
M. Mortazavi
Department of Pilot Biotechnology, Pasteur institute of Iran, Tehran, Iran
A. Akbarzadeh
Department of Pilot Biotechnology, Pasteur institute of Iran, Tehran, Iran
A. Farhangi
Department of Pilot Biotechnology, Pasteur institute of Iran, Tehran, Iran
M. Mehrabi
Department of Pilot Biotechnology, Pasteur institute of Iran, Tehran, Iran
Z. Hosseinian
Department of Pilot Biotechnology, Pasteur institute of Iran, Tehran, Iran
M. J. Mokhtari
Department of Pilot Biotechnology, Pasteur institute of Iran, Tehran, Iran
Z. Saffari
Department of Pilot Biotechnology, Pasteur institute of Iran, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2011 | Volume: 14 | Issue: 2 | Page No.: 128-132







ABSTRACT
The aim of this study was to investigate the antiproliferative proteins that probably have a role in Helicobacter pylori evade of immune response and cause chronic infection disease and also to see if coccoid form had a role in its chronicity. H. pylori strain VacA s2/m2 positive and CagA negative, from a gastric biopsy of a patient with peptic ulcer disease, was isolated and cultured in brucella agar. Both spiral and coccoid forms were harvested and ruptured by sonication. The cytoplasmic solutions of both forms were collected and their fractions obtained by gel chromatography and preparative polyacrylamide gel electrophoresis. The fractions were analyzed by MTT assay for their antiproliferative activity. We isolated two proteins with a significant dose dependent antiproliferative activity that analyzed with sodium dodecyl sulfate-polyacrylamide gel electrophoresis, one of them that was urease positive showed two bands with 61 and 27 kDa, which is resumed to urease of H. pylori, another consist of 57 and 63 kDa. Helicobacter pylori secret some proteins like urease that inhibit immune cells proliferation response against its antigens.
PDF Abstract XML References Citation
Received: November 04, 2010;
Accepted: January 11, 2011;
Published: February 09, 2011
How to cite this article
M. Mortazavi, A. Akbarzadeh, A. Farhangi, M. Mehrabi, Z. Hosseinian, M. J. Mokhtari and Z. Saffari, 2011. Immunosuppressive Proteins Isolated from Spiral and Coccoid Cytoplasmic Solutions of Helicobacter pylori. Pakistan Journal of Biological Sciences, 14: 128-132.
DOI: 10.3923/pjbs.2011.128.132
URL: https://scialert.net/abstract/?doi=pjbs.2011.128.132
DOI: 10.3923/pjbs.2011.128.132
URL: https://scialert.net/abstract/?doi=pjbs.2011.128.132
INTRODUCTION
Helicobacter pylori is a gram negative and spiral bacterium that has infected more than 50% of world’s population. H. pylori infection is related to gastric ulcer and MALT lymphoma (Kimmel et al., 2000).
Although, H. pylori are noninvasive bacteria, it induces chronic infection (Marshall, 2002). H. pylori infection is almost asymptomatic and less than 10% of the infected individuals have ulcer or gastric associated diseases. These patients are in high risk of stomach cancer. Relationship between H. pylori infection and gastric cancer is so significant that World Health Organization (WHO) has organized H. pylori as a type I carcinogen. It is postulated that long-term inflammation induced by H. pylori has an important role in gastric cancer development (Bah et al., 2001).
Helicobacter pylori releases factors that are responsible for colonization and pathogenesis (Franco et al., 2008). It has been shown by Ibraghimov and Jacques (2000) that H. pylori secrets a protein that inhibits epithelial and lymphocyte cell lines proliferation and that its effects is dose dependent. It was also shown that urease, VacA and CagA have cytotoxic effects on cells. In the stomach, mostly spiral forms of H. pylori were found and coccoid forms were in the more damaged regions of gastric mucosa (Saito et al., 2003).
