
F. A. Bhuiyan
Department of Applied Bioresource Science, The United Graduate School of Agricultural Sciences, Ehime University, 3-5-7 Tarumi, Matsuyama, Ehime, 790-8566, Japan
S. Nagata
Department of Bioresources Science, Faculty of Agriculture, Kochi University, 200 Monobe, Nankoku, Kochi 783-8502, Japan
K. Ohnishi
Research Institute of Molecular Genetics, Kochi University, 200 Monobe, Nankoku, Kochi 783-8502, Japan
Pakistan Journal of Biological Sciences
Year: 2011 | Volume: 14 | Issue: 3 | Page No.: 204-211







ABSTRACT
Chitinase degrades chitin which comprises an important source of carbon and nitrogen in the marine environment. The aim of this study was to evaluate the population of chitinases in the marine sediments in southwest Japan. We collected marine sediments from eutrophic inner bay and offshore. Chitin-degrading bacteria were enriched from both sediments. Metagenomic DNA was isolated from the enriched chitin-degrading bacterial cell culture. At the same time, 25 chitin-degrading bacteria were isolated from the enriched culture. Partial fragments of chitinase genes were successfully amplified with degenerate primers designed for the glycoside hydrolase 18 family. We analyzed chitinase gene sequences of about 500 clones from metagenomic DNA prepared from chitin-degrading bacteria. Based on translated amino acid sequences, chitinases were grouped into five groups. Chitinases in groups II and III was most abundant and close to chitinase genes of several species of proteobacteria. On the other hand, chitinases in groups I, IV and V were unique and distinct from the known chitinases. These results indicate that the marine sediments used in this study contain diversity of chitinase genes.
PDF Abstract XML References Citation
Received: December 15, 2010;
Accepted: February 18, 2011;
Published: March 11, 2011
How to cite this article
F. A. Bhuiyan, S. Nagata and K. Ohnishi, 2011. Novel Chitinase Genes from Metagenomic DNA Prepared from Marine Sediments in Southwest Japan. Pakistan Journal of Biological Sciences, 14: 204-211.
DOI: 10.3923/pjbs.2011.204.211
URL: https://scialert.net/abstract/?doi=pjbs.2011.204.211
DOI: 10.3923/pjbs.2011.204.211
URL: https://scialert.net/abstract/?doi=pjbs.2011.204.211
INTRODUCTION
Chitin is an insoluble linear β-1, 4-linked polymer of N-acetylglucosamine (GlcNAc) (Fig. 1). It is found in the cell walls of fungi and certain algae, in the cuticles of arthropods, in the peritrophic membranes of annelids and in the shells of molluscs (Cauchie, 2002). Chitinases (EC 3.2.2.14) hydrolyze bonds between GlcNAc residues releasing oligomeric, dimeric (chitobiose) or monomeric products (Karlsson and Stenlid, 2009). Fungi and bacteria are thought to be important degraders of chitin in soil and thereby contribute to the recycling of carbon and nitrogen resources in soil ecosystems (Ikeda et al., 2007). In bacteria, the primary role of the chitinases is thought to digest and utilize chitin as a carbon and energy source (Cohen-Kupeic and Chet, 1998). Bacterial chitinases are often associated with the outer membrane or are secreted as extracellular enzymes which suggest that they must be adapted to the function under the physicochemical conditions of the environment (LeCleir et al., 2004).
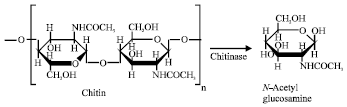 | |
| Fig. 1: | Chitin degradation by chitinase |
Chitinases are divided into two different glycoside hydrolase families (18 and 19) based on amino acid sequence similarity (Fukamizo, 2000). The vast majority of bacterial chitinases fall within family 18. These two families share limited similarity at the amino acid level and have different three-dimensional structures and moods of action (Li and Greene, 2010). Chitinase can display either exo or endo activity, depending on the structure of the catalytic site (Van Alten et al., 2000).
Glycoside hydrolase family 18 is subdivided into three groups, ChiA, ChiB and ChiC, based on the differences in the amino acid sequences of their catalytic domains (Suzuki et al., 1999). ChiA and ChiB are processive chitinases that degrade chitin chains in opposite directions, while ChiC is a nonprocessive endochitinase (Horn et al., 2006). Due to their prevalence in nature, ChiA have been used for studying the diversity and distribution of chitinolytic bacteria in both terrestrial systems and aquatic environment including the marine environment (Cottrell et al., 1999; Metcalfe et al., 2002; Ramaiah et al., 2000; Tsujibo et al., 2003).
