
P. Anand
Department of Physiology, Rajah Muthiah Medical College,Annamalai University, Annamalainagar-608 002, Tamilnadu, India
D. Rajakumar
Department of Physiology, Rajah Muthiah Medical College,Annamalai University, Annamalainagar-608 002, Tamilnadu, India
A. John William Felix
Department of Community Medicine, Rajah Muthiah Medical College,Annamalai University, Annamalainagar-608 002, Tamilnadu, India
T. Balasubramanian
Department of Physiology, Rajah Muthiah Medical College,Annamalai University, Annamalainagar-608 002, Tamilnadu, India
Pakistan Journal of Biological Sciences
Year: 2010 | Volume: 13 | Issue: 16 | Page No.: 785-793







ABSTRACT
The present study evaluates the effects of oral administration of taurine on haematological parameters in normal wistar rats. Tissue oxidative stress is implicated in pathogenesis of various clinical disorders and antioxidant taurine is emerging as prophylactic and therapeutic agent. However, studies on effects of taurine on normal physiology are not reported in literature. Normal saline (Group I) or 5% taurine in normal saline was administered in dose of 50 mg (Group II), 250 mg (Group III) or 500 mg kg-1 of b.wt. (Group IV) through intragastric intubation for 60 days. The blood cell counts, haemoglobin content, packed cell volume, blood indices, bleeding time and clotting time were estimated using routine laboratory haematological techniques. Neutrophil’s phagocytic activity was determined by nitroblue tetrazolium reduction test; serum lysozyme activity was estimated colorimetrically by the degree to lyse bacterial cell suspension and serum taurine levels were estimated by HPLC fluorimetric technique. Platelet count showed a decrease in Group III and IV when compared with Group I and II (p<0.001). Mean corpuscular haemoglobin of Group III and IV are significantly lowered when compared to Group I (p<0.001). A statistically significant decrease was observed in the mean corpuscular haemoglobin concentration between Group I and Group IV (p<0.001). The neutrophil percentage of Group II, Group III and Group IV showed a significant increase over Group I (p<0.001). The percentage of lymphocytes showed a significant decrease in Group II, III and Group IV when compared to Group I (p<0.001). Neutrophil’s phagocytic activity is significantly lowered in Group III and IV when compared to Group I (p<0.001). The serum lysozyme activity of Group III and IV showed a significant increase over Group I (at p<0.001). From the present study it may be concluded that long term oral administration of taurine affects normal haematological functions.
PDF Abstract XML References Citation
Received: March 10, 2010;
Accepted: May 20, 2010;
Published: August 18, 2010
How to cite this article
P. Anand, D. Rajakumar, A. John William Felix and T. Balasubramanian, 2010. Effects of Oral Administration of Antioxidant Taurine on Haematological Parameters in Wistar Rats. Pakistan Journal of Biological Sciences, 13: 785-793.
DOI: 10.3923/pjbs.2010.785.793
URL: https://scialert.net/abstract/?doi=pjbs.2010.785.793
DOI: 10.3923/pjbs.2010.785.793
URL: https://scialert.net/abstract/?doi=pjbs.2010.785.793
INTRODUCTION
In tissues, the oxidation-reduction state also called the redox state depends on the delicate balance between oxidants and antioxidants status at any given time. The oxidants are the free radicals and reactive oxygen or nitrogen species (ROS and RNS, respectively) produced in the body as a result of regular intermediary metabolism and other oxidative processes like respiratory bursts seen in neutrophil during phagocytosis. Apart from their roles in the intermediary metabolism and body defence, the oxidants also play a major role as regulatory mediators in signalling processes. However, at high concentration, oxidants can damage all major cellular constituents and is termed as oxidative stress (Valko et al., 2007). The oxidative stress is implicated in the pathogenesis of various disorders like cancer, diabetes, cardiovascular diseases, autoimmune diseases and neurodegenerative disorders (Diplock, 1994; Mates et al., 1999; Gwarzo et al., 2010; Kayode et al., 2009; Vaghasiya and Chanda, 2010). Potentially damaging oxidative stress is kept in check by endogenous cellular antioxidant mechanisms, which include antioxidant enzymes like superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidise (GPX) and nutrient derived antioxidant small molecules (vitamin C, vitamin E, carotenes, flavonoids, glutathione, uric acid and taurine (Sardesai, 1995).
In recent years, antioxidants have gained a lot of importance because of their potential as prophylactic and therapeutic agents in many diseases. Several antioxidants like SOD, CAT, lycopene, co-enzyme Q10, vitamin C and vitamin E have been found to be pharmacologically active as prophylactic and therapeutic agents for various clinical disorders (Venkat Ratnam et al., 2006). Taurine (2-aminoethane sulfonic acid), a conditionally essential amino acid is a well documented antioxidant agent. Clinically taurine has been used with varying degrees of success in the treatment of a wide variety of conditions including cardiovascular diseases, hypercholesterolemia, epilepsy and other seizure disorders, macular degeneration, Alzheimer’s disease, hepatic disorders, alcoholism and cystic fibrosis (Birdsall, 1998). Taurine was reported to be beneficial in preventing experimental diabetic neuropathy (Obrosova et al., 2001), lead-induced oxidative damage (Gurer et al., 2001), CCl4-induced oxidative stress (Vohra and Hui, 2001), cerulein-induced acute pancreatitis (Ahn et al., 2001) and early changes in experimental diabetic kidney (Ha et al., 1999) through antioxidant mechanisms. Protective effects on lens (Devamanoharan et al., 1998) and modulating effects on human endothelial cell death are among its other effects (Wang et al., 1996).Taurine is one of the key ingredients in energy drinks, which are widely available in the grocery stores and convenience stores for easy access. Taurine is marketed as a dietary supplement for promotion of biliary health, eye health and prevention and treatment of congestive heart failure. Little is known regarding the effects of high-dose or long-term taurine use in children and adolescents (Babu et al., 2008).
