
S. K. Shahani
University of Liverpool School of Veterinary Science, Leahurst Campus, Chester High Road, Neston. CH64 7TE, UK
S. G. Revell
Genus Breeding Limited, Freezing Unit, Llanrhydd, Ruthin, UK
C. G. Argo
University of Liverpool School of Veterinary Science, Leahurst Campus, Chester High Road, Neston. CH64 7TE, UK
R. D. Murray
University of Liverpool School of Veterinary Science, Leahurst Campus, Chester High Road, Neston. CH64 7TE, UK
Pakistan Journal of Biological Sciences
Year: 2010 | Volume: 13 | Issue: 16 | Page No.: 802-808







ABSTRACT
Recently positive correlation has been found between oxygen consumption (ZO2) in bull spermatozoa and non-return rates and concluded that an increase in ZO2, characteristic of the freeze/thaw process, was possibly associated with mitochondrial membrane damage during this procedure: alternatively, sperm may be hyperactivated through the capacitation-like effects of freezing/thawing. We speculated that the morphology of spermatozoa may be associated with their rate of ZO2 and fertility: for example, sperm mid-piece length where mitochondria are located. Such a relationship has not been investigated before, particularly in context of commercial cattle breeding programmes and bull fertility characteristics. Sperm biometry was performed on ejaculates obtained from 34 bulls representing six breeds: Holstein (yearlings and mature), Friesian, Belgian Blue, Aberdeen Angus, Charolais and Limousin. Five ejaculates were collected from every bull and from each sample a semen smear was fixed and stained with eosin/nigrosin: the mid-piece length of 40 sperm with normal morphology was measured in every sample. Data were analysed by breed, age and within each bull. Significant differences (p<0.01) between ejaculates in 9/34 bulls was found, as well as differences (p<0.001) between individual bulls within the same breed. The average mid-piece length for Aberdeen Angus was 13.35 μm, for Belgian Blues and Limousin around 13.8 μm and for Charolais 13.68 μm: for dairy breeds such as Holstein and Friesian it was about 13.4 μm. The mean value of mid-piece length for breed was compared with their 49 day non-return rate; a negative correlation (r = -0.53) was found in black and white dairy breeds.
PDF Abstract XML References Citation
Received: March 17, 2010;
Accepted: July 22, 2010;
Published: August 18, 2010
How to cite this article
S. K. Shahani, S. G. Revell, C. G. Argo and R. D. Murray, 2010. Mid-piece Length of Spermatozoa in Different Cattle Breeds and its Relationship to Fertility. Pakistan Journal of Biological Sciences, 13: 802-808.
DOI: 10.3923/pjbs.2010.802.808
URL: https://scialert.net/abstract/?doi=pjbs.2010.802.808
DOI: 10.3923/pjbs.2010.802.808
URL: https://scialert.net/abstract/?doi=pjbs.2010.802.808
INTRODUCTION
Sperm are highly differentiated, specialised cells that transfer a haploid set of paternal chromosomes to the female gamete at fertilisation. To achieve this, they must traverse the length of the female genital tract, bind to oviduct epithelium to undergo capacitation, attach to the zona pellucida and finally penetrate the oocyte itself (Foote, 2003).
Spermatozoa obtain energy for their motility through glycolysis and/or respiration, provided that appropriate substrate is available (Krzyzosiak et al., 1999). Rikmenspoel (1965) estimated that 70% of Adenosine Tri-Phosphate (ATP) produced by bovine spermatozoa is used for motility, the precise proportion being related to the incubation temperature: for example, at 18 and 37°C around 9 and 58% of sperm respectively showed progressive motility (Hammerstedt and Hay, 1980). Spermatozoa generate most of their energy by glycolysis rather than oxidative phosphorylation and a number of sperm-specific enzymes involved in the glycolytic pathways are located in the fibrous sheath (Miki et al., 2004). Mitochondria that produce ATP through oxidative phosphorylation are located in the sperm mid-piece, found at the extreme anterior end of the flagellum (Turner, 2003; Ford, 2006). This function is crucial because it correlates with the energy status of spermatozoa, their motility and potential for achieving successful fertilisation (Casey et al., 1993; Kasai et al., 2002).Hyperactivated motility in sperm is an essential precursor to successful fertilization that requires both increased ATP synthesis and immediate availability within mitochondria (Suarez and Ho, 2003; Quill et al., 2003). Their flagellar movements become asymmetrical and whip-like, have high amplitude and low frequency and this facilitates their detachment from the oviduct epithelium to position sperm in the ampulla prior to fertilization (Turner, 2003).