Information on microbial chitinases has been prominently restricted to a few culturable microorganisms belonging to β and γ proteobacteria, gram-positive bacteria and the domain Archaea (Cottrell et al., 2000; Gao et al., 2003; Tanaka et al., 1999). The use of chitinase genes as molecular markers has demonstrated the presence of a large pool of uncultured chitinolytic microorganisms in the marine and soil environments (Cottrell et al., 2000; Metcalfe et al., 2002; Ramaiah et al., 2000). In this study, we have exploited the metagenome which was extracted from marine sediment and we have investigated the diversity of chitinase genes using the molecular method. The 16S rDNA and chitinase gene were analyzed with the enriched cultures from different marine sediments with colloidal chitin.
MATERIALS AND METHODS
Sampling sites and sediment sample collection: Sediment samples were collected from two different locations at Kochi prefecture in Japan on November 20, 2006, February 27, 2007 and June 28, 2007. One was coastal site of Uranouchi Bay, where maximum depth is 18m and another was about 20 km distance from the coast, offshore, where maximum depth is 200 m. Marine sediment was collected with a core sampler with 30 cm in length and sliced into 5 cm slices. The sediment samples were cooled immediately on ice and stored at -80°C for molecular analysis.
Extraction of metagenomic DNA: Sediment was suspended in 100 mL of artificial seawater (TetraMarin, Tetra, Germany) with stirring for 30 min at 4°C at moderate speed. After removal of soil, microorganisms were trapped on a 0.3 μm membrane filter and then dispersed in 5 mL of artificial seawater. Cells were collected with ultra-centrifugation at 35,000 rpm for 30 min at 4°C and stored at -80°C until use. Microbial metagenomic DNA was extracted using SoilMaster DNA extraction kit (Epicentre, Madison, WI, USA) according to the manufacturer’s protocol with slight modification. Cells were suspended in the buffer and bead-beated with 1 mm beads for 30 sec at 4,500 rpm using MS-100 Micro Smash (Tomy, Tokyo, Japan).
Enrichment of chitin-degraders: Microorganisms trapped on the membrane filter were suspended in 100 mL of 4% colloidal alpha chitin (from crab shell, Sigma-Aldrich, St. Louis, MO, USA) and incubated at 25°C for 7-28 days, until chitin solution became clear. Microorganisms were spread on agar plates containing colloidal alpha chitin and incubated at 25°C for 7 days, until halos were formed around the colonies. Colonies were suspended in LB media and incubated overnight at 25°C. Chromosomal DNA was extracted using AquaPure Genomic DNA Kits (Bio-Rad Lab. Hercules, CA, USA).
Multiple displacement amplification: Metagenomic DNA was amplified by Multiple Displacement Amplification (MDA) using GenomiPhi DNA amplification kit (Amersham Bioscience, Piscataway, NJ) following the manufacturer’s recommendations.
PCR amplification of chitinase gene fragment: Degenerate PCR primers (chiAF2U: 5'-GGAGACAUCGTGGACATGGGGARTWYCC-3' and chiAR2U: 5'-GGGAAAGUCCCAGGCGCCGTAGAR RTCRTARSWCA-3') designed according to the conserved regions in the catalytic domain of glycoside hydrolase family 18 were used for amplification of chitinase gene fragments. PCR primers (chiAF2: 5'-CGTGGACATGGGGARTWYCC-3' and chiAR2: 5'-CCCAGGCGCCGTAGARRTCRTARSWCA-3') were used for chitinase gene detection. Average sizes of the amplified fragments were 200-300 bp. PCR condition was an initial denaturation for 5 min at 85°C, followed by 30 cycles of 95°C for 45 sec, 58°C for 1 min and 72°C for 1 min. PCR products of chitinase gene fragments were cloned using USER Friendly System cloning kit (New England Biolabs, Beverly, MA, USA) according to the manufacturer’s protocol.
