3 and 4 (n = 30 per group) were fed experimental diet supplemented with either 0.5% (w/w) of extracted diosgenin plus CrCl3 or pure standard diosgenin (0.5%) plus CrCl3 respectively, for 12 days. Blood samples were collected at days 0 and 12 for measuring levels of lipid profile. The work was carried out at Applied Science University and Amman University, Amman, Jordan during the period from October 2009 through October 2010. The mean levels of total cholesterol (TC) in control quails at d12 was significantly (p<0.01) increased compare to those at d0. Supplementation of diet with CrCl3 alone or CrCl3 with diosgenin either extracted or pure standard for 12 days showed a significant (p<0.01) decrease in TC and low density lipoprotein cholesterol (LDL-C) levels as compared to those in the control quails. While, high density lipoprotein cholesterol (HDL-C) increased significantly (p<0.01) in quails supplemented with diosgenin and CrCl3. At d12, the mean Superoxide Dismutase (SOD) activities in erythrocytes of quails in all supplemented groups was significantly (p<0.01) increased as compared to those in control group and was more pronounced in erythrocytes of quails supplemented with pure standard diosgenin plus CrCl3. These results indicated that the combined diosgenin and CrCl3 supplementation to high-cholesterol fed quails might induce a protective effect by both regulating lipid and antioxidative damage.
PDF Abstract XML References Citation
Received: April 18, 2011;
Accepted: June 08, 2011;
Published: July 13, 2011
How to cite this article
Hisham Y. Al-Matubsi, Nagham A. Nasrat, Ghaleb A. Oriquat, Mahmoud Abu-Samak, Khtan A. Al-Mzain and Maher Salim, 2011. The Hypocholesterolemic and Antioxidative Effect of Dietary Diosgenin and Chromium Chloride Supplementation on High-Cholesterol Fed Japanese Quails. Pakistan Journal of Biological Sciences, 14: 425-432.
DOI: 10.3923/pjbs.2011.425.432
URL: https://scialert.net/abstract/?doi=pjbs.2011.425.432
DOI: 10.3923/pjbs.2011.425.432
URL: https://scialert.net/abstract/?doi=pjbs.2011.425.432
INTRODUCTION
Mortality from cardiovascular disease is the second leading cause of death worldwide (Malach and Imperato, 2006). Hypercholesterolemia is generally accepted as an independent risk factor contributing to the development of coronary heart disease (Altman, 2003; Jalali-khanabadi et al., 2006). While controlled diet is the primary treatment of hyperlipidemia (Henley et al., 2002). It is more practicable to decrease the elevated lipid levels by treatment with anti-hyperlipidemic drugs. However, treatment by drugs is not always satisfactory due to side effects of chemically synthesized drugs (Yanovski and Yanovski, 2002; Garg and Simha, 2007). Phytopharmaceuticals are gaining importance in allopathic as well as traditional medicine owing to their non-addictive and non-toxic nature (Jenkins et al., 1983; Raskin et al., 2002).
Fenugreek (Trigonella foenum-graecum L.) is an herbaceous annual plant from the family of Leguminosae, cultivated in Mediterranean countries, North Africa and India. It seeds have long been known as a herbal remedy for various pathological conditions (Sharma, 1986; Subhashini et al., 2011). Fenugreek is commonly used against diabetes in Pakistan (Karim et at., 2011).
Steroidal Saponin obtained from fenugreek seeds are composed mainly of diosgenin. A number of pharmacological and clinical studies have shown glucose- and lipid-lowering properties of the seed itself, seed extracts or purified components (Sauvaire et al., 2000; Yadav et al., 2008).
Diosgenin has been shown to have favorable effects on glucose lowering (McAnuff et al., 2005), antioxidant activity (Son et al., 2007), lipid metabolism and myocardial infarction (Chiang et al., 2007; Jayachandran et al., 2009). Chromium (Cr+3) is an essential element for carbohydrate and lipid metabolism in animals and humans (El-Hommosany, 2008). Chromium has been shown to potentiate the action of insulin and accordingly to modulate carbohydrate, protein, and lipid metabolism (Vincent, 2000).
Some researchers highlighted its role with regard to the metabolism of lipids and the correlations between chromium status and cardiovascular disease (Rajpathak et al., 2004) in which low chromium level leads to high cholesterol and brings the risk of developing cardiovascular disorders. Furthermore, chromium chloride administration causes a substantial reduction of coronary lipid deposits, aortic lipid deposits and serum cholesterol concentration in rabbits (Bakhiet and Elbadwi, 2007; Price-Evans et al., 2009).