Biometric differences in sperm morphology between mammalian species were described by Cummins and Woodall (1985), who measured lengths of mid-piece, caput or head and principal piece for most species except primates and bats. Gage (1998) found a correlation between mid-piece and flagellar lengths for the 179 species he investigated. More recently, Aggarwal et al. (2007) observed a wide variation in the length of the mid-piece between buffalo bulls of different breeds and suggested that sperm biometric characteristics might be associated with male fertility. It is in this context that sperm competition has been considered in those species where two or more males compete for opportunities to mate with a single female, such as in canines and fertilize ova. Sperm mid-piece volume may be indicative of a higher mitochondrial content and is significantly larger in these species (Anderson et al., 2005): there is also a positive correlation between it and testis size. Mid-piece volume is correlated positively with motility in domestic fowl, possibly dependent upon an endogenous substrate (Froman and Feltmann, 1998) and in primates that exhibit multiple-partner mating activity (Anderson and Dixson, 2002; Dixson et al., 2004). Generally, sperm with longer flagella tend to swim faster to reach the ova sooner and achieve fertilization compared to those with shorter tails when ejaculates from different males intermingle in the same female tract (Gomendio and Roldan, 1991): there is a positive correlation between sperm length and its maximum velocity. By contrast, sperm from the Iberian red deer stag have long mid-pieces but swim relatively slowly (Malo et al., 2006), suggesting that their activated spermatozoa with shorted mid-pieces may exhibit greater motility.
Several methods have been used to assess semen quality, using laboratory parameters to predict the fertilization potential of bulls and correlating these results with actual pregnancy rates achieved in the field. Post thaw motility is assessed most frequently as a measure of sperm viability and is correlated (r = 0.4-0.5) with 56 day non-return rates in cows (Januskauskas et al., 2003). Also used are the percentage of viable spermatozoa present in an ejaculate, computer-based estimates of motility characteristics and acrosome integrity that are all correlated significantly with good fertility (Budworth et al., 1988; Kjaestad et al., 1993; Zhang et al., 1998; Januskauskas et al., 2003). Hence, bulls with apparently normal semen that are subfertile are worthy of further investigation using more recently devised molecular techniques (Aggarwal et al., 2007).
Froman and Feltmann (1998) and Froman et al. (1999) concluded that mitochondrial function and ATP content of avian sperm were the most important factors affecting their motility and sperm motility was the primary determinant for optimum fertility. For example, the mean rate of oxygen consumption was 1.14 μL O2 min-1 in fresh semen ejaculates of high motility compared with 0.62 μL O2 min-1 in those presenting only moderate motility. Recently, Garrett et al. (2008) found a positive correlation between oxygen consumption (ZO2) in bull spermatozoa and non-return rates and concluded that an increase in ZO2, characteristic of the freeze/thaw process, was possibly associated with mitochondrial membrane damage during this procedure: alternatively, sperm may be hyperactivated through the capacitation-like effects of freezing/thawing.
We speculate that the morphology of spermatozoa may be associated with their rate of ZO2 and fertility: for example, sperm mid-piece length where mitochondria are located. Such a relationship has not been investigated before, particularly in context of commercial cattle breeding programmes and bull fertility characteristics. This present study investigates variation in sperm mid-piece biometry by breed, age and between bulls used in current breeding programmes for dairy and beef herds in both intensive and semi-intensive cattle livestock farms, worldwide.