Phylogenetic analysis: The 16S rDNA was amplified by PCR with primers (Pr0RU 5'-GGAGACAUAGAGTTTGATCMTGGCTCAG-3' and 907RU: 5'-GGGAAAGUCCGTCAATTCCTTTRAGTTT-3'). Sequence analysis was conducted with a big dye Terminator ver. 3.1 cycle sequencing kit (Applied Biosystems, Foster City, CA, USA) using primer, RV-M (5'-GAGCGGATAACAAT TTCACACAGG-3') or M13-47 (5'-CGCCAGGGTTTTCCCAGTCACGAC-3'). Nucleotide sequences were determined by an ABI PRISM 3100-Avant Genetic Analyzer (Applied Biosystems). The 16S rDNA was classified using Ribosomal Database Project (Wang et al., 2007). The phylogenetic tree was constructed using the V1-V5 regions of the amplified 16S rDNA sequences with the Weighbor Joining method (The Ribosomal Database Project, http://rdp.cme. msu.edu/index.jsp). The known 16S rDNA sequences are retrieved from GenBank / EMBL / DDBJ databases. Bootstrap values with 100 replicates are shown for branches.
The translated amino acid sequences of glycoside hydrolase 18 family of chitinase genes, encoding catalytic domain, were aligned with CLUSTAL W (Thompson et al., 1994). The phylogenetic trees were constructed by using neighbor-joining method and visualized with tree view software (Page, 1996). Chitinase of Bacillus circulans No.4.1 (AAF23368) was used as an out-group. The known chitinase sequences of Vibrio parahaemolyticus RIMD2210633 (NP-800687), an uncultured organism ChiCSR14 (AAU12436), Vibrio sp. CJ11027 (AAV38107), Vibrio proteolyticus (BAF76068), Vibrio cholerae V51 (EAZ50998), Moritella sp. PE36 (ZP-01898217), Pseudoalteromonas sp. S9 (AAC79667), a marine bacterium EE36 (AAF21119), Aeromonas hydrophila JP101 (AAG09437), Shewanella denitrificans OS217 (ABE55418), Shewanella woodyi ATCC 51908 (ZP-01540956), Burkholderia cepacia AMMD (YP-773320), Epiphyas postvittana NPV (NP-203279), Reinekea sp. MED297 (ZP-01113517), an uncultured bacterium GG1-1 (BAE94857) and Cellulomonas uda ATCC21399 (AAG27061) are retrieved from GenBank/EMBL / DDBJ databases. Bootstrap analysis for 100 replicates was performed to estimate the confidence of phylogenetic tree topologies.
RESULTS
Diversity of bacterial population in marine sediments of two different locations was analyzed by using 16S rDNA sequences. In southwest Japan, Uranouchi Bay is a eutrophic inner bay and the offshore is 20 km far away from coast which is poor in nutrient. Metagenomic DNA isolated from marine sediments was treated with MDA and partial fragments of 16S rDNA were amplified and cloned. We randomly analyzed 16S rDNA sequences of 186 clones from Uranouchi Bay sediment and 48 clones from the offshore sediment. The 16S rDNA sequences were assigned to the bacterial taxonomy (Table 1) using naïve Bayesian classifier (Wang et al., 2007). The most abundant phylum was unclassified bacteria for Uranouchi Bay and proteobacteria for the offshore. These results indicated that bacterial population in the marine sediments of the bay and offshore were quite different. This might reflect that Uranouchi Bay is nutrient rich and the offshore is nutrient poor.
| Table 1: | Bacterial population in the sediment |
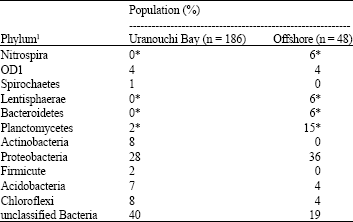 | |
| 1Based on naïve Bayesian rRNA classifier of the Ribosomal Database Project (Wang et al., 2007). *p-valuefor the Student's t-test was less than 0.01 which indicates that two values are significantly different | |
Sediment samples were enriched with 4% colloidal alpha chitin and after several passages, metagenomic DNA was extracted. Since enough numbers of bacterial cells were obtained, we did not apply MDA for these metagenomic DNAs. Partial fragments of 16S rDNA were amplified and cloned. We randomly analyzed 16S rDNA sequences of 113 clones from Uranouchi Bay sediment and 45 clones from the offshore sediment and assigned 16S rDNA sequences to the bacterial taxonomy (Table 2). Unlike the results with non-enriched sediments, all the clones were assigned to the known phyla. More than 60% of the bacterial clones enriched from Uranouchi Bay were identified as Rhodobactereaceae which was not detected from the offshore sediments (Fig. 2). On the other hand, more than 70% clones isolated from the offshore belong to Vibrionaceae which is abundant in the ocean environment (Table 2). These results indicated that different populations of chitin-degrading bacteria were enriched from the different marine sediments.