Despite its extensive use, clinical and animal studies conducted with various doses of taurine on normal physiological functions are not reported in literature. In light of this context, the present study was designed to investigate the effects of oral administration of graded doses of taurine on haematological parameters in normal Wistar rats.
MATERIALS AND METHODS
Experimental animals: Albino Wistar rats (150-200 g; 20 weeks old ) of either sex were obtained from the Central Animal house, Rajah Muthiah Medical College, Annamalai University between December 2008 and February 2010. Animals were kept in polypropylene cages under standard laboratory conditions. The animals were maintained on standard rat chow and tap water ad libitum. The experiment was conducted in accordance with the accepted principles and guidelines of the Committee for the Purpose of Control and Supervision on Experiments on Animals (CPCSEA), India.
Chemicals and reagents: Taurine, homoserine, dowex resins, o-phthalaldehyde and mercaptoethanol were obtained from Sigma Chemicals Co.; St. Louis, MO. Methanol was purchased from Qualigens chemicals, India. Haemoglobin test kit was purchased from Diagnova, Ranbaxy India. All other chemicals used were of analytical grade.
Experimental design: Forty albino Wistar rats were divided into 4 groups of 10 rats each. Animals in Group 1 (Control) were administered 2.5 mL of 0.9% saline orally through intragastric intubation for 60 days. Animals in Group 2 (Low dose-Taurine), Group 3 (Medium dose-Taurine) and Group 4 (High dose-Taurine), were given 5% taurine in 0.9% saline orally at a dose level of 50, 250 and 500 mg kg-1 of b.wt., respectively through intragastric intubation for 60 days (Cetiner et al., 2005; Balasubramanian et al., 2004). At the end of 60 days blood samples were collected for analysis followed by determination of Bleeding Time (BT) and Clotting Time (CT).
Collection of blood samples: Following a fasting period of 12 h blood samples were withdrawn between 08:00 and 10:00 h from the retro-orbital sinus using fine capillary tube and collected into capped heparin coated tubes (1 mL) or capped plain tubes (0.5 mL). Blood samples collected in heparin coated tubes were used for haematological studies. Blood in plain tubes were allowed to clot and the clot to retract at room temperature, then centrifuged at 3000 rpm for 15 min at 4°C and the supernatant serum was collected and stored at -80°C for taurine estimation.
Bleeding time: Bleeding Time (BT) was assessed in rats restrained manually and amputating 5 mm of the tail tip with a scalpel blade and blood was blotted onto a filter paper every thirty seconds until the paper was no longer stained with blood. BT was measured in seconds from the time the tail amputated until bleeding ceased completely (Dejana et al., 1982).
Blood clotting time: Clotting time was determined using capillary glass tube method. Blood was drawn into capillary glass tube from retro orbital sinus and the time of collection was noted. Pieces of capillary glass were broken from one end at every thirty seconds and the appearance of fibrin threads was used as the end point and the time was noted in seconds (Harris et al., 1956).
Blood cell count: Red blood cell count (RBC count), Total Leukocyte Count (TLC) and platelet count were carried out manually using hemocytometer with phosphate buffered saline, turks fluid and 1% ammonium oxalate respectively and all the counts were expressed as number of cells per μL of blood (Lewis et al., 2006). The person who carried out the count was unaware of the experimental group of the animals.
Estimation of haemoglobin content: Haemoglobin content was estimated using haemoglobin test kit (Diagnova, Ranbaxy India) based on cyanmethemoglobin method and the Hb content was reported in g%.
Haematocrit [Packed cell volume (PCV)]: The microhaematocrit method was followed to determine the hematocrit. The plain capillary tube was filled with blood and the tube was sealed with a plastic seal and centrifuged at 12000 g for 5 min. After centrifugation, proportion of cells to the whole column was measured and the haematocrit value was expressed in percentage (Lewis et al., 2006).
Blood indices: Mean Corpuscular Volume (MCV), Mean Corpuscular Haemoglobin (MCH) and Mean Corpuscular Haemoglobin Concentration (MCHC) were calculated using the formulae described by Dacie and Lewis and expressed as μm3, pg and percentage, respectively (Lewis et al., 2006).
Differential count: Differential count was performed in blood smears stained with Leishman’s stain. The count was expressed in percentage for each type (Lewis et al., 2006). The person who did the count was unaware of the experimental group of the sample.
Estimation of serum taurine: Serum taurine levels were estimated by the method described by Waterfield (1994) using high-performance liquid chromatography with fluorimetric technique and the values are expressed as μM L-1.