MATERIALS AND METHODS
Six different breeds were chosen during 2009 for investigation. Holstein and Friesian represented the dairy breeds and Belgian Blue, Aberdeen Angus, Charolais and Limousin the beef breeds used commonly as terminal sires. Five ejaculates were collected and examined from each bull. To prepare semen samples for microscopic examination, 100 μL of raw semen was added to 250 μL of Eosin Nigrosin stain and, after shaking well, drops of stained semen were placed at one end of a clean microscopic glass slide and a smear made by drawing them out with the clean edge of another glass slide. The slides were labelled with bull identity and date and allowed to dry at room temperature. The mid-piece length of forty morphologically normal sperm per ejaculate was measured, selected from various fields at random on each of five slides, using a x100 oil immersion objective with a Nikon Labophot 2 Microscope fitted with a digital CCD camera (Penguin-600 CL-CU) with imaging software within the Penguin/Pro application suite (Penguin/Pro Application Suite v.3.5.0, Pixera corporation USA).
Sperm with abnormal morphology of either head or tail and post-ejaculatory injury were excluded so that only those with normal morphology and a distinct mid-piece, either straight or curved, were measured (Fig. 1).
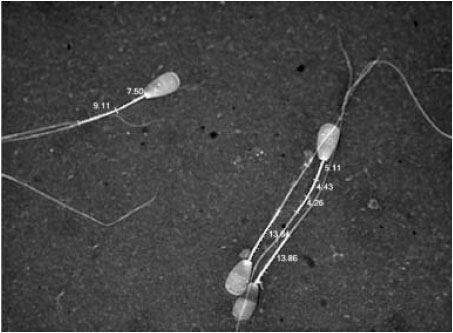 | |
| Fig. 1: | Spermatozoa with both straight and bent mid-pieces that were measured either as a single straight line or the sum of several measurements |
Before taking the measurement, the captured digital image of the individual spermatozoan was magnified to a level where the mid-piece could be distinguished clearly from the principal piece. Measurements (in microns) were made of those sperm with straight flagellum by dragging the cursor from the distal point of the mid-piece to its anterior end at the base of head region: where the flagella were bent, mid-pieces were measured using a series of short straight lines that described their curve, the sum of these being considered as a single value.
Data and statistical analysis: The estimated mid-piece lengths were recorded and organized into Excel files with the independent variables being breed and ejaculate number. After cleaning, the data were investigated using one-way Analysis of Variance (ANOVA) in Minitab; relationships between independent variables were investigated using linear correlation coefficient. Significance was accepted at a p-value <0.05 and results presented as Mean±SD, except where stated.
RESULTS
Differences between ejaculates from the same bull: The mid-piece length was compared between young Holstein bulls less than two years old with older, mature animals (Fig. 2a-c). Nine of the 34 bulls examined (31%) showed significant variation between their own ejaculates: four Holsteins (F = 2.48, p<0.05: F = 3.75, p<0.01), one Friesian (F = 2.92, p<0.05), two Belgian Blues (F = 2.98, p<0.05), one Aberdeen Angus (F = 2.54, p<0.05) and a Limousin (F = 2.71, p<0.05). The ejaculates obtained from Friesian bulls showed least variation between collections whereas one Holstein bull (AHO2) showed just over 2% variation in mid-piece length (F = 5.89, p<0.01).
Differences between breeds: There were significant differences (p<0.001) between bulls within the same breed: for example, between juvenile Holstein bull 2 and the other four within that group and two Belgian Blue bulls - 2 and 5-that had significantly shorter mid-pieces than the other three (Table 1). Despite this intra-breed variation, there was no significant difference in mid-piece length between the breeds. Sperm from Belgian Blue and Limousin bulls presented with mid-pieces of similar length that were longer than the other breeds investigated: those with the shortest length were from juvenile Holstein bulls.
Mid-piece length and 49-day non-return rates: Data were analysed from 5850 individual cow breeding records that were served using semen collected from the five mature Holstein and five Friesian bulls evaluated in this study. The overall 49-day non-return rate for the Holstein bulls was 73% and for Friesians 80%. However, within this dairy bull sample the non-return rate was lower by 5% from the breed average when mid-piece length was significantly longer (r = -0.53): the small numbers of bulls involved prevented meaningful statistical relationships being established.