This study was carried out to investigate the combined effect of natural saponin (diosgenin) extracted from fenugreek seeds, in comparison with the pure standard diosgenin and chromium chloride (CrCl3) supplementation on high-cholesterol fed Japanese quails.
MATERIALS AND METHODS
Extraction of diosgenin from fenugreek seeds: Fenugreek seeds were purchased from the local market and their botanical identity was confirmed by a botanist from our University. Diosgenin was alcohol-extracted from fenugreek seeds according to the method of Prasanna (2000).
Chromatographic investigation: The bioactive extract of fenugreek seeds, diosgenin, and standard diosganin (≥99.0% purity, purchased from Fluka, BioChemika, West Germany) were subjected to chromatographic identification using thin layer chromatography (TLC) (National Research Council, 1994).
Animals and diets: Male Japanese quail (Coturnix japonica) 45 days old were purchased. They were housed in wire cages (5 birds/pen) at 22±2°C with a controlled 12-hr daylight cycle. The quails had ad libitum access to basal diet (Table 1) and water and kept under observation for 7 days before treatment was initiated. The basal diet was formulated using the National Research Council, (Yuan et al. 1998) guidelines and contained 20% crude protein and 12.9% MJ kg-1 Metabolizable Energy (ME). One hundred and five (105) of these quails were then fed an experimental diet (diet contained basal diet supplemented with 1% cholesterol) and continued throughout the experiment.
| Table 1: | Ingredients of basal diet |
 | |
| Vitamin premix provides the following per kilogram (kg) of diet: all-trans retinyl acetate: 1.8 mg; cholecalciferol: 0.025 mg; all-rac-α-tocopheryl acetate: 1.25 mg; menadione sodium bisulfite: 1.1 mg; thiamine-hydrochloride: 1.1 mg; riboflavin: 4.4 mg; Ca-pantotheenate: 10 mg; pyridoxine hydrochloride: 2.5 mg; vitamin B-6: 2.2 mg; vitamin B-12: 0.02 mg; folic acid: 0.55 mg; niacin: 35 mg; d-biotin: 0.1 mg; Mineral premix provides the following per kg:manganese: 40 mg; iron: 12.5 mg; zinc:25 mg; copper: 3.5 mg; iodine: 0.3 mg; selenium: 0.15 mg; choline chloride: 175 mg | |
| Table 2: | Effects of dietary diosgenin (either extracted or pure standard) and chromium chloride supplementation on performance of Japanese quails fed high-cholesterol for 12 days. |
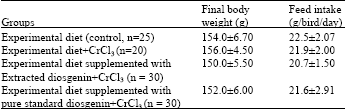 | |
| Values are expressed as Mean±SEM | |
Quails were randomly divided into four groups. Group one (n=25) fed experimental diet only (control), group 2 (n = 20) fed experimental diet supplemented with chromium chloride only (CrCl3, 400 μg kg-1 of body weight), groups 3 and 4 (n = 30 per group) were fed experimental diet supplemented with either 0.5% (w/w) of extracted diosgenin (Son et al., 2007) plus CrCl3, (400 μg kg-1 of body weight) or pure standard diosgenin (0.5%) plus CrCl3 (400 μg kg-1 of body weight) respectively for 12 days.
Diosgenin, either extracted or pure standard that fed to groups 3 and 4 was dissolved in chloroform, added to the basal diet on metal tray, air dried and mixed mechanically. The 1% cholesterol was added to the mixed diet. The feed intake was measured daily throughout the experiment (Table 2). Daily replacement of diets minimized exposure of birds to oxidized dietary lipids. Chromium chloride was dissolved in 1 ml distilled water and administered by oral gavage. The control group received water as placebo. The study was approved by the university committee for research.
Blood sampling: Fasting blood samples were collected, at day 0 and day 12 of experimental dietary treatments, from jugular vein of Japanese quail and placed in tubes containing either no anticoaggulant for measuring serum levels (mg dL-1) of Total Cholesterol (TC), Triglyceride (TG), high density lipoprotein cholesterol (HDL-C), low density lipoprotein cholesterol (LDL-C) and very LDL-C (VLDL-C), or EDTA for superoxide dismutase (SOD) enzyme activity in erythrocytes (measured at day 12 only).
Determination of serum lipid profile: The serum TC, TG and HDL-C, levels were measured using assay kits (Linear chemicals, Spain) based on enzymatic calorimetric methods. All other chemicals were of reagent grade. Serum LDL-C and VLDL-C levels were calculated according to Friedewald's formula (Guemouri et al., 1991) in which LDL-C = TC-HDL-C-TG/5 and VLDL-C = TG/5.