Assay of serum lysozyme enzyme activity: The enzymatic activity of serum lysozyme against M. lysodeikticus cells was determined in 0.01 M sodium phosphate buffer, pH 6.2 at 37°C. The suspension was freshly prepared by dispersing 0.07 mg mL-1 M. lysodeikticus cells in phosphate buffer. Each assay was initiated by adding and mixing 0.1 mL hen egg white lysozyme (HEWL) solution or serum to 1 mL of bacterial cell suspension and 1 mL of sodium phosphate buffer. In the control sample 0.1 mL of deionised water was added instead of lysozyme solution. The solution was mixed and incubated at 37°C for 30 min and then, the test tubes were immediately placed on ice. The decrease in turbidity due to digestion of M.lysodeikticus by lysozyme was colorimetrically measured at 540 nm. Serum Lysozyme activity was expressed as μg mL-1 equivalent to HEWL activity (Ramanaviciene et al., 2002).
Assessment of neutrophil NADPH oxidase function (Nitroblue tetrazolium reduction test [NBT test]): A 0.5 mL heparinized blood samples were incubated at 37°C for 30 min on an acetone dried glass slide. The slides were then washed gently with cold saline and 0.6 mL NBT medium [(0.2 mL of 0.34% sucrose solution, 0.2 mL 0.28% Nitro Blue Tetrazolium (NBT) and 0.2 mL inactivated fetal calf serum (iFCS)] was added to each slide. After incubating for 30 min at 37°C, the slides were washed with cold saline and air-dried. Methanol was added to the smear (3 min) and washed with distilled water, air dried and stained with safranin. After staining, each slide was microscopically examined under oil immersion objective and 100 Neutrophils/slide were counted. The results were expressed in percentage of cells that reduced the NBT to dark blue formazan (NBT positive) (Gifford and Malawista, 1970).
Statistical analysis: One-way ANOVA statistical test was applied to find out any significant difference between the groups. If ANOVA results are significant Scheffe’s multiple comparison test was applied to find out which of the group differs. The level of significance was fixed at 5%.
RESULTS
Effects on bleeding time and clotting time: The bleeding time for Group I (control) is 81±14.5 sec, for group II (low dose-taurine) is 72±11.8 sec, for group III (Medium dose-taurine) is 82.5±7.9 sec and for Group IV (High dose-taurine) is 82.5±7.2 sec. The clotting time for Group I (control) is 112.4±28.2 sec, for group II (low dose-taurine) is 108±30.7 sec, for group III (Medium dose-taurine) is 114±62.6 sec and for Group IV (High dose-taurine) is 109.5±18.8 sec. There is no significant difference in bleeding time or clotting time between groups (Table 1).
Effects on blood cell counts: The red blood cell count for Group I (control) is 5.86±0.9 ×106 μL-1 of blood, for group II (low dose-taurine) is 6.35±0.8 ×106 μL-1 of blood, for group III (Medium dose-taurine) is 6.05±1.5 ×106 μL-1 of blood and for Group IV (High dose-taurine) is 6.4±0.5 ×106 μL-1 of blood. The total leukocyte count for Group I (control) is 5.82±2.07 ×103 μL-1 of blood, for group II (low dose-taurine) is 5.16±8.6 ×103 μL-1 of blood, for group III (Medium dose-taurine) is 5.5±6.2 ×103 μL-1 of blood and for Group IV (High dose-taurine) is 6.5±2.5×103 μL-1 of blood. The platelet count for Group I (control) is 5.62±0.9×105 μL-1 of blood, for group II (low dose-taurine) is 5.82±0.9×105 μL-1 of blood, for group III (Medium dose-taurine) is 4.56±0.4 ×105 μL-1 of blood and for Group IV (High dose-taurine) is 3.7±1.1 ×105 μL-1 of blood.
| Table 1: | Effects of taurine on hemostatic parameters |
 | |
| BT: Bleeding time; CT: Clotting time; Group I: Control, Group II: Low dose taurine; Group III: Medium dose taurine; Group IV: High dose taurine | |
| Table 2: | Effects of taurine on RBC count, Hb content, PCV and blood indices |
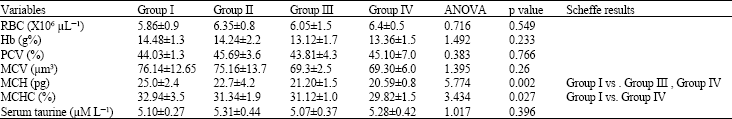 | |
RBC: Total RBC count; Hb: Hemoglobin content; PCV: Packed Cell Volume; MCV: Mean corpuscular volume; MCH: Mean corpuscular hemoglobin; MCHC: Mean corpuscular hemoglobin concentration; Group I: Control; Group II: Low dose taurine; Group III: Medium dose taurine; Group IV: High dose taurine. n = 10 for each group | |
| Table 3: | Effects of taurine on leukocytes count |
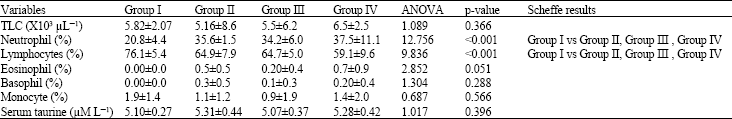 | |
| TLC: Total leukocyte count; Group I: Control; Group II: Low dose taurine; Group III: Medium dose taurine; Group IV: High dose taurine. n = 10 for each group | |
There is no significant difference in RBC count or total leukocyte count between groups. Platelet count is significantly lowered in the medium (Group III) and high (Group IV) dose taurine administered groups when compared to the control (Group I). The difference was statistically significant 5.62±0.9 ×105 μL-1 of blood vs 4.56±0.4 ×105 μL-1 of blood and 3.7±1.1 ×105 μL-1 of blood) at p<0.001 (Table 1).