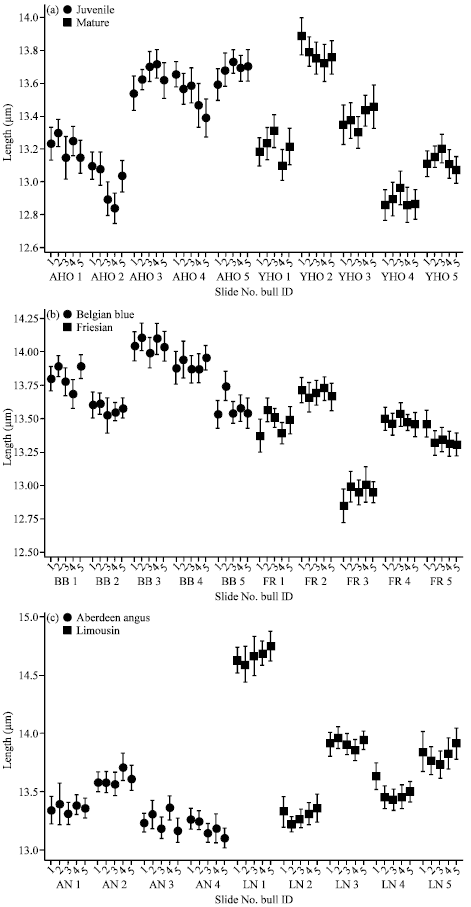 | |
| Fig. 2: | Variation in sperm mid-piece length between ejaculates within five different bull breeds: (a) Holsteins juvenile and Holsteins mature (b) Belgian Blue and Friesian and (c) Aberdeen Angus and Limousin |
| Table 1: | Mean mid-piece length (SD) of 200 spermatazoa examined from five ejaculates, each obtained from either juvenile and mature Holstein or five other breeds of bull |
 | |
DISCUSSION
Variations in the length of mammalian spermatozoa are due mostly to variation in tail length (Cummins and Woodall, 1985) and the size of the mid-piece contributes to such species differences. Anderson and Dixson (2002) found that mid-piece volume was greater in primates whose normal mating patterns involved multiple rather than single partners. Earlier, Gomendio and Roldan (1991) had found similar differences between monoandrous and polyandrous species: in rodent and other species where males face sperm competition after mating with a single female, spermatozoa are longer than in species where only a single dominant male will mate with a female e.g. some wild cats and deer. The longer the sperm flagellum, the greater the forces generated by its motions (Katz and Drobins, 1990) and its requirement for immediately available energy. This present study is the first to record variation in mid-piece length of a single species, the bovine, associated with collecting their ejaculates over a period of time. Whilst no significant differences were found between different breeds of bull, Aggarwal et al. (2007) found such variation within eight various breeds of buffalo. Results from this study showed significant variation in mid-piece length between ejaculates collected from nine of the 34 bulls investigated. The mid-piece lengths we obtained were similar to those found by Venkataswami and Vedanayagam (1962) who found an average mid-piece length of 12.41±0.85 μm for buffalo and 14.19±1.57 μm for domestic cattle. It is unlikely that the variation found in this present study occurred because of measuring technique since spermatozoa with straight tails could be measured as one entity and sperm with severely bent tails, due to cold shock, were positively excluded. In any case, the SD in this present study was ten-fold less than that recorded by the Indian researchers, suggesting this current computer-aided technique is robust. However, no attempt was made biochemically to distinguish between spermatozoa that were either non-apoptotic or undergoing programmed cell death: mid-piece morphology and mitochondrial volume is affected by apoptosis (Hendricks and Hansen, 2009) and this may have contributed to some variation found in this study.