Erythrocytes superoxide dismutase extraction:: Blood samples (5 mL) were collected in EDTA tubes. The plasma and the majority of white blood cells were removed by centrifugation at 1000 rpm for 10 min. The red blood cells were washed twice with saline and hemolyzed by adding approximately 1.5 volume of distilled water. Five hundred micro-litters (500 μL) of hemolysate were then added to 4 mL of ice cold distilled water followed by 1 mL of ethanol and 0.6 mL of chloroform. The mixture was then shacked, centrifuged for 10 min at 3000 rpm and the clear top layer which contains the enzyme SOD was separated and stored at -20°C until assayed. The SOD activity in erythrocytes was assayed by the method described by Guemouri et al. (1991) using commercially available kits (Randox Lab. Ltd., Ireland) based on the photochemical Nitrobluetetrazolium (NBT) method.
Statistical analysis: Data are expressed as the Mean±SEM. The significance of differences between the groups was assessed by ANOVA using SPSS 11.5 statistical software. Value of (p<0.05) was considered significant.
RESULTS
Extraction of diosgenin from fenugreek seeds and TLC analysis: Thin layer chromatography (TLC) revealed that extracted diosgenin which appears as a dark pink spots after development with 50% H2SO4 has rate of flow (Rf) value equal 0.66 which is identical to the Rf value of the standard diosgenin as illustrated in the chromatogram (Fig. 1).
Body weights and feed intake: The effects of dietary diosgenin (either extracted or pure standard) and the chromium chloride supplementation on performance of Japanese quails fed high-cholesterol for 12 days are shown in Table 2. Final body weights and feed intake of birds were not significantly affected by dietary supplementation of either chromium chloride alone or with diosgenin among all the groups.
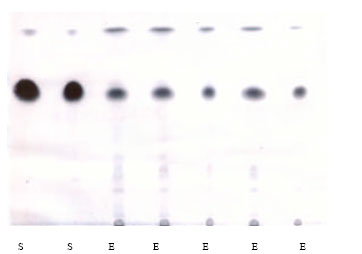 | |
| Fig. 1: | Chromatogram from thin layer chromatography (TLC) analysis using (Chloroform: Methanol) as mobile phase and developed by 50% H2SO4 S: Standard, E: Extract |
| Table 3: | Serum total cholesterol (TC) and triglyceride (TG) levels in Japanese quails fed different experimental diets for 12 days |
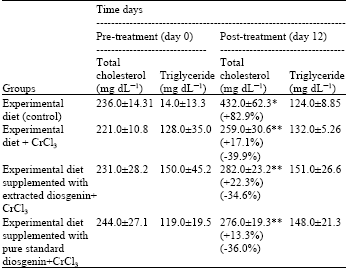 | |
| Values are expressed as Mean±SEM. *Significant difference (p<0.01) compared with control d0. **Significant difference (p<0.01) compared with control d12 | |
Serum total cholesterol (TC) and triglyceride (TG): The effect of supplemented CrCl3 alone or CrCl3 with diosgenin either extracted or pure standard in Japanese quails on serum TC and TG I is summarized in Table 3.
The mean levels of TC in quails fed with experimental diet only (control) for twelve days (induced hypercholesterolemia) was significantly (p<0.01) increased compare to those at pretreated (d0). The mean levels of TC in control quails at d12 (432.0±62.3 mg dL-1) was 82.9% more than that of d0 (236.0±14.3 mg dL-1).
At d12, the mean levels of TC in quails supplemented with CrCl3 alone or CrCl3 with diosgenin either extracted or pure standard was also increased by 17.1% (259.0± 30.6 mg dL-1), 22.3% (282.0±23.2 mg dL-1) and 13.3% (276.0±19.3 mg dL-1) above that determined at d0 (221.0±10.8 mg dL-1) (231.0±28.2 mg dL-1) and (244.0±27.1 mg dL-1) respectively, such increase did not carry statistical significance.
Supplementation of CrCl3 alone or CrCl3 with diosgenin either extracted or pure standard to the hypercholesterolemic quails produced a significant (p<0.01) drop in serum TC compared with the control group. Serum TC levels were significantly (p<0.01) decreased by 39.9% (259.0±30.6 mg dL-1) in quails supplemented with CrCl3 alone, 34.6% (282.0±23.2 mg dL-1) in quails supplemented with extracted diosgenin plus CrCl3 and by 36% (276.0±19.3 mg dL-1) in those supplemented with pure standard diosgenin plus CrCl3 as compared to control group (432.0±62.3 mg dL-1).