Effects on haemoglobin content: The haemoglobin content for Group I (control) is 14.48±1.3 g%, for group II (low dose-taurine) is 14.24±2.2 g%, for group III (Medium dose-taurine) is 13.12±1.7 g% and for Group IV (High dose-taurine) is 13.36±1.5 g%. No significant difference was observed in the haemoglobin content between control and experimental groups (Table 2).
Effects on haematocrit [Packed Cell Volume (PCV)]: The Packed Cell Volume (PCV) for Group I (control) is 44.03±1.30%, for group II (low dose-taurine) is 45.69±3.6%, for group III (Medium dose-taurine) is 43.81±4.30% and for Group IV (High dose-taurine) is 45.10±7.00%. There is no significant difference in the PCV between groups (Table 2).
Effects on blood indices: The Mean Corpuscular Volume (MCV) for Group I (control) is 76.14±12.65 μm3, for group II (low dose-taurine) is 75.16±13.70 μm3, for group III (Medium dose-taurine) is 69.30±2.50 μm3 and for Group IV (High dose-taurine) is 69.30±6.0 μm3. The Mean Corpuscular Haemoglobin (MCH) for Group I (control) is 25.00±2.40 pg, for group II (low dose-taurine) is 22.70±4.20 pg, for group III (Medium dose-taurine) is 21.20±1.50 pg and for Group IV (High dose-taurine) is 20.59±0.80 pg. The mean corpuscular haemoglobin concentration (MCHC) for Group I (control) is 32.94±3.5%, for group II (low dose-taurine) is 31.34±1.9%, for group III (Medium dose-taurine) is 31.12±1.0% and for Group IV (High dose-taurine) is 29.82±1.5% (Table 2).
There is no significant difference in the MCV between groups. MCHs of medium (Group III) and high (Group IV) taurine administered groups are significantly lowered when compared to control rats (Group I) (25.0±2.4 pg vs. 21.20±1.5 pg, 20.59±0.8 pg) (Table 2). A statistically significant decrease was observed in the MCHCs between the control rats (Group I) and high taurine administered (Group IV) rats (32.94±3.5%, vs 29.82±1.5%) (Table 2).
Effects on differential leukocyte count (Table 3): The neutrophil percentage for Group I (control) is 20.80±4.40%, for group II (low dose-taurine) is 35.60±1.50%, for group III (Medium dose-taurine) is 34.20±6.00% and for Group IV (High dose-taurine) is 37.50±11.10%.
| Table 4: | Effects of taurine on neutrophil NAPDH oxidase function and lysozyme activity |
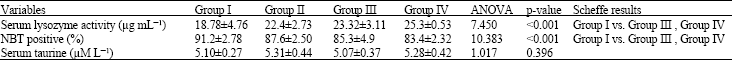 | |
| NBT positive: Nitroblue tetrazolium reduction test positive cells; Group I: Control; Group II: Low dose taurine; Group III: Medium dose taurine, Group IV: High dose taurine. n = 10 for each group | |
The eosinophil percentage for Group I (control) is 0.00±0.00%, for group II (low dose-taurine) is 0.50±0.50%, for group III (Medium dose-taurine) is 0.20±0.40% and for Group IV (High dose-taurine) is 0.70±0.90%. The basophil percentage for Group I (control) is 0.00±0.00%, for group II (low dose-taurine) is 0.30±0.50%, for group III (Medium dose-taurine) is 0.10±0.30% and for Group IV (High dose-taurine) is 0.20±0.40%. The monocyte percentage for Group I (control) is 1.90±1.40%, for group II (low dose-taurine) is 1.10±1.20%, for group III (Medium dose-taurine) is 0.90±1.90% and for Group IV (High dose-taurine) is 1.40±2.00%. The percentage of lymphocyte for Group I (control) is 76.10±5.40%, for group II (low dose-taurine) is 64.90±7.90%, for group III (Medium dose-taurine) is 64.70±5.00% and for Group IV (High dose-taurine) is 59.10±9.60%.
The eosinophil, basophil and monocyte percentage of the experimental groups did not differ significantly from control group. The neutrophil percentage of the taurine administered groups (Group II, Group III and Group IV) showed a significant increase over the control rats (Group I) (20.80±4.40% vs. 35.60±1.50%, 34.20±6.00% and 37.50±11.10) at p<0.001 (Table 3). The percentage of lymphocytes showed a significant decrease in the taurine administered groups (Group II, Group III and Group IV) when compared to the control group (Group I) (76.10±5.40% vs. 64.90±7.90%, 64.70±5.00% and 59.10±9.60%) at p<0.001 (Table 3).