It has been shown that chronic inflammation promotes cancer (Shacter et al., 2002). Despite local and systemic response to H. pylori infection (Bhat et al., 2005), H. pylori induces chronic gastric inflammation. Epidemiological studies have shown that gastric cancer and MALT lymphoma are related to H. pylori infection (Feldman, 2001). H. pylori is acquired in childhood (Mitchell et al., 2003), but gastric cancer is seen mostly after the six decade of life. It is postulated that chronic infection with H. pylori is necessary for gastric cancer (Feldman, 2001).
-It has been shown that lymphocytes from H. pylori, that infected individuals sometimes shows a decrease in proliferation, in response to helicobacter antigens in vitro. This had revealed that H. pylori might down regulate the cellular immune responses toward itself (Windle et al., 2005). It is possible that H. pylori secrets proteins that inhibit immune response to eradicate the infection locally (Ibraghimov and Jacques, 2000). The aim of this study was to isolate the cytoplasmic proteins of H. pylori responsible for the inhibition of T cells proliferation.
In this study, we isolated different fractions of both spiral and coccoid forms of H. pylori and evaluated their antiproliferative activity.
MATERIALS AND METHODS
Bacterial isolation and culture conditions: The H. pylori strain was isolated from gastric biopsy of a patient suffering from a gastric ulcer. Present study was performed in 2006 and it took 7 mounts. The cells were grown on brucella agar (Merck, Germany) supplemented with 10% sheep blood, vancomycin (10 mg mL-1), nystatin (1 mg mL-1) and trimethoprim (5 mg mL-1) and incubated for 48 h in CO2 incubator (10% CO2, 5% N2 and 85% O2) (Ramarao and Meyer, 2001). Bacterial cells were harvested after the second and 13th days of incubation and suspended in PBS (pH 7.2). To assess the relative percentages of coccoid and bacillary forms, Gram-staining was used for each suspension. The Gram staining showed that the suspension of 2nd day culture, contained only spiral forms and 13th day culture contained coccoid forms.
Preparation of cytoplasmic solutions: The cells were collected and washed three times with 20 mL of PBS. The packed cells were frozen at -80°C before use. The cells were sonicated on ice (eight times for 30 sec each time). Unbroken cells and debris were precipitated by centrifuge at 4,000 x g for 10 min at 4°C. The supernatants were collected and centrifuged at 45,000 x g for 20 min at 4°C. The pellets contained cell walls with outer membrane protein (Ge et al., 2001) and the supernatants contained cytoplasmic proteins.
Chromatography: The cytoplasmic solutions obtained from spiral and coccoid forms were concentrated with freeze-drying and then dialyzed against PBS for three times. The concentrated solutions of spiral and coccoid forms (7 and 3 mg mL-1) were applied in gel filtration column using S-200 and eluted with sodium phosphate buffered saline, pH 7.2, (0.02 M sodium phosphate, 0.15 M NaCl). Optical density was measured at 280 nm and fractions with maximum OD were pooled.
Preparative PAGE: Fractions with antiproliferative activity obtained by chromatography were used in preparative PAGE to isolate proteins with antiproliferative effect. A piece of the gel was cut from length and stained by silver stain. According to the stained gel, the protein bands were cut from wide of the remained gel and cut into pieces in eluting solution (0.1 M Tric-HCl; pH 7) and then centrifuged at 3200 x g for 15 min after shaking. The supernatants were collected and stored at 4°C for 30 min. Proteins were precipitated by acetone and the solution was stirred magnetically for 15 min at 20°C. After being centrifuged at 3200 x g, the acetone was removed and the proteins were diluted in PBS. The isolated proteins were dialyzed against PBS and prepared for proliferation test. The concentration of proteins was determined by Bradford assay (Kruger, 2002). The presence of urease activity in the stomach biopsies and the isolated proteins was determined with a Christensen urease medium. A rapid color change suggests the presence of urease activity.