Age influences mid- piece length in bulls. Cummins and Woodall (1985) found that sperm of bulls over 10 years old possess mid-pieces around 4% longer than younger fully mature males. In this present study, Holstein bulls under 18 months old had mid-pieces 1% shorter than older males, suggesting a slow, progressive lengthening of the mid-piece from puberty through to old age. Male reproductive potential generally falls with increasing age, associated with two factors. First, there is greater opportunity for mutations to adversely affect the mitochondrial genome and impair oxidative phosphorylation. The mid-piece is crucial for capacitation that involves tyrosine phosphorylated proteins, two of which are phospholipid hydroperoxide glutamate peroxidise and ATP synthase beta subunit: both have a primary role in sperm motility (Shivaji et al., 2009) and disruption of their synthesis will reduce male fertility. Second, mitochondria are a source of Reactive Oxygen Species (ROS) which is a normal product of cellular respiration: normally these are removed from cells by antioxidant defence mechanisms. In sperm, ROS can accumulate within the mid-piece and damage mitochondria: the likelihood of this occurring increases as males grow older (Desai et al., 2010). Further preliminary data from this present study associated Non-Return Rates (NRRs) in cows bred with semen collected from specific bulls whose mid-piece lengths were measured: longer mid-pieces correlated with up to a 5% reduction in NRR in the dairy bulls investigated. The relationship between mid-piece length, its mitochondrial volume and the efficiency of oxidative phosphylation should be investigated further.
There are several other factors that affect mid-piece length in bulls. For example, the mid-piece length of mature spermatozoa in pure-bred black-and-white cattle is 2% longer than in crossbred Friesian x Sahiwal animals (Sarder, 2005). Heterosis is well known as a positive influence for reproductive efficiency in cattle, particularly in reducing embryonic mortality (Heins et al., 2008) but the contribution of the male gamete to such improved fertility is unknown. Also, sperm from unimproved Indo-Pak zebu breeds tend to have mid-pieces 2-9% longer than European cattle (Venkataswami and Vedanayagam, 1962): again, the significance of this in terms of breeding efficiency is not known.
This study has described differences in spermatozoa mid-piece length in different ejaculates collected from the same bull and variation between breeds. These preliminary results suggest that further work is necessary to relate mid-piece length with mitochondrial density and volume and energy metabolism related to sperm motility, capacitation and fertility potential of known beef and dairy bulls.
ACKNOWLEDGMENTS
The author is grateful to the Livestock and Fisheries Department, Government of Sindh, Pakistan for financial support. The author also thankful to Mr. Ghulam Sarwar Shaikh, Director General, Livestock, Sindh and Mr. Abdul Qadir Junejo, Animal Breeding Division, Hyderabad for their keen interest and advice.
REFERENCES
- Aggarwal, R.A., S.P. Ahlawat, Y. Kumar, P.S. Panwar, K. Singh and M. Bhargava, 2007. Biometry of frozen-thawed sperm from eight breeds of Indian buffaloes (Bubalus bubalis). Theriogenology, 68: 682-686.
Direct Link - Anderson, M.J., J. Nyholt and A.F. Dixson, 2005. Sperm competition and the evolution of sperm midpiece volume in mammals. J. Zool., 267: 135-142.
CrossRef - Budworth, P.R., R.P. Amann and P.L. Chapman, 1988. Relationships between computerized measurements of motion of frozen thawed bull spermatozoa and fertility. J. Androl., 9: 41-54.
PubMed - Casey, P.J., R.B. Hillman, K.R. Robertson, A.I. Yudin, I.K. Liu and E.Z. Drobnis, 1993. Validation of an acrosomal stain for equine sperm that differentiates between living and dead sperm. J. Androl., 14: 289-297.
PubMed - Cummins, J.M. and P.F. Woodall, 1985. On mammalian sperm dimensions. J. Reproduct. Fertil., 75: 153-175.
PubMed - Dixson, A.F., J. Nyholt and M.J. Anderson, 2004. A positive relationship between baculum length and prolonged intromission patterns in mammals. Acta. Zool. Sinica., 50: 490-503.
CrossRef - Foote, R.H., 2003. Fertility estimation: a review of past experience and future prospects. Anim. Reproduct. Sci., 75: 119-139.
PubMed - Ford, W.C.L., 2006. Glycolysis and sperm motility: Does a spoonful of sugar help the flagellum go round?. Human Reproduct. Update, 12: 269-274.
CrossRef - Froman, D.P and A.J. Feltmann, 1998. Sperm mobility: A quantitative trait of the domestic fowl (Gallus domesticus). Biol. Reprod., 58: 379-384.