Effects on neutrophil NADPH oxidase function (Nitroblue tetrazolium reduction test [NBT test]) (Table 4): The percentage of NBT positive cells for Group I (control) is 91.20±2.78%, for group II (low dose-taurine) is 87.60±2.50%, for group III (Medium dose-taurine) is 85.30±4.90% and for Group IV (High dose-taurine) is 83.40±2.32%. The percentage of NBT positive cells is significantly lowered in Group III (Medium dose-taurine) and in Group IV (High dose-taurine), when compared to the control group (91.20±2.78% vs. 85.30±4.90%, 83.40±2.32%) (Table 4). The difference was statistically significant at p<0.001.
Effects on serum lysozyme activity: The serum lysozyme activity for Group I (control) is 18.78±4.76 μg mL-1, for group II (low dose-taurine) is 22.40±2.73 μg mL-1, for group III (Medium dose-taurine) is 23.32±3.11 μg mL-1 and for Group IV (High dose-taurine) is 25.30±0.53 μg mL-1. The serum lysozyme activity of the medium (Group III) and high (Group IV) dose taurine administered group showed a significant increase over the control rats (Group I) (18.78±4.76 μg mL-1 vs. 23.32±3.11 μg mL-1, 25.30±0.53 μg mL-1) at p<0.001 (Table 4).
Effects on serum taurine levels (Table 1-4): Taurine concentration in the serum for Group I (control) is 5.10±0.27 μM L-1, for group II (low dose-taurine) is 5.31±0.44 μM L-1, for group III (Medium dose-taurine) is 5.07±0.37 μM L-1 and for Group IV (High dose-taurine) is 5.28±0.42 μM L-1. There is no significant difference in serum taurine levels between groups.
DISCUSSION
In many nations, including USA and UK, supplemental amino acids are available for non-prescription sales without any specific limits. Further, the occurrence of many amino acids including taurine in various dietary supplements and functional foods and beverages has increased dramatically in recent years (Shao and Hathcock, 2008). The increase in use and popularity of the antioxidant taurine as one of the ingredients in many dietary supplements necessitate the present study on the effects of oral administration of taurine on haematological parameters in otherwise normal rats. In the present study oral administration of taurine is not found to produce any significant change in the bleeding or clotting time. Studies on effects of oral administration of other antioxidants on bleeding or clotting time have not been reported in literature. However, the effects of administration of antioxidants on other haemostatic measures have been reported. Dereska et al. (2006) observed that moderate dose of oral dl-α-tocopherol acetate had no demonstrable effect on simulated bleeding time in the whole of blood of healthy individuals. In healthy male subjects, vitamin C supplementation was not associated with marked effects on haemostatic measures such as platelet adhesion and aggregation, levels of tissue plasminogen activator antigen, plasminogen activator inhibitor, fibrinogen, plasma viscosity and Von Willebrand factor (Tofler et al., 2000). Khaw and Woodhouse (1995) showed an inverse relationship between vitamin C levels and fibrinogen in humans aged 65 to 74 years. Bayele et al. (2002) demonstrated that redox state might have a role in controlling effector proteins of the coagulation system. He suggested that haemostasis might be influenced by dietary intake of antioxidants such as the carotenoids, flavonoids, tocopherols (vitamin E) and vitamin C (L-ascorbic acid). From available literature the effects of administration of antioxidants on bleeding time or clotting time can not be clearly defined. Further studies may throw more light on the effects of antioxidants including taurine on bleeding time or clotting time.
Recently, it has been shown by Fandrey et al. (1994) Huang et al. (1996) and Nagata et al. (2007) that erythropoietin (EPO) production and erythroid differentiation are regulated by ROS, especially by H2O2, which are involved in redox-sensitive signalling pathways through down-regulation of transcription factors. This means that ROS generation can suppress EPO synthesis whereas antioxidants can stimulate its synthesis. Attempts have been made by many researchers to evaluate the effects of antioxidants, such as β-carotene, desferrioxamine, tea polyphenols, α-tocopherol, ascorbic acid, reduced glutathione, N-acetylcysteine and α-lipoic acid on haematological parameters (RBC count, WBC count, MCH, MCHC and MCV) or EPO gene expression under normoxia or hypoxic conditions (Senturk et al., 2005; Freudenthaler et al., 2002; Hildebrandt et al., 2002; Niess et al., 2004; Jelkmann et al., 1997). Studies on N-acetyl cysteine and α-lipoic acid have shown a relationship between oxidative stress and erythropoiesis and suggested that only a few antioxidants could modulate haematological parameters (Zembron-Lacny et al., 2009).
Brown et al. (1994) reported a significant reduction in platelet count in non smokers as well as in smokers after vitamin E supplementation. An inverse relationship between platelet count and dietary vitamin E levels has been reported by Bendich et al. (1986). Hayes et al. (1989) reported that taurine stabilized platelets against aggregation such that during taurine depletion platelets became highly sensitive whereas during supplementation their tendency to aggregate was depressed. In the present study a decrease in platelet count was observed as the taurine dose was increased. This effect of taurine administration on platelet count is similar to results reported by others for other antioxidants.
Nakamura et al. (2001) in their study on oral toxicity of tocotrienol preparation in rats, have reported a significant decrease in platelet count, MCV and MCH in male rats and there was no change in RBC count, haemoglobin content and haematocrit values. In converse, study on protective role of vitamin E on diazinon (pesticide) toxicity, has shown that there was no significant change on haematological indices (RBC count, WBC count and Hb content, MCH, MCHC, MCV and thrombocyte count) between the control and vitamin E treated animals (Kalender et al., 2006). Oral administration of taurine is found to significantly produce a decrease in platelet count, MCH and MCHC and no change in the RBC count, Hb content or MCV.