Cell viability assay: The antiproliferative effect of the fractions on Jurkat cell lines, a CD4+ leukemia T-cell line, were determined by MTT assay described by Young et al. (2005). The cells were maintained in the culture medium in 10 mL culture flasks at 37°C in a 7% CO2 humidified atmosphere. Cell viability was assessed by trypan blue and the exclusion was over 90%. The cells were seeded in 96-well microtiter plates at a density of approximately 4x106 cells/well containing 100 μL RPMI 1640 (Gibco, UK), 10% heat-inactivated Fetal Calf Serum (FCS), 100 U L-1 penicillin and 100 mg L-1 streptomycin (Gibco, Scotland). Each fraction was added in various concentrations and some wells were leaved without samples as controls. All MTT assays were measured in triplicate. 10 μL of the MTT (3-[4, 5-dimethylthiazol-2-yl]-2, 5-diphenyltetrazolium-bromide, 5 mg mL-1; Sigma) solution was added to each well and incubated for 4 h until a purple precipitate was visible. The developed crystals were dissolved in 100 μL of 0.04 M HCl in isopropanol and the OD was read in a microplate spectrophotometer at a wavelength of 570 nm.
The stimulation index was calculated by dividing OD at 570 nm with sample by OD at 570 nm without samples. Student's t-test was utilized for the statistical analysis of the samples. Differences between the treatment means were considered statistically significant if p<0.05.
RESULTS AND DISCUSSION
Isolation of antiproliferative proteins from cytoplasmic solutions: After chromatography of cytoplasmic solutions, various fractions were pooled as S1, S2 and S3 for spiral forms and C1, C2 and C3 for coccoid forms (Fig. 1a, b). The fractions with antiproliferative activity (S2 and C2) were applied in preparative PAGE for isolation of proteins with antiproliferative activity. The preparative PAGE (Fig. 2) was cut in 6 pieces (P1, P2, P3, P4, P5 and P6) from length in both S2 and C2. P5 band was not seen in preparative PAGE of C2. Both P4 and P5 consisted of one band in preparative PAGE. The P5 from S2 with urease activity showed 61 and 27.5 kDa bands in SDS-PAGE, P4 showed two bands of 63 and 57 kDa (Fig. 3a, b).
Antiprolifrative activity of the isolated proteins: After gel chromatography, the fractions were analyzed by MTT assay for their antiproliferative activity (Fig. 4a). The cells treated with S2 and C2 showed lower OD than negative controls (p<0.05). No statistical difference was observed between other fractions.
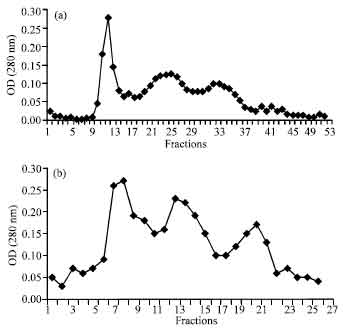 | |
| Fig. 1: | Cytoplasmic solutions of (a) spiral and (b) coccoid forms were applied in gel filtration column, using S-200. The 1-17, 18-30 and 31-56 fractions were pooled as S1, S2 and S3 for Spiral forms and 1-11, 12-17 18-23 as C1, C2 and C3 for coccoid forms, respectively |
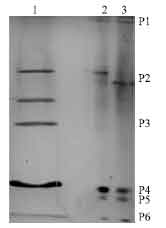 | |
| Fig. 2: | PAGE analysis of spiral and coccid forms fractions 2, S2 and C2, obtained by gel chromatography. Lane 1. C2, Lane 2 and 3. S2. The preparative PAGE of S2 and C2 were cut in 6 pieces (P1-P6), from its wide |
The P1 to P6 fractions obtained by preparative PAGE were analyzed by MTT assay (Fig. 4b). The P4 and P5 fractions showed significant dose dependent antiproliferative activity on Jurkat cells.
