
M. H. Abdally
Department of Microbiology and Parasitology, College of Veterinary Medicine and Animal Resources, King Faisal University, P.O. Box 1757, Al-Ahsa 31982, Al-Ahsa, Kingdom of Saudi Arabia
M. M. Abo-Elsaad
Date Palm Research Center (Center of Excellence), King Faisal University, Al-Ahsa, Kingdom of Saudi Arabia
A. A. Al-Shaggag
Al-Hasa National Date Palm Research Center, Ministry of Agriculture, Al-Ahsa, Kingdom of Saudi Arabia
M. M. Al-Bagshy
Al-Hasa National Date Palm Research Center, Ministry of Agriculture, Al-Ahsa, Kingdom of Saudi Arabia
A. A. Al-Shawaf
Al-Hasa National Date Palm Research Center, Ministry of Agriculture, Al-Ahsa, Kingdom of Saudi Arabia
Pakistan Journal of Biological Sciences
Year: 2010 | Volume: 13 | Issue: 5 | Page No.: 223-228







ABSTRACT
The hemagglutination activities of midgut (MG) fractions of red palm weevil, Rhynchophorus ferrugineus against mammalian erythrocytes (RBC) from man (ABO), rabbit, horse and sheep were studied. The highest titers were seen with rabbit RBC (p<0.05) followed by human group B, human group O, horse, human group A, human group AB and sheep, respectively. Insect age and nutritional status significantly influenced the agglutinating activities. Some of the unfed insects were having low activities (p>0.01) than the well fed insects (p<0.05). The agglutination activities were linearly correlated with the insect age. The agglutination activities were higher in old insects than the young insects (p<0.05). Further studies are needed to investigate the lectins in the hemolymph of the red palm weevil, Rhynchophorus ferrugineus beside demonstrating their agglutination activities against the natural entomopathogenes organisms of this economically important insect for use in its biological control.
PDF Abstract XML References Citation
How to cite this article
M. H. Abdally, M. M. Abo-Elsaad, A. A. Al-Shaggag, M. M. Al-Bagshy and A. A. Al-Shawaf, 2010. Detection of Insect Immunity Substances (Lectins) in the Midgut Extracts from Larvae and Adult Red Palm Weevil Rhynchophorus ferrugineus (Olivier) in Al-Ahsa, Saudi Arabia. Pakistan Journal of Biological Sciences, 13: 223-228.
DOI: 10.3923/pjbs.2010.223.228
URL: https://scialert.net/abstract/?doi=pjbs.2010.223.228
DOI: 10.3923/pjbs.2010.223.228
URL: https://scialert.net/abstract/?doi=pjbs.2010.223.228
INTRODUCTION
The Red Palm Weevil (RPW), Rhynchophorus ferrugineus (Curculionidae: Coleoptera) is an economically important tissue boring pest of date palm in many parts of the world (Kaakeh, 2005) with its native home in Southern Asia and Melanesia and is a serious pest of coconuts. In the Middle-East, it gained importance in the mid 1980’s when, it was first reported from UAE in 1985 and spread to the entire region including the kingdom of Saudi Arabia thereafter (Kaakeh, 2005; El-Ezaby, 1997). Currently, pheromone traps (monitors) are used as new tactics in an Integrated Pest Management (IPM) of the red palm weevil (Abraham et al., 1998). These management tactics were used in Saudi Arabia and other Middle East countries (Vidyasgar et al., 2000; Abbas et al., 2006). These methods may give good results if employed with the knowledge of refractoriness (defense mechanisms) of this insect and well studied (from all sides including lectins) against the microbial entomopathogenes e.g., Metarhizium anisopliae (Kershaw et al., 1999) and Bacillus thuringiensis (Mohan and Gujar, 2002).
Generally, all the insects (including the non-blood sucking) display diverse arrays of defense mechanisms to counteract the wide spectrum of potential pathogens and harm substances in the surrounding environment (Lackie, 1988). In fact, both cellular (haemocyte-mediated) and non-cellular or humoral (haemolymph and midgut lections) defense responses were reported in blood sucking insects against naturally-occurring pathogenic organisms such as bacteria and parasites (Ingram and Molyneux, 1988; Abdally, 1996). These defense mechanisms were studied also in non blood sucking insects like American cockroaches (Periplanata americana) and locusts (Schistocerca gregaria) by detecting the lectins (agglutinins) using agglutination activities of these substances against some mammalian RBCs. These lectins were then considered to be a part of those insects with immune response (Molyneux et al., 1986; Whitcomb et al., 1974; Lackie, 1981). In another study (of the same insects), haemagglutination activity was tested against nine types of RBC. Of these nine, human (ABO) and rabbit RBC showed the highest titres (Hapner, 1983).