In the present study oral administration of taurine significantly increased neutrophil count and decreased lymphocyte count without any significant change in total leukocyte count. The major function of taurine in leukocytes is to trap chlorinated oxidants (HOCl) and convert them into less toxic taurine chloramine (TauCl) and TauCl production is found to result in decreased NO production by Marcinkiewicz et al. (1998). Hence, taurine induced neutrophilia and lymphopenia without any change in total leukocyte count seen in our present study could be taurine’s action in converting HOCl to TauCl resulting in a decreased production of NO as supported by studies of Geffner et al. (1995) who have reported a decrease in NO production resulted in neutrophilia, lymphopenia without any change in total leukocyte count. However, the mechanism for this effect is not explained.
Studies on the effects of taurine or other antioxidants on serum lysozyme activity are not reported in literature. Lysozyme secreted by monocytes (Gordon et al., 1974) and present in lysosome of polymorphonuclear (PMN) leukocytes (Cohn, 1968) is a part of the non-specific innate immune system. In our present study, the serum lysozyme activity shows a significant rise in taurine administered groups. The mechanism for this rise in serum lysozyme activity can not be explained from the present study.
The NBT test is a simple method to evaluate neutrophil NADPH oxidase function that generates ROS during phagocyosis. The ability of neutrophils to reduce the almost colourless compound nitroblue tetrazolium (NBT) to dark blue formazan during phagocytosis reflects the production of intracellular superoxide anion. The significantly lower percentage of formazan positive neutrophils in taurine administered groups in our present study indicates decreased production of oxidative radicals by PMN and hence a decreased neutrophil phagocytic activity.
This could be secondary to an increased serum lysozyme activity found in our present study since lysozyme is found to significantly dampen several responses of neutrophils to inflammatory stimulants such as PMN chemotaxis, migration, oxidative metabolism and generation of ROS (Gordon et al., 1979). Further, the decrease in the production of intracellular oxidative radicals as represented by a decrease in formazan positive cells in our present study could be the result of a decrease in levels of pro-inflammatory cytokines reported for TNF-α and IL-6 by taurolidine (TRD) a synthetic derivative of taurine and taurine chloramine, a chlorinated form of taurine (Marcinkiewicz et al., 1998, 2006). Since Guerreiro et al. (2005) have reported that pro-inflammatory cytokines IFN-γ and TNF-α increase the formazan positive neutrophils , suggesting a positive correlation between IFN-γ and TNF-α concentration and neutrophil phagocytic activity.
Contrary to our in vivo findings, Farriol et al. (2002) using isolated neutrophils from healthy and burned patients have reported that addition of taurine to the samples increased neutrophil phagocytic activity as measured by nitroblue tetrazolium reduction test (NBT test). The difference between our in vivo finding and the in vitro observation reported by Farriol et al. (2002) could be due to the complex in vivo environments the neutrophils face in the body which are to be elucidated by more in vivo studies. The observed haematological changes following oral administration of taurine are seen without any significant changes in serum taurine levels. Sved et al. (2007) have pointed out that high concentration of taurine are maintained in blood cells, muscle and brain and specific Na+-dependent carriers in these cells and tissues are responsible for the high intracellular to extracellular ratios. It can be concluded that taurine exerts its effects at the cellular level and further studies are needed to establish the taurine’s mechanism of action for these observed haematological changes.
CONCLUSION
Oral administration of taurine to wistar rats in graded doses did not produce any change in serum taurine levels but produced a significant decrease in platelet and lymphocyte counts, MCH, MCHC and neutrophil phagocytic activity and a significant increase in neutrophil count and serum lysozyme activity. The study did not explore the underlying mechanisms for the observed haematological changes. However, the effects of large intake of taurine as dietary supplements and in fortified foods can be expected to affect the normal haematological functions and further studies are needed to establish the consequences of these physiological effects.
ACKNOWLEDGMENTS
I am grateful to Mrs. Radhika Balasubramanian for her help in preparing this manuscript.
REFERENCES
- Bendich, A., E. Gabriel and L.J. Machlin, 1986. Dietary vitamin E requirement for optimum immune responses in the rat. J. Nutr., 116: 675-681.
PubMed - Ahn, B.O., K.H. Kim, G. Lee, H.S. Lee and C.D. Kim et al., 2001. Effects of taurine on cerulein-induced acute pancreatitis in the rat. Pharmacology, 63: 1-7.
CrossRefDirect Link - Shao, A. and J.N. Hathcock, 2008. Risk assessment for the amino acids taurine, L-glutamine and L-arginine. Regulatory Toxicol. Pharmacol., 50: 376-399.
PubMed - Balasubramanian, T., M. Somasundaram and A.J.W. Felix, 2004. Taurine prevents ibuprofen-induced gastric mucosal lesions and influences endogenous antioxidant status of stomach in rats. Sci. World J., 4: 1046-1054.
PubMed - Waterfield, C.J., 1994. Determination of taurine in biological samples and isolated hepatocytes by high-performance liquid chromatography with fluorimetric detection. J. Chromatogr., 657: 37-45.