Although there is no significant difference in levels of TC between the three supplemented groups at d12, quails supplemented with CrCl3 alone shows the higher reduction in mean values of TC when compared with other groups supplemented with diosgenin (either pure standard or extracted) plus CrCl3. It should be mentioned that the supplementation of hypercholesterolemic quails with CrCl3 alone or CrCl3 with diosgenin either extracted or standard is effective in reducing TC values and succeeded to normalize them nearly to the base line.
As shown in Table 3, the mean serum levels of TG for quails supplemented with CrCl3 alone or CrCl3 with diosgenin either extracted or pure standard for 12 days (132.0±5.26, 151.0±26.6 and 148.0±21.3 mg dL-1; respectively) were not significantly (p>0.05) different from those in control quails (124.0±8.85 mg dL-1). Furthermore, there is no significant difference in the mean levels of TG at d12 when compared to those at d0 in all groups.
Serum lipid profile: The changes of lipid profile in the serum quails are presented in Table 4. At d0, the mean levels of HDL-C were not significantly different between control and supplemented groups.
The levels of HDL-C are significantly (p<0.01) increased at d12 in all groups when compared with those determined at d0. The level of HDL-C of the control quails fed only experimental diet for 12 days (90.9± 14.2 mg dL-1) was 86.6% higher than those in d0 (48.7±1.93 mg dL-1).
However, the serum levels of HDL-C of the quails supplemented with CrCl3 alone or CrCl3 with diosgenin either extracted or standard for 12 days (102.0±15.3, 135.0±20.8 and 126.0±15.7 mg dL¯1; respectively) were 129.0%, 199.0% and 156.0% higher than those determined at d0 (44.6±3.34, 45.2±3.86 and 49.1±6.18 mg dL-1; respectively).
Supplementation of the quails with diosgenin either extracted or standard plus CrCl3 for 12 days had a significant (p<0.01) effect on increasing HDL-C levels by 48.6 and 38.3%; respectively as compared to those in the control quails. However, supplementation of the quails with CrCl3 alone increase HDL-C levels by 12.4%, yet such increase did not carry statistical significance.
At d0, in a pattern similar to that of HDL-C, the serum levels of LDL-C were not significantly changed in all supplemented groups compared with those of controls.
In control quails, the mean levels of LDL-C at d12 was significantly (p<0.01) above (92.2%) the mean levels at d0 (316.0±42.8 vs 164.0±13.3 mg dL-1).
The serum levels of LDL-C in quails supplemented with CrCl3 alone or CrCl3 with diosgenin either extracted or pure standard for 12 days (131.0±18.4, 117.0±20.8 and 120.8±22.7 mg dL-1; respectively) were decreased by 13.4%, 24.8% and 29.2% below than those at d0 (151.0±18.1, 156.0±24.4 and 171.0±21.1 mg dL-1; respectively). Yet these differences were not statistically significant.
Supplementation of diet with CrCl3 alone or CrCl3 with diosgenin either extracted or pure standard to quails fed high cholesterol for 12 days showed a significant (p<0.01) decrease in LDL-C by 58.6, 62.9 and 61.7%, respectively, below the mean values for control quails.
| Table 4: | Serum HDL-C, LDL-C, VLDL-C levels in Japanese quails fed different experimental diets for 12 days |
 | |
| Values are expressed as Mean±SEM. *Significant difference (p<0.01) compared with d0 of the same group. **Significant difference (p<0.01) compared with control d12 | |
| Table 5: | Erythrocytes superoxide dismutase (SOD) activities in Japanese quails fed different experimental diets for 12 days |
 | |
| Values are expressed as Mean±SEM. *Significant difference (p<0.01) compared with control d12 | |
It should be note that there is no significant difference in serum LDL-C levels between all supplemented groups.
On day 12, the ratio of HDL:LDL was higher in the quails fed diet supplemented with CrCl3 alone (78.2 vs 29.5%) or CrCl3 with diosgenin either extracted (116.0 vs 29.1%) or pure standard (104.0 vs. 28.8%), when compared to day 0.
As shown in Table 4, serum levels of VLDL-C on d12 not differ from those in d0 in all groups. Furthermore, VLDL-C levels following 12 days supplementation with CrCl3 alone or CrCl3 with diosgenin either extracted or standard were not significantly differ compared to control quails (26.3±2.03, 30.1±7.08 and 29.6±4.48 vs 24.8±6.20 mg dL-1; respectively).
