
Nwagu Tochukwu Nwamaka
Department of Microbiology and Biotechnology, University of Nigeria, Nsukka, Nigeria
Amadi Chike
Department of Microbiology and Biotechnology, Caritas University Amorji Nike, Enugu, Nigeria
Alaekwe Obiajulu
Department of Microbiology and Biotechnology, Caritas University Amorji Nike, Enugu, Nigeria
Pakistan Journal of Biological Sciences
Year: 2010 | Volume: 13 | Issue: 10 | Page No.: 497-503







ABSTRACT
Study was carried out to determine the spoilage association of traditionally fermented ugba, product of African oil bean seed. Samples were collected from three markets within Enugu metropolis in Eastern Nigeria. Microbial population of ugba was isolated, characterized and identified. Effect of time on product quality, total viable cell count and individual growth pattern of isolates was evaluated. Microorganisms isolated were Proteus sp., Klebsiella sp., Staphylococcus epidermidis, E. coli, Bacillus licheniformis and Bacillus subtilis. Initial viable cell count increased from 4.6x108-6.2x108, 6.0x109-6.9x109 and 3.9x108-5x108 cfu g-1 by the 4th day for samples from Mayor, Kenyatta and Garki markets, respectively. Organoleptic changes in texture and colour were seen to be a factor of time of storage. A decline in growth of Proteus sp., E. coli, Klebsiella sp. and S. epidermidis by the 4th day of storage was observed for all samples. Viable cells of B. licheniformis increased from 6.1x107-17.7x107 cfu g-1, 13.9x108-20.0x108 and 8.7x107-15.5x107 by the 4th day of storage for samples from Mayor, Kenyatta and Garki markets respectively. Growth of B. subtilis increased from 9.2x107-19.9x107, 14.9x108-21.2x108 and 11.5x107-17.2x107 cfu g-1 for samples from Mayor, Kenyatta and Garki markets respectively. Our results indicate that ugba spoilage is primarily a result of the continued activity of African oil bean seed fermentative organisms B. subtilis and B. licheniformis.
PDF Abstract XML References Citation
Received: February 27, 2010;
Accepted: May 20, 2010;
Published: June 23, 2010
How to cite this article
Nwagu Tochukwu Nwamaka, Amadi Chike and Alaekwe Obiajulu, 2010. Role of Bacteria Isolates in the Spoilage of Fermented African Oil Bean Seed Ugba. Pakistan Journal of Biological Sciences, 13: 497-503.
DOI: 10.3923/pjbs.2010.497.503
URL: https://scialert.net/abstract/?doi=pjbs.2010.497.503
DOI: 10.3923/pjbs.2010.497.503
URL: https://scialert.net/abstract/?doi=pjbs.2010.497.503
INTRODUCTION
Ugba is a fermented product from African oil bean seed Pentaclethra macrophylla Benth, a woody plant predominant in the rain forest areas of West and Central Africa belonging to the family Leguminosaea, sub-family Mimosoidae (Keay, 1989). Ugba is of primary importance as cheap and easily available source of plant protein in developing countries of the world and Africa in particular. Fermented seeds are not just palatable but serve as a delicacy amongst consuming regions where it is consumed garnished with other vegetables or staples. Consumption of ugba seeds could pose as a means of addressing the prevailing Protein Energy Malnutrition (PEM) in developing countries (Enujiugha and Akanbi, 2008).
African oil bean seeds are oval, flat and black in colour. The seeds are composed of 35-52% oil, 17-22% protein and 12-23% carbohydrates (Oboh and Ekperigin, 2004). Unprocessed seeds are bitter and possess anti-nutritional factors amongst which are pancine, cyanide, oxalates, saponin, phytic acid, phytate and tannins (Enujiugha and Akanbi, 2005). Processing of these seeds entails boiling, removal from pod, cutting into slices, further boiling, wrapping in banana/ plantain leaves and fermentation. Thermal treatment induces a resultant rise in nutrient bioavailability and seed digestibility. Processing ugba seeds drastically reduces the levels of the anti nutritional compounds mentioned while increasing iron, calcium, potassium, thiamine and riboflavin levels (Enujiugha and Ayodele-Oni, 2003).