PubMed - Dejana, E., S. Villa and G. de Gaetano, 1982. Bleeding time in rats: A comparison of different experimental conditions. Thrombosis Haemostasis., 48: 108-111.
PubMedDirect Link - Devamanoharan, P.S., A.H. Ali and S.D. Varma, 1998. Oxidative stress to rat lens In vitro: Protection by taurine. Free Radic. Res., 29: 189-195.
PubMed - Jelkmann, W., H. Pagel, T. Hellwig and J. Fandrey, 1997. Effects of antioxidant vitamins on renal and hepatic erythropoietin production. Kidney Int., 51: 497-501.
PubMed - Fandrey, J., S. Frede and W. Jelkmann, 1994. Role of hydrogen peroxide in hypoxia�induced erythropoietin production. Biochem. J., 303: 507-510.
PubMed - Farriol, M., Y. Venereo, J. Rossello, P. Gomez and R. Palao et al., 2002. Effects of taurine on polymorphonuclear phagocytosis activity in burned patients. Amino Acids, 23: 441-445.
PubMed - Freudenthaler, S.M., K.H. Schreeb, A. Wiese, J. Pilz and C.H. Gleiter, 2002. Influence of controlled hypoxia and radical scavenging agents on erythropoietin and malondialdehyde concentrations in humans. Acta Physiol. Scand., 174: 231-235.
PubMed - Tofler, G.H., J.J. Stec, I. Stubbe, J. Beadle and D. Feng et al., 2000. The effect of vitamin C supplementation on coagulability and lipid levels in healthy male subjects. Thrombosis Res., 100: 35-41.
PubMed - Gifford, R.H. and S.E. Malawista, 1970. A simple rapid micromethod for detecting chronic granulomatous disease of childhood. J. Lab. Clin. Med., 5: 511-519.
PubMedDirect Link - Gordon, S., A. Todd and Z.A. Cohn, 1974. In vitro synthesis and secretion of lysozyme by mononuclear phagocytes. J. Exp. Med., 139: 1228-1248.
PubMed - Gurer, H., H. Ozgunes, E. Saygin and N. Ercal, 2001. Antioxidant effect of taurine against lead-induced oxidative stress. Arch. Environ. Contam. Toxicol., 41: 397-402.
CrossRefPubMedDirect Link - Nakamura, H., F. Furukawa, A. Nishikawa, M. Miyauchi and H.Y. Son et al., 2001. Oral toxicity of a tocotrienol preparation in rats. Food Chem. Toxicol., 39: 799-805.
PubMed - Ha, H., M.R. Yu and K.H. Kim, 1999. Melatonin and taurine reduce early glomerulopathy in diabetic rats. Free Radic. Biol. Med., 26: 944-950.
CrossRefPubMedDirect Link - Hayes, K.C., A. Pronczuk, A.E. Addesa and Z.F. Stephan, 1989. Taurine modulates platelet aggregation in cats and humans. Am. J. Clin. Nutr., 49: 1211-1216.
PubMed - Bayele, H.K., P.J. Murdock, D.J. Perry and K.J. Pasi, 2002. Simple shifts in redox/thiol balance that perturb blood coagulation. FEBS Lett., 510: 67-70.
CrossRef - Hildebrandt, W., S. Alexander, P. Bartsch and W. Droge, 2002. Effect of N-acetylcysteine on the hypoxic ventilatory response and erythropoietin production: Linkage between plasma thiol redox state and O2 chemosensitivity. Blood, 99: 1552-1555.
PubMed - Huang, L.E., Z. Arany, D.M. Livingston and H.F. Bunn, 1996. Activation of hypoxia-inducible transcription factor depends primarily upon redox-sensitive stabilization of its α-subunit. J. Biol. Chem., 271: 32253-32259.
PubMed - Guerreiro, J.B., M.A. Porto, S.B. Santos, L. Lacerda, J.L. Ho and E.M. Carvalho, 2005. Spontaneous neutrophil activation in HTLV-1 infected patients. Braz. J. Infect. Dis., 9: 510-514.
PubMed - Geffner, J.R., A.S. Trevani, I. de D`EIia, M. Diament, D. Klein and M. Giordano, 1995. Involvement of nitric oxide in the regulation of peripheral blood leukocyte counts. J. Leukocyte Biol., 58: 391-394.
Direct Link - Brown, K.M., P.C. Morrice and G.G. Duthie, 1994. Vitamin E supplementation suppresses indexes of lipid peroxidation and platelet counts in blood of smokers and nonsmokers but plasma lipoprotein concentrations remain unchanged. Am. J. Clin. Nuir., 60: 383-387.
Direct Link - Babu, K.M., R.J. Church and W. Lewander, 2008. Energy drinks: The new eye-opener for adolescents. Clin. Pediatr. Emerg. Med., 9: 35-42.
CrossRefDirect Link - Khaw, K.T. and P. Woodhouse, 1995. Interrelation of vitamin C, infection, haemostatic factors and cardiovascular disease. Br. Med. J., 310: 1559-1563.
PubMed - Gordon, L.I., S.D. Douglas, N.E. Kay, O. Yamada, E.F. Osserman and H.S. Jacob, 1979. Modulation of neutrophil function by Lysozyme. Potential negative feedback system of inflammation. J. Clin. Invest., 64: 226-232.