Erythrocytes SOD enzyme activity: The activities of SOD enzyme erythrocytes of the control and all supplemented quails are represented in Table 5. The antioxidative enzyme activity (SOD) was not only affected by CrCl3 supplementation but also by diosgenin plus CrCl3 supplementation. There was a significant (p<0.01) increase in the activities of SOD in all supplemented groups as compared to those in control group. The SOD activities in erythrocytes of quails supplemented with CrCl3 alone for 12 days was 4.88±0.76 U mg-1 Hb and showed a 52.5% above the mean values for control quails (3.20±0.71 U mg-1 Hb). However, this activity in erythrocytes of quails supplemented with extracted diosgenin plus CrCl3 was 5.34±0.73 U mg-1 Hb and showed a 66.9% above the mean values for control quails. The SOD activity in erythrocytes of quails supplemented with pure standard diosgenin plus CrCl3 was more pronounced and reach 72.8% (5.53±0.82 U mg-1 Hb) as compared to the mean value of controls. The mean SOD activity in erythrocytes of quails supplemented with CrCl3 alone did not show significant variation when compared p to those supplemented with diosgenin either extracted or pure standard lus CrCl3.
DISCUSSION
Using of quail model in this study has numerous advantages over others, including their small size, ease of management, their short life span, the rabidity of atherosclerosis plaque development. In addition, quails are naturally deficient in apolipoprotein E and exhibiting structural features (e.g., focal hemorrhage, calcification and fibrosis) that closely resemble those in the human disorders (Ojerio et al., 1972).
A recent study reports that Vytorin and Zetia, two major drugs in the treatment of high cholesterol, failed to decrease the incidence of heart disease (Mitka, 2008). Also, uses of anti-obesity drugs are severely restricted due to accompanying side effects (Wasan and Looije, 2005). Thus, there is still a need for efficient, safe and economic means to combat dyslipidemia and associated metabolic disorders.
One of naturally cholesterol absorption inhibitors is the saponins, the utility of the naturally occurring plant saponins as hypolipidemic agents is limited by difficulties in obtaining large quantities from natural sources. This is due in part to the presence of multiple saponins of differing structure and biological activity within a given species (Price et al., 1987). Thus, in the present study we used one fraction of saponin which is diosgenin in order to clear its individual effect and not to interfere with effects of other sapogenins. The present study demonstrated for the first time the hypocholesterolemic and antioxidative effect of natural saponin (diosgenin) extracted from fenugreek seeds, in comparison with the pure standard, plus CrCl3 supplementation on high-cholesterol fed Japanese quails.
The dose used in our pilot study (0.5%) was within (Rajpathak et al., 2004) or below than that used in previous studies. Previous investigators have been used diosgenin at dose of 80 mg kg-1 in rats (Jayachandran et al., 2009).
The Rf value for diosgenin recorded in this study was approximately similar to that reported by Li et al. (2002) which was 0.65. It is important to mention that the normal range for Rf value of diosgenin is between 0.20-0.70 (National Research Council, 1994). Although using of TLC, rather than high performance liquid chromatography, for the chromatographic determination of diosgenin in the extract is not the most adequate technique but it can be considered as an auxiliary technique in our case.
Trivalent chromium (Cr3+) is an essential trace element for animals and humans. It was suggested that deficiency of chromium could cause hypoglycemia and high level of cholesterol which are critical cardiovascular risk factors (Rajpathak et al., 2004). Chromium supplementation lowers blood levels of proinflammatory cytokines, oxidative stress, and lipids levels in diabetic rats (Jain et al., 2007).
The changes of total cholesterol and triglycerides levels during the experimental period are presented in Table 3. Feeding quails with the high-cholesterol diet for 12 days resulted in hypercholesterolemia, as was evident from the significant increase in serum total cholesterol level of control group at d12 compared to d0 and does not induce hypertriglyceridemia. Supplementation of the high-cholesterol diet with diosgenin either extracted or pure standard plus CrCl3 reduced serum TC by about 34.6% and 36.0% respectively compared to the quails fed on the high-cholesterol diet alone. These results are consistent with recent studies revealed that dietary diosgenin (Temel et al., 2009) has a marked hypocholesterolemic effect.
It is noteworthy to mention that the potent effect of reducing elevated TC and LDL-C levels were similar in both diosgenin supplemented groups (extracted and pure standard) which conforms diosgenin purity and successful extraction steps.