CagA, VacA and urease are antiproliferative proteins (Takagi et al., 2000) but VacA s2/m2 does not have the cytotoxic effect that is due to defect in N-terminal of this protein. For inhibiting the interference of CagA and VacA cytotoxic effect the selected strain was urease positive, VacA s2/m2 positive and CagA negative.
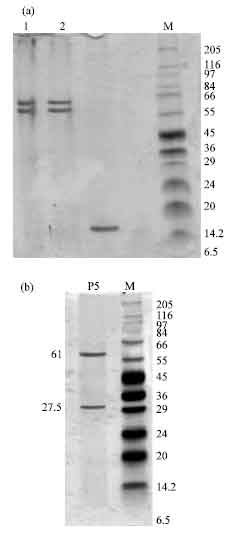 | |
| Fig. 3: | SDS-PAGE analysis of fractions isolated from preparative PAGE (a) Lane 1. P4 isolated from S2, lane 2. P4 isolated from C2, (b) lane 1. P5 isolated from preparative PAGE of S2 |
Spiral forms had two proteins with antiproliferative activity (p<0.05): P5 with urease activity in SDS-PAGE showed two bands, 61 and 27.5 kDa, the other (P4) showed two bands of 63 and 57 kDa. It is likely that P5 protein and its subunits that were shown in SDS-PAGE are similar to urease protein of H. pylori (Figueroa et al., 2002). It has been shown that in coccoid forms, urease mRNA does not exist (Bah et al., 2001). We have shown that in coccoid forms, only P4 was expressed and urease was not expressed.
The isolated protein (P4) showed a dose dependent antiproliferative activity that is necessary be cloned for more recognition and sequencing.
Relation between H. pylori and gastric cancer is so certain that WHO has classified H. pylori in class I carcinogens (Romero-Gallo et al., 2008). Chronic inflammation alone is not sufficient to induce gastric cancer.
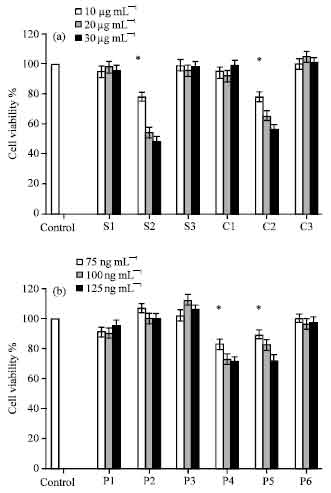 | |
| Fig. 4: | In vitro influence of different concentrations of H. pylori fractions on proliferation of Jurkat T cell was estimated by MTT assay. Experiments were performed in triplicate and at least 3 separate experiments, the Means±SD of the values are indicated. (A) S1, S2 and S3 from spiral forms and C1, C2 and C3 from coccoid forms, obtained by gel filtration (10, 20 and 30 μg mL-1) and (B) P1, P2, P3, P4, P5 and P6 obtained by preparative PAGE (75, 100, 125 ng mL-1) were utilized in MTT assay. Untreated control cells were given the value of 1 (100% SI). *Indicates a significant difference in comparison with control as shown by Student t-test, at p<0.05 |
H. pylori requires a direct effect of such mutagens to induce tumorogenes. H. pylori induces progression of carcinoma, but it may have direct effect on gastric to induce carcinoma.
Helicobacter pylori has a role in development of gastric carcinoma. It is postulated that H. pylori induces mutation in cells and inhibits proliferation of immune cells and also induces initiation and progression of carcinoma (Leon-Barua et al., 2006).
CONCLUSION
In this study it was shown that both spiral and coccoid forms of H. Pylori have two antiproliferative proteins that one of them had urease activity and the other showed two bands of 63 and 57 kDa. It is recommended to evaluate the effect of the unknown protein on immune response since it perhaps have a role in the chronic infection and also stomach cancer.
ACKNOWLEDGMENTS
The financial support by the Research Council of Pasteur institute of Iran is gratefully acknowledged.