Bacterial cells grown in the chitin-enriched culture were spread on agar media containing colloidal alpha chitin. We isolated more than 40 strains as chitin degraders which formed halos around colonies. Isolated bacteria were classified based on 16S rDNA sequences and there was no significant difference between species isolated from Uranouchi Bay and the offshore (Table 3). Almost all strains belong to the familyVibrionaceae.
We used degenerate PCR primers, chiAF2 and chiAR2 which were designed by Hobel et al. (2005) to amplify the central catalytic domain of chitinase genes belonging to group A of glycoside hydrolase family 18 (GH18) (Coutinho and Henrtissat, 1999). We could amplify chitinase gene fragments from all the culturable chitin-degrading bacteria and the size of all fragments was 215 bp. 11 and 2 different deduced amino acid sequences were obtained from Uranouchi Bay and the offshore, respectively.
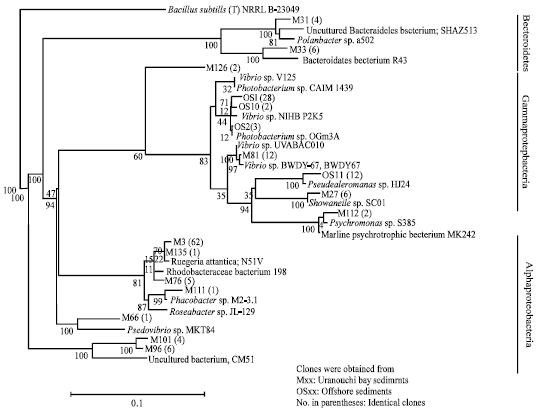 | |
| Fig. 2: | Phylogenetic tree based on 16S rDNA prepared from sediments enriched with chitin. A scale bar displays 0.1 nucleotide substitutions per site |
| Table 2: | Bacterial population after enrichment with chitin |
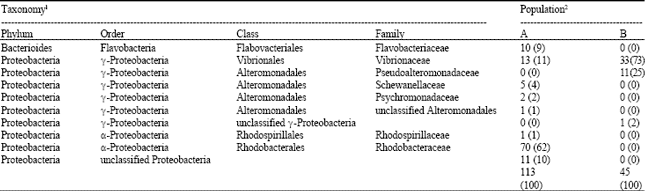 | |
| 1Based on naïve Bayesian rRNA classifier of the Ribosomal Database Project (Wang et al., 2007). 2Numbers of 16S rDNA clones isolated from A) Uranouchi Bay and B) the offshore were indicated. Percentage of clones belonging to each family was indicated in parentheses | |
| Table 3: | Chitin-degrading bacteria isolated from marine sediments |
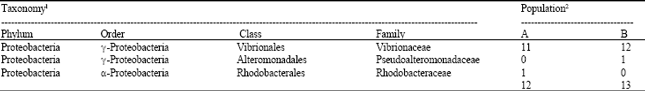 | |
| 1Based on naïve Bayesian rRNA classifier of the Ribosomal Database Project (Wang et al., 2007). 2Numbers of bacteria isolated from A) Uranouchi Bay and B) the offshore were indicated | |
From metagenomic DNA, several different sizes, 182, 215 and 239 bp, of fragments were amplified. After cloning, we randomly analyzed approximately 500 clones, from both Uranouchi Bay and the offshore. Nucleotide sequences of 182 and 239-bp fragments were quite different from those of 215-bp fragments.