In the present study, reduction of TC levels was not statistically different between extracted and standard diosgenin (plus CrCl3), though as can be seen in Fig. 1 (TLC chromatogram), the extract has a much lower concentration of diosgenin than the standard. The possible explanation is that the level of supplementation of both extract and standard were well above the minimum, thus both supplements were capable of returning TC values nearly to base line. Moreover, it is possible that the lack of difference is in part due to the high dosage of CrCl3.
The reduction in serum TC reported in the present study was reflected by significant reduction in the serum levels of LDL-C. These results are in agreement with other studies observed that the reduction in serum cholesterol was in LDL (Sowmya and Rajyalakshmi, 1999). Furthermore, the significant increase in the serum level of HDL-C associated with the reduction in serum TC of quails fed diet supplemented with CrCl3 alone or CrCl3 with diosgenin either extracted or standard for 12 days contradict the recent study carried by Niu et al. (2009) finding that the serum level of HDL was decreased in hyperlipidemic rats after administration niacin and chromium for 12 weeks comparing with high-fat group.
Yet in the present study, the results indicated that the supplemented diets, CrCl3 alone or CrCl3 with diosgenin either extracted or pure standard, offered to quails fed high cholesterol for 12 days, provided a better serum lipid profiles when compared to the control diet, as was evident from HDL:LDL ratio.
It is well known that diosgenin lowers plasma cholesterol by increasing faecal cholesterol excretion. Until recently it was believed that this effect was due both to an inhibition of intestinal absorption of cholesterol as a stimulation of its biliary secretion (by increasing LDL and HDL hepatic clearance). However, Temel et al. (2009) demonstrated that stimulation of faecal cholesterol excretion by diosgenin is independent of Niemann-Pick C1-Like 1 (NPC1L1)-mediated cholesterol absorption (NPC1L1 is the protein with major role in intestinal cholesterol absorption). Therefore, the hypocholesterolemic effect of dietary diosgenin by increasing of faecal cholesterol excretion is primarily attributable to its impact on hepatic cholesterol metabolism rather than intestinal cholesterol absorption.
Furthermore, we found that the diosgenin (0.5%) and CrCl3 (400 μg kg-1 of body weight) supplementation had no effect on the serum triglyceride level. This result may be explained by the short duration of the supplementation.
In this study, supplementation of the high-cholesterol diet with CrCl3 alone reduced serum TC by about 39.9% which is in agreement with previous reports showed that dietary Cr3+ lowers circulating cholesterol concentrations in rats (Vincent, 2000; Jain et al., 2007) and birds (Kim et al., 1996). However, others failed to find the effects of Cr3+ on blood cholesterol concentrations (Kim et al., 2010).
The dose of CrCl3 used in this study (400 μg CrCl3 kg-1 of body weight) is similar to the previous studies using quail (Sahin et al., 2005). Thus, CrCl3 supplementation dose used per body weight in the present study is much higher than that used in human clinical trials. Whether or not there are any differences in the absorption of Cr3+ between humans and quails is not known.
The biological effects of free radicals are controlled in vivo by a wide range of antioxidants such as vitamins E and C, carotenoids, glutathione and anti-oxidative enzymes. Among these enzymes, SOD catalyzes dismutation of the superoxide anion into hydrogen peroxide (Ojerio et al., 1972). A reduced antioxidative defense status in the plasma and erythrocytes might result in increased peroxidation of cell membrane lipids and hence an increased plasma concentration of lipid peroxides (Buczynski et al., 1993), thus, it might play an important role in atherogenesis.
Under hypercholesterolemic conditions, the red blood cells are known to accumulate cholesterol from LDL by a nonreceptor-mediated process which is enhanced by oxidation of LDL (Panasenko et al., 1991). Red blood cells may therefore provide a useful experimental model system to investigate influences of hyperlipidemia on tissue antioxidant status.
In this study, we examined the possible preventive role of combined effect of two hypocholesterolemic and antioxidant agents, diosgenin (either extracted or pure standard) plus CrCl3 or CrCl3 alone on high-cholesterol fed Japanese quails. Present results revealed higher erythrocyte SOD activities in all supplemented groups compared to those in the control group and was more pronounced in erythrocytes of quails supplemented with pure standard diosgenin plus CrCl3. Therefore, diosgenin may play a role like that of a lipid peroxidation chain-breaking antioxidant in cell membrane and might provide protection against the oxidative damaging effects of polyunsaturated fatty acid.