Direct Link - Froman, D.P., A.J. Feltmann, M.L. Rhoads and J.D. Kirby, 1999. Sperm mobility: A primary determinant of fertility in the domestic fowl (Gallus domesticus). Biol. Reprod., 61: 400-405.
CrossRefPubMedDirect Link - Gomendio, M. and E.R.S. Roldan, 1991. Sperm size and sperm competition in mammals. Proc. Roy. Soc. London B., 243: 181-185.
CrossRef - Hammerstedt, R.H. and S.R. Hay, 1980. Effect of incubation temperature on motility and cAMP content of bovine sperm. Arch. Biochem. Biophys., 199: 427-437.
CrossRef - Heins, B.J., L.B. Hansen, A.J. Seykora, D.G. Johnson, J.G. Linn, J.E. Romano and A.R. Hazel, 2008. Crossbreds of Jersey x Holstein compared with pure holsteins for production, fertility and body and udder measurements during first lactation. J. Dairy Sci., 91: 1270-1278.
CrossRef - Hendricks, K.E.M. and P.J. Hansen, 2009. Can programmed cell death be induced in post-ejaculatory bull and stallion spermatozoa?. Theriogenology, 71: 1138-1146.
PubMed - Januskauskas, A., A. Johannisson and H. Rodriguez-Martinez, 2003. Subtle membrane changes in cryopreserved bull semen in relation with sperm viability, chromatin structure and field fertility. Theriogenology, 60: 743-758.
PubMed - Kasai, T., K. Ogawa, K. Mizuno, S. Nagai and Y. Uchida et al., 2002. Relationship between sperm mitochondrial membrane potential, sperm motility and fertility potential. Asian. J. Androl., 4: 97-103.
PubMed - Kjaestad, H., E. Ropstad and K.A. Berg, 1993. Evaluation of spermatological parameters used to predict the fertility of frozen bull semen. Acta Vet. Scand., 34: 299-303.
PubMed - Krzyzosiak, J., P. Molan and R. Vishwanath, 1999. Measurements of bovine sperm velocities under true anaerobic and aerobic conditions. Anim. Reproduct. Sci., 55: 163-173.
PubMed - Malo, A.F., M. Gomendio, J. Garde, B. Lang-Lenton, A.J. Soler and E.R.S. Roldan, 2006. Sperm design and sperm function. Biol. Lett., 2: 246-249.
PubMed - Miki, K., W. Qu, E.H. Goulding, W.D. Willis and D.O. Bunch et al., 2004. Glyceraldehyde 3-phosphate dehydrogenase-S, a sperm specific glycolytic enzyme is required for sperm motility and male fertility. Proc. Nat. Acad. Sci. USA., 101: 16501-16506.
CrossRef - Quill, T.A., S.A. Sugden, K.L. Rossi, L.K. Doolittle, R.E. Hammer and D.L. Garbers, 2003. Hyperactivated sperm motility driven by CatSper2 is required for fertilization. Proc. Nat. Acad. Sci., 100: 14869-14874.
CrossRef - Rikmenspoel, R., 1965. The tail movement of bull spermatozoa: Observations and model calculations. Biophys. J., 5: 365-392.
PubMed - Sarder, M.J.U., 2005. Study on dimensional characteristics of different breeds of bull spermatozoa. J. Anim. Vet. Adv., 4: 755-760.
CrossRefDirect Link - Shivaji, S., V. Kota and A. B. Siva, 2009. The role of mitochondrial proteins in sperm capacitation. J. Reproduct. Immunol., 83: 14-18.
PubMed - Suarez, S.S and H.C. Ho, 2003. Hyperactivated motility in sperm. Reproduct. Dom. Anim., 38: 119-124.
PubMed - Turner, R.G., 2003. Tales from the tail: What do we really know about sperm motility. J. Androl., 24: 790-803.
PubMed - Zhang, B.R., B. Larsson, N. Lundeheim and H. Rodriguez-Martinez, 1998. Sperm characteristics and zona pellucida binding in relation to field fertility of frozen-thawed semen from dairy Al bulls. Int. J. Androl., 21: 207-216.
CrossRef