Preparation of ugba is by mixed fermentation carried out spontaneously by a number of microorganisms. Microorganisms isolated from fermented ugba include Micrococcus sp., Lactobacilli, Staphyloccus sp., Leuconostoc mesenteroides, Proteus and E. coli (Isu and Njoku, 1997; Mbata and Orji, 2008; Obeta, 1983). The major problem with the fermented oil bean seed ugbais the restricted availability due to its very short shelf life. Under room temperature, fermented ugba spoils within three to four days. Spoilage is identified with increased softness (Enujiugha et al., 2008), color change, off flavor and sliminess (Mbata and Orji, 2008) and production of pungent ammonical odor (Ogbulie et al., 1993).
Extending the shelf life of ugba has been a case of interest to many researchers. Reports on strategies to extend the shelf life of fermented ugba include treatment with varying concentrations of sodium chlorid (Ogbulie et al., l993), preservation in high density polyethylene sachets and aluminum foil wraps (Ogbulie et al., 1998), canning within tomato puree, brine solution or refined groundnut oil (Enujiugha and Akanbi, 2005) and use of starter cultures to shorten period of fermentation (Mbata and Orji, 2008). The deterrent in the methods reported so far include the cost of implementation, no remarkable difference in shelf life extension and lack of ease in method application making it difficult or impossible for local commercial dealers to implement such methods.
A key selling point to using fermented oil bean seed ugba to address Protein Energy Malnutrition (PEM) issues is the ease of process adoption by local producers. Advocating for commercialization of this process though commended may inadvertently lead to a hike in product prize making this rich source of protein unavailable to the impoverished that are direly in need of the nutrients, which this can provide. A concise knowledge of the spoilage associations of fermented oil bean seed will enable the identification of the chief sources of spoilage and therefore help to identify simpler techniques to address spoilage problems.
This study is aimed to isolate spoilage associations of fresh ugba collected from different areas and determine succession or increase in microbial population as related to deterioration of product.
MATERIALS AND METHODS
Sampling: Fermented ugba was purchased from local dealers from different markets in Enugu State, Nigeria. A total of 12 samples were collected in 2009, three each from Garki, Mayor and Kenyatta markets in the Enugu metropolis of Eastern Nigeria. Samples were freshly fermented (3 days) according to dealers to ensure uniformity of results and unbiased random sampling.
Isolation and enumeration of microorganisms: Fermenting samples were taken aseptically (using sterile forceps) from the fermenting beans at 24 h intervals for five days. A 10 g quantity of each sample was homogenized in a tared blender with sterilised cups using 90 mL of sterile 0.1% peptone water as diluent Subsequent decimal dilutions (10-2, 10-3, 10-4, 10-5 etc.) were made from this homogenate by adding serially 10 mL of solution from proceeding concentration of 90 mL of the diluent. On to a Petri dish containing 0.1 mL of the inoculum 10-15 mL of sterile medium (Nutrient Agar NA), blood agar and Cystine Lactose Electrolyte Deficient Agar (CLED) was poured for the isolation of bacteria while 1 mL of the inoculum was used for the isolation of yeasts and fungi. The plates were gently rotated to distribute the inoculum evenly in the plate and left to solidify under a laminar airflow. Half of the plates for each medium were incubated aerobically at 30°C for 24-48 h while the other half were incubated anaerobically at 30°C for 72 h using a Gas Pak anaerobic system. Anaerobiosis was confirmed with a BBL anaerobic indicator. Incubation of the mould and yeasts was at 25°C for 5 days. Nutrient agar (DIFCO) and plate count agar (DIFCO) were the isolation and enumeration media for the bacteria. Potato Dextrose Agar (OXOID) and Malt Extract Agar (OXOID) were used for the isolation of the fungi and yeast (Cheesbrough, 2000). At the end of the various incubation periods, only plates showing between 30 and 300 colonies were counted using the digital illuminated colony counter (Gallenkamp). Colony counts were expressed as colony forming units per gram of sample. All counts were done in triplicate and average values were reported.