On the other hand, it might be assumed that the increased activity of antioxidative enzymes in all supplemented groups played an important role in restricting the production of the active oxygen species induced by hypercholesterolemia. This dogma further supported by previous study showed that an excess of trivalent chromium can act as a prooxidant (Terpilowska and Zaporowska, 2004).
In conclusion, the present study revealed that the combined diosgenin and chromium chloride supplementation to high-cholesterol fed quails produced greater decrease in cholesterol and LDL-C and greater increase in HDL-C than did chromium chloride alone by both improving the lipid profile and modulating oxidative stress. Thus, suggests its potential usefulness as an agent for treating human hypercholesterolemia.
REFERENCES
- Altman, R., 2003. Acute coronary disease Athero-Inflammation: Therapeutic approach. Thromb. J., 1: 2-7.
PubMed - Jalali-Khanabadi, B.A., H. Mozaffari-Khosravi, M. Rafiei, S.M. Ghoreishian and F. Darabi, 2006. Association of cholesterol-rich lipoproteins with coronary artery disease in subjects who referred to yazd cardiovascular research center for coronary angiography. Pak. J. Biol. Sci., 9: 2777-2780.
CrossRefDirect Link - Henley, E., L. Chang and S. Hollander, 2002. Treatment of hyperlipidemia. J. Fam. Pract., 51: 370-376.
PubMed - Garg, A. and V. Simha, 2007. Update on dyslipidemia. J. Clin. Endocrinol. Metab., 92: 1581-1589.
PubMed - Raskin, I., D.M. Ribnicky, S. Komarnytsky, N. Ilic and A. Poulev et al., 2002. Plants and human health in the twenty-first century. Trends Biotechnol., 20: 522-531.
CrossRefPubMedDirect Link - Jenkins, D.J., G.S. Wong, R. Patten, J. Bird and M. Hall et al., 1983. Leguminous seeds in the dietary management of hyperlipidemia. Am. J. Clin. Nutr., 38: 567-573.
PubMed - Sharma, R.D., 1986. Effect of fenugreek seeds and leaves on blood glucose and serum insulin responses in human subjects. Nutr. Res., 6: 1353-1364.
Direct Link - Subhashini, N., G. Nagarajan and S. Kavimani, 2011. Anti-inflammatory and in vitro antioxidant property of Trigonella foenum graecum seeds. J. Pharmacol. Toxicol., 6: 371-380.
CrossRefDirect Link - Yadav, M., R. Tomar, GBKS. Prasad, S. Jain and H. Yadav, 2008. Complementary hypoglycemic and anti-hyperglycemic activity of various extracts of fenugreek seeds in rats. Asian J. Biochem., 3: 182-187.
CrossRefDirect Link - Chiang, C.T., T.D. Way, S.J. Tsai and J.K. Lin, 2007. Diosgenin, a naturally occurring steroid, suppresses fatty acid synthase expression in HER2-overexpressing breast cancer cells through modulating Akt, mTOR and JNK phosphorylation. FEBS Lett., 581: 5735-5742.
CrossRefPubMedDirect Link - McAnuff, M.A., W.H. Wayne, F.O. Omoruyi, H. Jacobs, E.Y. Morrison and H.N. Asemota, 2005. Hypoglycemic effect of steroidal sapogenins isolated from Jamaican bitter yam, Dioscorea polygonoides. Food Chem. Toxicol., 43: 1667-1672.
CrossRef - Jayachandran, K.S., H.R. Vasanthi and G.V. Rajamanickam, 2009. Antilipoperoxidative and membrane stabilizing effect of diosgenin, in experimentally induced myocardial infarction. Mol. Cell. Biochem., 327: 203-210.
CrossRefPubMedDirect Link - Son, I.S., J.H. Kim, H.Y. Sohn, K.H. Son, J.S. Kim and C.S. Kwon, 2007. Antioxidative and hypolipidemic effects of diosgenin, a steroidal saponin of yam (Dioscorea spp.), on high-cholesterol fed rats. Biosci. Biotechnol. Biochem., 71: 3063-3071.
CrossRefDirect Link - El-Hommosany, Y.M., 2008. Study of the physiological changes in blood chemistry, humoral immune response and performance of quail chicks fed supplemental chromium. Int. J. Poult. Sci., 7: 40-44.
CrossRefDirect Link - Vincent, J.B., 2000. Elucidating a biological role for chromium at a molecular level. Acc. Chem. Res., 33: 503-510.