PubMed - Marcinkiewicz, J., A. Grabowska, J. Bereta, K. Bryniarski and B. Nowak, 1998. Taurine chloramine down-regulates the generation of murine neutrophil inflammatory mediators. Immunopharmacology, 40: 27-38.
PubMed - Marcinkiewicz, J., M. Kurnyta, R. Biedron, M. Bobek, E. Kontny and W. Maslinski, 2006. Anti-inflammatory effects of taurine derivatives (taurine chloramine, taurine bromamine and taurolidine) are mediated by different mechanisms. Adv. Exp. Med. Biol., 583: 481-492.
PubMed - Mates, J.M., C. Perez-Gomez and I.N. de Castro, 1999. Antioxidant enzymes and human diseases. Clin. Biochem., 32: 595-603.
CrossRefPubMedDirect Link - Cetiner, M., G. Sener, A.O. Sehirli, E. Eksioglu-Demiralp and F. Ercan et al., 2005. Taurine protects against methotrexate-induced toxicity and inhibits leukocyte death. Toxicol. Applied Pharmacol., 209: 39-50.
CrossRef - Nagata, M., N. Arimitsu, T. Ito and K. Sekimizu, 2007. Antioxidant N-acetylcysteine inhibits erythropoietin-induced differentiation of erythroid progenitors derived from mouse fetal liver. Cell Biol. Int., 31: 252-256.
CrossRef - Niess, A.M., E. Fehrenbach, I. Lorenz, A. Muller, H. Northoff and H.H. Dickhuth, 2004. Antioxidant intervention does not affect the response of plasma erythropoietin to short-term normobaric hypoxia in humans. J. Applied Physiol., 96: 1231-1235.
CrossRef - Dereska, N.H., E.C. McLemore, D.J. Tessier, D.S. Bash and C.M. Brophy, 2006. Short-term, moderate dosage vitamin E supplementation may have no effect on platelet Aggregation, coagulation profile, and bleeding time in healthy individuals. J. Surgical Res., 132: 121-129.
PubMed - Obrosova, I.G., L. Fathallah and M.J. Stevens, 2001. Taurine counteracts oxidative stress and nerve growth factor deficit in early experimental diabetic neuropathy. Exp. Neurol., 172: 211-219.
PubMed - Sardesai, V.M., 1995. Role of antioxidants in health maintenance. Nutr. Clin. Pract., 10: 19-25.
CrossRefPubMedDirect Link - Senturk, U.K., O. Yalcin, F. Gunduz, O. Kuru, H.J. Meiselman and O.K. Baskurt, 2005. Effect of antioxidant vitamin treatment on the time course of hematological and hemorheological alterations after an exhausting exercise episode in human subjects. J. Applied Physiol., 98: 1272-1279.
PubMed - Sved, D.W., J.L. Godsey, S.L. Ledyard, A.P. Mahoney and P.L. Stetson et al., 2007. Absorption, tissue distribution metabolism and elimination of taurine given orally to rats. Amino Acids, 32: 459-466.
CrossRef - Ratnam, D.V., D.D. Ankola, V. Bhardwaj, D.K. Sahana and M.N.V.R. Kumar, 2006. Role of antioxidants in prophylaxis and therapy: A pharmaceutical perspective. J. Controlled Release, 113: 189-207.
CrossRef - Vohra, B.P. and X. Hui, 2001. Taurine protects against carbon tetrachloride toxicity in the cultured neurons and in vivo. Arch. Physiol. Biochem., 109: 90-94.
CrossRefDirect Link - Wang, J.H., H.P. Redmond, R.W. Watson, C. Condron and D. Bouchier-Hayes, 1996. The beneficial effect of taurine on the prevention of human endothelial cell death. Shock, 6: 331-338.
PubMed - Kalender, Y., M. Uzunhisarcikli, A. Ogutcu, F. Acikgoz and S. Kalender, 2006. Effects of diazinon on pseudocholinesterase activity and haematological indices in rats: The protective role of vitamin E. Environ. Toxicol. Pharmacol., 22: 46-51.
CrossRefDirect Link - Zembron-Lacny, A., M. Slowinska-Lisowska, Z. Szygula, K. Witkowski and K. Szyszka, 2009. The comparison of antioxidant and haematological properties of N-acetylcysteine and alpha-lipoic acid in physically active males. Physiol. Res., 58: 855-861.
PubMed - Valko, M., D. Leibfritz, J. Moncol, M.T.D. Cronin, M. Mazur and J. Telser, 2007. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol., 39: 44-84.
CrossRefPubMedDirect Link - Gwarzo, M.Y., V.A. Nwachuku and A.O. Lateef, 2010. Prevention of alloxan induced diabetes mellitus in rats by vitamin a dietary supplementation. Asian J. Anim. Sci., 4: 190-196.
CrossRefDirect Link - Kayode, A.A.A., O.T. Kayode and A.A. Odetola, 2009. Anti-ulcerogenic activity of two extracts of Parquetina nigrescens and their effects on mucosal antioxidants defence system on ethanol-induced ulcer in rats. Res. J. Med. Plants, 3: 102-108.
CrossRefDirect Link