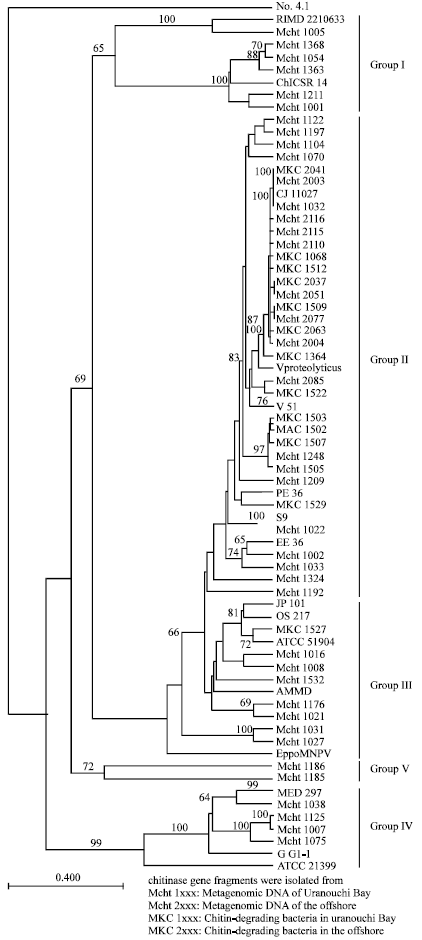 | |
| Fig. 3: | Phylogenetic tree of chitinases. Bootstrap values with 100 replicates are shown for branches with more than 50% bootstrap support |
The phylogenetic tree was constructed with deduced amino acid sequences which were grouped into five groups (Fig. 3). As expected, chitinase genes of the shorter fragments and longer fragments formed distinct clades, group IV, group I and group V, respectively. All these novel GH18 family of chitinase genes were isolated from Uranouchi Bay sediment (Fig. 3). Chitinase genes belonging to group II and group III were isolated from Uranouchi Bay and the offshore in a culture-dependent way and culture-independent metagenome.
DISCUSSION
Marine sediments support high biodiversity and vast ecosystem services and microbial populations widespread more in marine environment than terrestrial earth (Snelgrove, 1999). From the ecological viewpoints of the metagenome-based study, there were great interests that varieties of bacterial population existed in both marine sediments (Table 1, Fig. 2). We observed bacterial population of 12 phyla, Nitrospira, Spirochaetes, Lentisphaerae, Bacteroidetes, Planctomycetes, Actinobacteria, Proteobacteria (α, β, γ and δ subdivisions), Frimicute, Acidobacteria, Chloroflexi, unclassified bacteria and OD1 (Table 1). The presence of unclassified bacteria in the Uranouchi Bay sediment was numerically higher than in the offshore sediment. In the assessment, bacterial populations in the Uranouchi Bay sediment and in the offshore sediment were quite different and relatively more varieties of bacterial populations existed in the Uranouchi Bay sediment, because Uranouchi Bay is eutrophic. Bacterial population has changed in response to the changing environment in a coastal system, such as dissolved organic carbon (Takenaka et al., 2007). Taken together, it was demonstrated that marine sediments are good reservoirs of a variety of bacterial populations.
Although several novel enzymes for biocatalysts have been screened from metagenomic libraries, there are very few metagenomic studies targeting chitinases from a marine environment (Steele et al., 2009). In this study, nucleotide sequences of approximately 500 clones containing gene fragments amplified with degenerate primers for ChiA (Hobel et al., 2005) were determined and all of them were the GH18 family of chitinase genes. The phylogenetic tree was constructed based on deduced amino acid sequences of chitinase genes and chitinase genes detected in this study were grouped into five groups (Fig. 3). Chitinase genes isolated from metagenomic DNA of Uranouchi Bay sediment were affiliated with all five groups. Chitinase genes in group I form a clade with a clone ChiCSR14 which is isolated from an intertidal hot springs on the seashoreof northwest Iceland (Hobel et al., 2005). The clone Mcht1005 belonging to group I is closest to the chitinase gene of a clinical isolate of Vibrio parahaemolyticus (Makino et al., 2003). The clone Mcht1038 belonging to group IV is closest to the chitinase gene of Reinekea blandensis MED297 which is collected in the NW Mediterranean Sea.
Chitinase genes obtained from the culturable chitin-degrading bacteria and isolated from metagenomic DNA of the offshore sediment belong to either group II or III. Group II contains the most abundant chitinase genes in this study which exist in Vibrio sp. Group III contains chitinase genes of several species of proteobacteria. Chitin-degrading bacteria are isolated from sediment from the Gulf of Eilat in Israel (Hoster et al., 2005). Most of bacteria belong to Streptomyces and Bacillus while all the bacteria isolated in this study were classified as proteobacteria. This could support the idea that chitinase genes are laterally transferred between groups of bacteria (Cottrell et al., 2000). Group V consists of only two clones, Mct1185 and Mct1186 which have no close relatives and form a new phylotype.