PubMed - Rajpathak, S., E.B. Rimm, T. Li, J.S. Morris, M.J. Stampfer, W.C. Willett and F.B. Hu, 2004. Lower toenail chromium in men with diabetes and cardiovascular disease compared with healthy men. Diabetes Care, 27: 2211-2216.
PubMed - Bakhiet, A.O. and S.M.A. Elbadwi, 2007. Effects of dietary chromium supplementation on the performance and some serum parameters in bovans-type chicks. J. Pharmacol. Toxicol., 2: 402-406.
CrossRefDirect Link - Prasanna, M., 2000. Hypolipidemic effect of fenugreek: A clinical study. Indian J. Pharmacol., 32: 34-36.
Direct Link - NRC., 1994. Nutrient Requirements of Poultry. 9th Rev. Edn., National Academy Press, Washington, DC., USA., ISBN-13: 978-0309048927, Pages: 176.
Direct Link - Yuan, Y.V., D.D. Kitts and D.V. Godin, 1998. Interactive effects of increased intake of saturated fat and cholesterol on atherosclerosis in the Japanese quail (Coturnix japonica). Br. J. Nutr., 80: 89-100.
PubMed - Guemouri, L., Y. Artur, B. Herbeth, C. Jeandel, G. Cuny and G. Siest, 1991. Biological variability of superoxide dismutase, glutathione peroxidase and catalase in blood. Clin. Chem., 37: 1932-1937.
Direct Link - Ojerio, A.D., G.J. Pucak, T.B. Clarkson and B.C. Bullock, 1972. Diet-induced atherosclerosis and myocardial infarction in Japanese quail. Lab. Anim. Sci., 22: 33-39.
PubMed - Wasan, K.M. and N.A. Looije, 2005. Emerging pharmacological approaches to the treatment of obesity. J. Pharm. Pharm. Sci., 8: 259-271.
PubMed - Price, K.R., I.T. Johnson, G.R. Fenwick and M.R. Malinow, 1987. The chemistry and biological significance of saponins in foods and feedingstuffs. Crit. Rev. Food Sci. Nutr., 26: 27-135.
CrossRefPubMedDirect Link - Li, H., A. Radunz, P. He and G.H. Schmid, 2002. Influence of different light intensities on the content of diosgenin, lipids, carotenoids and fatty acids in leaves of dioscorea zingiberensis. Z. Naturforsch. C., 57: 135-143.
PubMed - Jain, S.K., J.L. Rains and J.L. Croad, 2007. Effect of chromium niacinate and chromium picolinate supplementation on lipid peroxidation, TNF-α, IL-6, CRP, glycated hemoglobin, triglycerides and cholesterol levels in blood of streptozotocin-treated diabetic rats. Free Radical Biol. Med., 43: 1124-1131.
CrossRef - Temel, R.E., J.M. Brown, Y. Ma, W. Tang and L.L. Rudel et al., 2009. Diosgenin stimulation of fecal cholesterol excretion in mice is not NPC1L1 dependent. J. Lipid. Res., 50: 915-923.
PubMed - Sowmya, P. and P. Rajyalakshmi, 1999. Hypocholesterolemic effect of germinated fenugreek seeds in human subjects. Plant Food Hum. Nutr., 53: 359-365.
CrossRefPubMedDirect Link - Sahin, N., K. Sahin, M. Onderci, M.F. Gursu, G. Cikim, J. Vijaya and O. Kucuk, 2005. Chromium picolinate, rather than biotin, alleviates performance and metabolic parameters in heat-stressed quail. Br. Poult. Sci., 46: 457-463.
CrossRefDirect Link - Buczyński, A., B. Wachowicz, K. Kedziora-Kornatowska, W. Tkaczewski and J. Kedziora, 1993. Changes in antioxidant enzymes activities, aggregability and malonyldialdehyde concentration in blood platelets from patients with coronary heart disease. Atheroscler, 100: 223-228.
PubMed - Panasenko, O.M., T.V. Volnova, O.A. Azizova and Y.A. Vladimirov, 1991. Free radical modification of lipoprteins and cholesterol accumulation in cells upon atherosclerosis. Free Radic. Biol. Med., 10: 138-148.
CrossRef - Terpiłowska, S. and H. Zaporowska 2004. The role of chromium in cell biology and medicine. Przegl. Lek., 61: 51-54.
PubMed - Karim, A., M.N. Sohail, S. Munir and S. Sattar, 2011. Pharmacology and phytochemistry of Pakistani herbs and herbal drugs used for treatment of diabetes. Int. J. Pharmacol., 7: 419-439.
CrossRef