REFERENCES
- Franco, A.T., E. Johnston, U. Krishna, Y. Yamaoka and D.A. Israel et al., 2008. Regulation of gastric carcinogenesis by Helicobacter pylori virulence factors. Cancer Res., 68: 379-387.
Direct Link - Ibraghimov, A. and P. Jacques, 2000. The immune response against Helicobacter pylori: A direct linkage to the development of gastroduodenal disease. Microbes Infect., 2: 1073-1077.
PubMed - Saito, N., K. Konishi, F. Sato, M. Kato, H. Takeda, T. Sugiyama and M. Asaka, 2003. Plural transformation-processes from spiral to coccoid Helicobacter pylori and its viability. J. Infect., 46: 49-55.
PubMed - Shacter, E. and S.A. Weitzman, 2002. Chronic inflammation and cancer. Onchology, 16: 217-226.
Direct Link - Bhat, N., J. Gaensbauer, R.M. Peek, K. Bloch, K.T. Tham, M.J. Blaser and G. Perez-Perez, 2005. Local and systemic immune and inflammatory responses to Helicobacter pylori strains. Clin. Diagnostic Lab. Immunol., 12: 1393-1400.
CrossRef - Feldman, R.A., 2001. Would eradication of Helicobacter pylori infection reduce the risk of gastric cancer. Alimentary Pharmacol. Therapeutics, 15: 2-5.
PubMed - Mitchell, A., M.J.S. Terezinha, J.B. Leah, A.M.L. Aldo and L.G. Richard, 2003. Age-specific Helicobacter pylori seropositivity rates of children in an impoverished urban Area of Northeast Brazil. J. Clin. Microbiol., 41: 1326-1328.
CrossRef - Windle, H.J., Y.S. Ang, V.A. Morales, R. McManus and D. Kelleher, 2005. Human peripheral and gastric lymphocyte responses to Helicobacter pylori NapA and AphC differ in infected and uninfected individuals. Gut, 54: 25-32.
CrossRef - Ramarao, N. and T.F. Meyer, 2001. Helicobacter pylori resists phagocytosis by macrophages: Quantitive assessment by confocal microscopy and fluorescence-activated cell sorting. Infect. Immun., 69: 2604-2611.
Direct Link - Ge, Z., P. Doig and J.G. Fox, 2001. Characterization of proteins in the outer membrane preparation of a murine pathogen, Helicobacter bilis. Infect. Immun., 69: 3502-3506.
CrossRef - Young, F.M., W. Phungtamdet and B.J.S. Sanderson, 2005. Modification of MTT assay conditions to examine the cytotoxic effects of amitraz on the human lymphoblastoid cell line, WIL2NS. Toxicol. In vitro, 19: 1051-1059.
PubMed - Takagi, A., S. Watanabe, M. Igarashi, J. Koike and K. Hasumi et al., 2000. The effect of Helicobacter pylori on cell proliferation and apoptosis in gastric epithelial cell lines. Aliment. Pharmacol. Ther., 14: 188-192.
PubMed - Figueroa, G., G. Faundez, M. Troncoso, P. Navarrete and M.S. Toledo, 2002. Immunoglobulin G antibody response to infection with coccoid forms of Helicobacter pylori. Clin. Diagn. Lab. Immunol., 9: 1067-1071.
CrossRef - Romero-Gallo, J., E.J. Harris, U. Krishna, M.K. Washington, G.I. Perez-Perez and Jr. R.M. Peek, 2008. Effect of Helicobacter pylori eradication on gastric carcinogenesis. Lab Invest., 88: 328-336.
PubMed - Leon-Barua, R., S. Recavarren-Arce, E. Chinga-Alayo, C. Rodriguez-Ulloa and D.N. Taylor et al., 2006. Helicobacter pylori-associated chronic atrophic gastritis involving the gastric body and severe disease by Vibrio cholerae. Trans. Roy Soc. Trop. Med. Hyg., 100: 567-572.
Direct Link