Characterization and identification of isolates: Representative colonies of isolates streaked were on nutrient agar medium. Pure cultures resulting from the isolation were subcultured and preserved on nutrient agar slants at 4°C. Cultural characteristics of the isolates on the plates were noted. The motility of the isolates was examined by the hanging drop technique. Gram reactions and cell morphology were examined from heat-fixed smears. The microorganisms were identified by the methods described by Gordon et al. (1973), Cowan and Steel (1974) and Bergeys Manual of Determinative Bacteriology (Buchanan and Gibbons, 1974).
Monitoring proliferation of isolates: Sampling was carried out at 24 h intervals from mixed samples taken from the surfaces and central parts of the wraps from Garki, Mayor and Kenyatta market. Samples (1 g each) were placed in sterile 100 mL Erlenmeyer flask under water; 10 mL distilled water and thoroughly mixed by shaking. Mixture was serially diluted, plated out and counted using a digital illuminated colony counter as earlier stated. Colony counts were expressed as colony forming units per gram of isolates.
RESULTS
Isolation of microorganisms: A total of six bacteria genera were isolated from various samples collected from three different markets (Table 1). Further characterization revealed these organisms to be Bacillus subtilis and Bacillus licheniformis, Escherichia coli, Staphylococcus epidermidis, Micrococcus roseus, Proteus sp. and Klebsiella aerogenes (Table 2).
| Table 1: | Morphological features and characterization of bacterial isolates from Ugba |
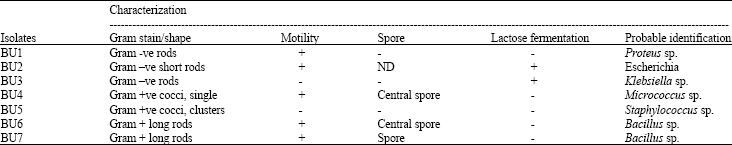 | |
| +: Present; -: Absent | |
| Table 2: | Biochemical reaction patterns in primary tests for identification of isolated microorganisms from ugba |
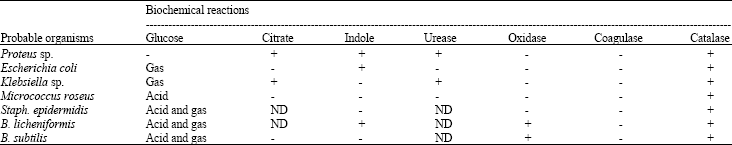 | |
| ND: Not detected, +: Present, -: Absent | |
Proteus sp., grew on blood agar with characteristic swarming over the surface of blood agar and distinctive fishy smell. On Cystine-Lactose Deficient Agar (CLED), Proteus sp was non-lactose fermenting.
E. coli yielded 1-4 mm diameter mixed colonies, which were partially hemolytic on blood agar. On CLED, lactose fermenting mucoid colonies was seen.
Klebsiella sp., colonies were seen as large mucoid non-hemolytic colonies on blood agar. On CLED, large lactose fermenting mucoid colonies, which exhibited a drawing characteristic while being picked with the wire loop, was observed.
Micrococcus roseus produced hemolytic smooth convex regular edged colonies in blood agar while growth on CLED showed non-lactose fermenting colonies.
Staphylococcus epidermidis appeared as whitish yellowish to creamy, slightly raised 1-2 mm in diameter colonies on blood agar. Growth in CLED yielded smaller (0.1-0.5 mm) colonies, seen as cocci which appeared singly or in clusters.
Bacillus licheniformis were seen as light green raised colonies with rough surfaces on blood agar. Smaller (<0.1-0.5 mm) non-lactose fermenting colonies were seen on CLED.
Bacillus subtilis grew in form of dry, opaque circular raised surface, about 0.1-0.5 mm in diameter in blood agar. On CLED, colonies were smaller and lactose fermenting.