As we did in this study, Hjort et al. (2010) screen for chitinase genes in a phytopathogen-suppressive soil in different ways, from a microbial metagenomic library and from bacterial isolates with chitinase activities. Chitinase gene groups are specifically detected in different sources of DNA. Some clusters contain chitinase sequences from the metagenomic DNA library and sequences from isolates. Other cluster contains contained chitinase sequences from the metagenomic library. This data is consistent with present results in this study.
A total of 18 chitinase genes are detected using the chiA specific primer (Hobel et al., 2005) from deep-sea sediment (at 5246 m depth) of a station within the Pacific nodule province in the eastern tropical Pacific (Lian et al., 2007). Most of them belong to the Serratia-like chitinase and do not show similarity to any of our chitinase sequences. Unlike Uranouchi bay at 18 m depth or the offshore at 200 m depth in this study, most deep-sea areas are permanently cold region. Some clones from the Pacific nodule province are closely related to a chitinase gene from the lake sediment of Antarctic Ardley Island (Lian et al., 2007).This could indicate the adaptation to cold environment and this might why chitinase sequences from two different sediments are quite different.
CONCLUSION
We could isolate new and novel GH18 family of chitinase genes from marine sediments in the eutrophic inner bay, while the relatively common chitinase genes were obtained from the offshore sediments. The novel chitinase genes were isolated only from metagenomic DNA. These results indicate that the marine sediments used in this study contain a very diversity of chitinase genes and we are in progress to isolate full length of chitinase genes.
ACKNOWLEDGMENTS
This study was supported in part by The Kochi University President's Discretionary Grant to K.O.
REFERENCES
- Cauchie, H.M., 2002. Chitin production by arthropods in the hydrosphere. Hydrobiology, 470: 63-95.
CrossRef - Cottrell, M.T., D.N. Wood, L. Yu and D.L. Kirchman, 2000. Selected chitinase genes in cultured and uncultured marine bacteria in the alpha- and gamma-subclasses of the proteobacteria. Applied Environ. Microbiol., 66: 1195-1201.
Direct Link - Cottrell, MT., J.A. Moore and D.L. Kirchman, 1999. Chitinases from uncultured marine microorganisms. Applied Environ. Microbiol., 65: 2553-2557.
Direct Link - Fukamizo, T., 2000. Chitinolytic enzymes: Catalysis, substrate binding and their application. Curr. Protein Pept. Sci., 1: 105-124.
PubMedDirect Link - Gao, J., M.W. Bauer, K.R. Shockley, M.A. Pysz and R.M. Kelly, 2003. Growth of hyperthermophilic archaeon Pyrococcus furiosus on chitin involves two family 18 chitinases. Applied Environ. Microbiol., 69: 3119-3128.
CrossRef - Hjort, K., M. Bergstrom, M.F. Adesina, J.K. Jansson, K. Smalla and S. Sjoling, 2010. Chitinase genes revealed and compared in bacterial isolates, DNA extracts and a metagenomic library from a phytopathogen-suppressive soil. FEMS Microbiol. Ecol., 71: 197-207.
CrossRef - Hobel, C.F.V., V.T. Marteinsson, G.O. Hreggvidsson and J.K. Kristjansson, 2005. Investigation of the microbial ecology of intertidal hot springs by using diversity analysis of 16S rRNA and chitinase genes. Applied Environ. Microbiol., 71: 2771-2776.
CrossRef - Horn, S.J., A. Sorbotten, B. Synstad, P. Sikorski, M. Sorlie, K.M. Varum and V.G. Eijsink, 2006. Endo/exo mechanism and processivity of family 18 chitinases produced by Serratia marcescens. FEBS J., 273: 491-503.
Direct Link - Hoster, F., J.E. Schmitz and R. Danial, 2005. Enrichment of chitinolytic microorganisms: Isolation and characterization of chitinase exhibiting antifungal activity against phytopathogenic fungi from a novel Streptomyces strain. Applied Microbiol. Biotechnol., 66: 434-442.
Direct Link - Karlsson, M. and J. Stenlid, 2009. Evolution of family 18 glycoside hydrolases: Diversity, domain structures and phylogenetic relationships. J. Mol. Microbiol. Bitechnol., 16: 208-223.