Effect of time on the quality and microbial population of ugba: Initial microbial load of samples from Mayor market was 4.6×108 cfu g-1, by the fourth day, bacterial population had increased to 6.2×108 cfu g-1.
| Table 3: | Effect of period of storage on quality and microbial population ugba from mayor market |
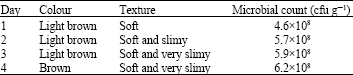 | |
| Table 4: | Effect of period of storage on quality and microbial population ugba Kenyatta market |
 | |
Product turned from brown to a darker shade of brown and was very soft and slimy (Table 3). Samples from Kenyatta market had the largest number of bacterial contaminants with 6.0×109 cfu g-1 on the first day (Table 4) while samples from Mayor market had 4.6×108 cfu g-1 and samples from Garki market had the lowest number of 3.9×108 cfu g-1 on the first day (Table 5). Samples from Kenyatta market experienced rapid organoleptic changes: colour turned from light brown to brown and then to dark brown with ugba been very slimy and soft by the fourth day (Table 4). Organoleptic changes of finished product progressed slowly for samples from Garki market, product turned from light brown to brown by the fourth day, corresponding with microbial proliferation which increased from 3.9×108 to 5×108 (Table 5).
Daily growth pattern of individual bacteria isolates: For samples from Mayor market, growth of Proteus sp., was 3.1×107cfu g-1 on the first day, there was no change in viable cell count on the 3rd and 4th day, growth was stationary with same value of 6.3×107 cfu g-1 (Fig. 1).
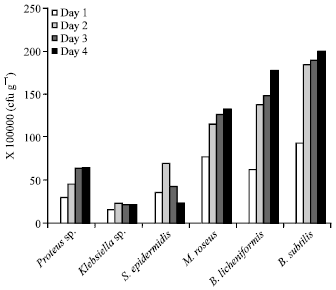 | |
| Fig. 1: | Growth of microorganisms in fermented ugba from Mayor market over a period of 4 days. Viable cells counts were in cfu g-1 |
| Table 5: | Effect of period of storage on quality and microbial population ugba from Garki market |
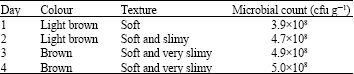 | |
Initially viable cell count of Klebsiella sp was 1.5×107, a decline in growth from 2.3×107 cfu g-1 to 2.1×107 on the 3rd day and 2.0×107 cfu g-1 on the 4th day was observed. Similar growth pattern was seen in the case of S. epidermidis with viable cell counts increasing from 3.5×107 cfu g-1 on 1st day to 6.8x107 and 4.2×107 on the 2nd and 3rd day, followed by a decline to 2.2x107 on the 4th day. Growth increased from 7.6×107-13.2×107, 6.1×107 -17.7×107 and 9.2×107-19.9×107 for M. roseus, B. licheniformis and B. subtilis respectively after storage for 4 days as shown in Fig. 1.
A decline in the growth of E. coli from 5.5×108 on the first day to 2.1×108 on the fourth day was seen in samples from Kenyatta market (Fig. 2). Growth of K. aerogenes reduced from 5×108 on the first day to 2×108 on the fourth day, while that of S. epidermidis reduced from 7.3×108 to 4.2×108 cfu g-1. Growth increased from 9.1×108-11.5×108, 13.9×107-20.0×107 and 14.9×108-21.2×108 cfu g-1 for M. roseus, B. licheniformis and B. subtilis respectively after storage for 4 days as shown in Fig. 2.
Growth of Proteus sp in samples from Garki market increased from 5.78×107 to 7.34×107 on the 2nd day, followed by a decline to 6.13×107 and 3.45×107 by the 3rd and 4th day, respectively (Fig. 3).
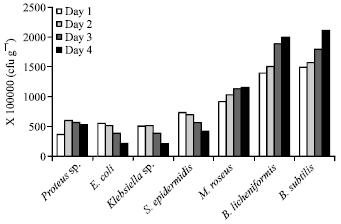 | |
| Fig. 2: | Growth of microorganisms present in fermented ugba from Kenyatta market over a period of 4 days. Viable cells counts were in cfu g-1 |
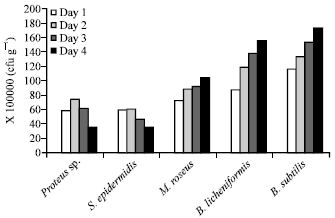 | |
| Fig. 3: | Growth of microorganisms present in fermented ugba from Garki market over a period of 4 days. Viable cells counts were in cfu g-1 |
S. epidermidis increased from 5.78×107-5.90×107 cfu g-1 on the 2nd day followed by a gradual decline to 3.45×107 by the 4th day. Growth increased from 7.2×107-10.3×107, 8.7×107 -15.5×107 and 11.5×107-17.2×107 for M. roseus, B. licheniformis and B. subtilis, respectively after storage for 4 days as shown in Fig. 3.
DISCUSSION
Microorganisms isolated from fermented African oil bean seed ugba corresponded with organisms previously isolated by other workers (Obeta, 1983; Isu and Njoku, 1997). Only bacteria were isolated, no molds and yeast were involved, which correlated with earlier reports (Njoku et al., 1990). Mbajunwa et al. (1998) reported the isolation of Bacillus cereus, B. subtilis, Pseudomonas chlororaphis, Micrococcus roseus and Staphylococcus saprophyticus. Though researchers have attempted using starter cultures for fermentation of African oil bean seed, ugba production is still by spontaneous inoculation from water used for soaking and steeping (Njoku et al., 1990), from the air and from leaves used for wrapping the ugba (Mbajunwa et al., 1998). The random and indefinite source of inoculants for ugba production partly explains the wide range of microorganisms isolated. From our work, it is evident that microorganisms present were introduced during processing; there was no change or evidence of microbial succession as keeping time of ugba increased. The change witnessed was the increase in number of existing microorganisms, which probably led to change of product colour and texture. To our knowledge no similar work had been carried out on ugba, so there was no way to compare our results with that of other investigators. The variation in types of microorganism present and microbial load of each specie leads to non-uniform products with short shelf lives. Freshly fermented ugba was light brown in colour and soft. There was a darkening of colour to brown and dark brown and change in texture, with product becoming extremely soft as fermentation continued unstopped in the finished product. Microbial content was seen to increase with increase in time, indicating microbial proliferation as a function of time. The increase observed in microbial load possibly caused a resultant increase in cellular activities leading to unsolicited organoleptic change in product quality. This is in agreement with the report by Mbata and Orji (2008) that spontaneous activities of microorganisms present lead to short shelf -life of product. Colour change is thought to be as a result of the presence of polyphenol oxidase (a major enzyme contained in oil bean seed) that catalyzes the oxidation of phenolic substances to quinine, which is polymerized to form a brown pigment (Enujiugha and Akanbi, 2005). Colour change in ugba has also been attributed to the activities of microorganisms present in ugba. Presence of Pseudomonas chlororaphis during ugba fermentation resulted to the production of a greenish colored ugba due to the presence of chlororaphin, a green phenazine pigment (Mbajunwa et al., 1998). Fermented ugba turned softer with time due to the presence of certain bacteria species. Softness during fermentation was attributed to Bacillus species especially Bacillus subtilis (Obeta, 1983; Mbata and Orji, 2008). Though activity of Pseudomonas chlororaphis was also responsible for ugba softening (Mbajunwa et al., 1998), this organism was not isolated from our work. Enujiugha and Akanbi (2008) isolated and used Pseudomonas fluorescence as starter culture for ugba production, but they reported that its activity during production was insignificant. It is also worthy to note that Bacillus sp. have been implicated in all the reported cases of ugba fermentation, either when fermented spontaneously or inoculated specifically as starter cultures (Enujiugha, 2003; Mbajunwa et al., 1998; Mbata and Orji, 2008; Njoku et al., 1990; Obeta, 1983). Moreover, Sanni et al. (2000) reported that amongst a total of 97 strains of spore forming Bacillus isolated from 45 samples of three Nigerian fermented soup condiments (iru, ugba and ogiri) obtained from different markets, 33% were B. subtilis; 22% were B. licheniformis; 19% B. pumulus; 12% B. megaterium; 9% B. brevis and 5%, B. polymyxa. This is in agreement with our observation of softening of product as growth of Bacillus subtilis and B. licheniformis increased and corresponds with the report by Obeta (1983) that B. subtilis was responsible for softening of ugba during fermentation. Deterioration of ugba quality was also witnessed in form of sliminess, a common occurrence in spoilage of food with high residual moisture content. This is in agreement with the report of Mbajunwa et al. (1998) that ugba fermentation by B. subtilis was accompanied by extensive production of whitish mucilaginous strands (gums) which caused the sticking together of ugba strands. According to Sanni et al. (2000) the residual moisture content of ugba was 46.32%. Slime is formed by a variety of Extracellular Polymeric Substances (EPS) that are long thread like molecules, which often appear like gel-like growth on the food. These extracellular polymers, EPS coat the bacteria cells into a common slime mass, which become clustered and bound within large volumes (90-95%) of water. Presence of these gums is probably responsible for ensuring that spores of B. subtilis and B. licheniformis dominate