CrossRef - LeCleir, G.R., A. Buchan and J.T. Hollibaugh, 2004. Chitinase gene sequences retrieved from diverse aquatic habitats reveal environment-specific distributions. Applied Environ. Microbiol., 70: 6977-6983.
CrossRef - Li, H. and L.H. Greene, 2010. Sequence and structural analysis of the chitinase insertion domain reveals two conserved motifs involved in chitin-binding. PLoS One, 5: e8654-e8654.
PubMed - Lian, M., S. Lin and R. Zeng, 2007. Chitinase gene diversity at a deep sea station of the east Pacific nodule province. Extremophiles, 11: 463-467.
PubMed - Makino, K., K. Oshima, K. Kurokawa, K. Yokoyama and T. Uda et al., 2003. Genome sequence of Vibrio parahaemolyticus: A pathogenic mechanism distinct from that of V cholerae. Lancet, 361: 743-749.
Direct Link - Metcalfe, A.C., M. Krsek, G.W. Gooday, J.I. Prosser and E.M.H. Wellington, 2002. Molecular analysis of a bacterial chitinolytic community in an upland pasture. Applied Environ. Microbiol., 68: 5042-5050.
CrossRef - Page, R.D.M., 1996. TreeView: An application to display phylogenetic trees on personal computers. Comput. Applied Biosci., 12: 357-358.
CrossRefPubMedDirect Link - Ramaiah, N., R.T. Hill, J. Chun, J. Ravel, M.H. Matte, W.L. Straube and R.R. Colwell, 2000. Use of a chiA probe for detection of chitinase genes in bacteria from the Chesapeake Bay (1). FEMS Microbiol. Ecol., 34: 63-71.
PubMedDirect Link - Snelgrove, P.V.R., 1999. Getting to the bottom of marine biodiversity: Sedimentary habitats: Ocean bottoms are the most widespread habitat on Earth and support high biodiversity and key ecosystem services. BioSciences, 49: 129-138.
Direct Link - Steele, H.L., K.E. Jaeger, R. Daniel and W.R. Streit, 2009. Advances in recovery of novel biocatalysts from metagenomes. J. Mol. Microbiol. Bitechnol., 16: 25-37.
CrossRefPubMedDirect Link - Suzuki, K., M. Taiyoji, N. Sugawara, N. Nikaidou and B. Henrissat et al., 1999. The third chitinase gene (chiC) of Serratia marcescens 2170 and the relationship of its product to other bacterial Chitinases. Biochem. J., 343: 587-596.
PubMed - Takenaka, T., T. Tashiro, A. Ozaki, H. Takakubo and Y. Yamamoto et al., 2007. Planktonic bacterial population dynamics with environmental changes in coastal areas of Suruga Bay. Microbes Environ., 22: 257-267.
CrossRef - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Tsujibo, H., T. Kubota, M. Yamamoto, K. Miyamoto and Y. Inamori, 2003. Characterization of chitinase genes from an alkaliphilic actinomycete, Nocardiopsis prasina OPC-131. Applied Environ. Microbiol., 69: 894-900.
CrossRef - Van Alten, D.M.F., B. Synstad, M.B. Brurberg, E. Hough, B.W. Riise, V.G.H. Eijsink and R.K. Wierenga, 2000. Structure of a two-domain chitotriosidase from Serratia marcescens at 1.9 � resolution. Proc. Natl. Acad. Sci. USA., 97: 5842-5847.
Direct Link - Wang, Q., G.M. Garrity, J.M. Tiedje and J.R. Cole, 2007. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Applied Environ. Microbiol., 73: 5261-5267.
CrossRefPubMedDirect Link - Ikeda, S., N. Ytow, H. Ezura, K. Minamisawa, K. Miyashita and T. Fujimura, 2007. Analysis of molecular diversity of bacterial chitinase genes in the maize rhizosphere using culture-independent methods. Microbes. Environ., 22: 71-77.
Direct Link - Tanaka, T., S. Fujiwara, S. Nishikori, T. Fukui, M. Takagi and T. Imanaka, 1999. A unique chitinase with dual active sites and triple substrate binding sites from the hyperthermophilic archaeon Pyrococcus kodakaraensis KOD1. Applied Environ, Microbiol., 65: 5338-5344.
Direct Link