the environment. Though many bacteria including members of the Enterobacteriaceae produce slime; slime layers in Bacilli spps, appear in form of capsules tightly bound to individual cells protecting each cell from adverse conditions. Slime is formed by bacteria under stress and layers of slime increase with duration of adverse conditions especially under aerobic (oxidative conditions) though slime bacteria also function under oxidation-reduction conditions. This may explain the ability of the Bacillus sp to withstand the high alkaline environment obtainable during ugba production. The continual increase in population of B. subtilis and B. licheniformis is an indication that our reports correspond with that of previous workers that these organisms play active role in ugba (Obeta, 1983; Mbajunwa et al., 1998; Enujiugha et al., 2008). However, our work further confirms that continuous activities of these organisms are actually responsible for the spoilage of ugba. To the best of our knowledge, no research has addressed the problem of ugba spoilage from this angle. Many have tried to identify the role of organisms during ugba production rather than their post production roles (Mbajunwa et al., 1998, Enujiugha et al., 2008). In our research, we worked with already produced ugba, monitoring activities of microorganisms present. Experiment showed that amongst all samples from various markets, colony forming units of E. coli, Klebsiella aerogenes, Staphylococcus epidermidis and Proteus sp., reduced drastically with increase in keeping time. This is an indication that these organisms were introduced into the food as microbial contaminants, which play no role in actual fermentation of ugba and could not tolerate the environment as spontaneous fermentation progressed unabated. The boiling stages involved in the preparation of African oil bean seed prior to the fermentation would eliminate most natural microbial flora of the seeds. However, Bacillus species possess heat resistant spores and therefore are able to resist such high temperatures. Sources of contamination could be air, water, equipments used during processing such as utensils, leaves used for wrapping or even through the human handler. Staphylococcus epidermidis is a member of the normal flora of the human skin, respiratory and gastrointestinal tracts, which could easily have been introduced into the food by the human handler. E. coli, Klebsiella sp. and Proteus sp are enteric bacteria, whose natural habitat is the intestinal tract of humans and animals (Jawetz et al., 1989). Enteric bacteria are also regarded as indicators of feacal contamination of water and could be introduced in food through water or poor hygiene during the fermented food production. This correlates with the work of Mbata and Orji (2008), who reported a controlled fermentation of African oil bean seed using Bacillus subtilis and B. megaterium isolated from traditionally produced ugba as starter cultures. The product was reported to meet the standard stipulated quality of ugba including correct colour, aroma, texture and taste. This indicates that use of these Bacillus sp. could successfully lead to good and proper ugba quality. The population of B. licheniformis, B. subtilis and Micrococcus roseus increased with keeping time for all samples from various markets. Micrococcus is usually isolated from air, water and salt containing food (Buchanan and Gibbons, 1974). Mbajunwa et al. (1998) reported that Micrococcus sp. does not play any significant role in ugba production while Bacillus sp. are known to play key roles. Fermented ugba seeds contain a high percentage of oil, protein and relatively small concentration of carbohydrate. Bacillus sp. and Micrococcus sp. are known producers of lipolytic enzymes and therefore possess the relevant machinery for efficient breakdown of oil and other substrates, to access their nutrients. The ability of these organisms to perform these functions will ensure their persistence in the environment and explain the increase in number with increase in keeping time. The proteolytic activities lead to an increase in ammonia nitrogen (Odunfa, 1981) and other amino acids with a resultant increase in pH (Mbajunwa et al., 1998). Since only a few bacteria are favored by environmental pH above neutral, the increase in pH to high alkalinity makes the environment hostile for other microorganisms while favoring that of Bacilli sp. This is in agreement with the report by Odunfa and Oyeyiola (1985). Though Odunfa and Oyeyiola (1985) reported that increase in pH encouraged growth of Staphylococcus species and Bacillus species, it is probable that there were other existing factors responsible for the reduction of Staphylococcus population in our samples as product deterioration progressed. The increase in population of Bacillus sp. shows that these organisms are the key spoilage organisms of ugba. Spoilage is most likely a result of unstopped fermentation by Bacillus sp., which are not killed or inactivated after ugba production.
CONCLUSION
From the results of our experiments, Bacillus sp. were observed to be the key spoilage organisms of ugba. Spoilage was noted to be due to continuous activities of Bacillus sp. after desired fermentation was reached. Other organisms isolated from ugba are likely contaminants exposed to the food after boiling, prior to fermentation through the air, water, leaves or poor aseptic measures during handling. To ensure a simple method of maintaining ugba quality with a longer shelf life, good hygienic practices, sterile water and clean environment should be maintained. Additionally, experiments should be targeted towards simple, uncomplicated methods for the removal, killing or inactivation of Bacillus sp. or their enzymes present in the finished products to maintain ugba quality for a longer period of time.
REFERENCES
- Enujiugha, V.N. and C.T. Akanbi, 2005. Compositional changes in African oil bean (Pentaclethra macrophylla Benth) seeds during thermal processing. Pak. J. Nutr., 4: 27-31.
CrossRefDirect Link - Enujiugha, V.N. and C.T. Akanbi, 2008. Quality evaluation of canned fermented oil bean seed slices during ambient storage. Afr. J. Food Sci., 2: 54-59.
Direct Link - Enujiugha, V.N., C.T. Akanbi and H.A. Adeniran, 2008. Evaluation of starters for the fermentation of African oil bean (Pentaclethra Macrophylla Benth) seeds. Nutr. Food Sci., 38: 451-457.
CrossRef - Enujiugha, V.N. and O. Ayodele-Oni, 2003. Evaluation of nutrients and some anti-nutrients in lesser-known, underutilized oilseeds. Int. J. Food Sci. Technol., 38: 525-528.
CrossRefDirect Link - Isu, N.R. and H.O. Njoku, 1997. An evaluation of the microflora associated with fermented African oil Bean (Pentaclethra macrophylla) seeds during ugba production. Plant Foods Hum. Nutr., 51: 145-157.
Direct Link - Mbajunwa, O.K., J.O. Akingbola, K. Mulongoy and G. Oguntimein, 1998. Starter culture evaluation from the production of ugba from African oil bean seed Pentaclethra macrophylla. J. Sci. Food Agric., 77: 127-132.
Direct Link - Mbata, T. and M.U. Orji, 2008. Process optimization in the production and preservation of Ugba, a Nigerian fermented food. Int. J. Microbiol., 4: 2-6.
Direct Link - Njoku, H.O., J.N. Ogbulie and C. Nnubia, 1990. Microbiological study of the traditional processing of African oil bean (Pentaclethra macrophylla) for ugba production. Food Microbiol., 7: 13-26.
CrossRef - Oboh, G. and M.M. Ekperigin, 2004. Nutritional evaluation of some Nigerian wild seeds. Nahrung Food, 48: 85-87.
CrossRefDirect Link - Odunfa, S.A., 1981. Microorganisms associated with the fermentation of African locust bean (Parkia filicoidea) during iru preparation. J. Plant Food, 25: 245-250.
CrossRef - Sanni, A.I., G.S. Ayernor, E. Sakyi-Dawson and S. Sefa-Dedeh, 2000. Aerobic spore-forming bacteria and chemical composition of some Nigerian fermented soup condiments. Plant Foods Human Nutr., 55: 111-118.
CrossRef