Previous studies found lectins in vector (blood sucking) and non-vector (non blood sucking) insects and they may play a very important role in resistance against pathogenic organisms (Ingram and Molyneux, 1988; Abdally, 1996) beside their digestion functions (Osir et al., 1995; Lehane, 1976). These substances can be detected only by agglutinating the Red Blood Cells (RBCs) (Ingram and Molyneux, 1988; Abdally, 1996). Earlier, Croft et al. (1982), Ingram and Molyneux (1988) and Welburn et al. (1989) used relatively heat-labile, human ABO(H) blood group against samples from testes flies Glossina morsitans morsitans, G. palpalis gambiensis and G. tachinoides. They reported that non-specific lectins or lectin-like agglutinins were detected in their midguts and haemolymph of Glossina species (titres ranged from 2-9-2-16 in haemolymph and from 2-3-2-7 in midgut). In the present study agglutination activity was determined against human (ABO), rabbit, sheep and horse RBCs according to Ingram and Molyneux (1988) and Abdally (1996) in order to detect lectins (the immunity substances) in the midgut (MG) of the red palm weevil with a long term objective of refining this pest by identifying weak links in the immunity system for contributing in its biological and Integrated Pest Managements (IPM).
According to our knowledge and survey of literature, this study is an introductory type of investigation (first in the world) of the lectins (hemagglutinins) in the midgut (MG) of the Red Palm Weevil (Rhynchophorus ferrugineus) of different ages, sexes and nutritional status.
MATERIALS AND METHODS
The study was carried at Department of Microbiology and Parasitology, College of Veterinary Medicine and Animal Resources, P.O. Box 1757, Al-Ahsa 31982, King Faisal University, Al-Ahsa, Kingdom of Saudi Arabia during 2006-2008.
Maintenance of the weevils: Weevils were obtained from colonies maintained in the laboratory at 25-28°C in dark place at Al-Hasa National Date Palm Research Center, Saudi Arabia. Larvae were reared on a mixture of sugar canes and palm tissues in plastic containers.
MG extraction: The samples were collected from L1, L2, L3 larvae (fed and unfed) and from adult insects of 1, 5, 10, 25 and 30 days old (fed and unfed). Then, the midguts were carefully removed from 4 larvae and 4 adults dissected insects (of each sex) at each sampling time. The tissues were placed in 1 mL of cold Phosphate Buffer Saline (PBS) with a pH of 7.4 and were kept in an ice bath until the completion of dissection (every stage alone). In these experiments, MG were homogenized and used in agglutination assays because, these tissues had the strongest agglutination activity without causing haemolysis of the RBC (pilot experiment). Tissues were washed twice in cold PBS and then homogenized in chilled PBS using a 1 mL homogeniser (UNIFORM Jencons, England). The Gut homogenates were stored in l mL aliquots and left at 4°C for 3-4 h, after which the samples were resuspended, frozen and thawed three times at -20°C. The samples were then kept overnight at 4°C to permit the release of agglutinins (lectins) from tissues. The gut extracts were then centrifuged at 6000 rpm for 5 min after which the supernatant was dispensed into another aliquot (1 mL), labeled and stored at -20°C until required. Then, the concentration of this homogenate was adjusted to 4 MG mL-1 by following the methods of Ingram and Molyneux (1988) and Abdally (1996).
Agglutination assays
Preparation of RBC suspension
Obtaining the blood samples: The required volumes of blood (2-5 mL) were taken aseptically from stocks of fresh rabbit and human (A, AB, B and O) blood. Rabbit blood was obtained via syringe (containing anti-coagulant) from the ear vein. While, horse and sheep blood were obtained from the jugular vein. Human blood was obtained from King Fahd Hospital, Hofuf. The samples were tested for the presence of rouleaux formation and intra-erythrocytic haemoglobin crystals. Rouleaux formation is found in the thicker regions of blood films and wet preparations of whole blood. It appears as a face to face superimposition of the cells into stacks of various lengths and can be mistaken for agglutination, while the intra-erythrocytic haemoglobin crystals can disrupt the spreading of the blood film and makes the red cells appear as they are agglutinated (Hawkey and Dennett, 1989). The blood samples, which showed abnormalities, were rejected. The erythrocyte suspensions were prepared by treating the samples according to the method described by Kabat (1971) and Abdally (1996).
Adjustment of RBC suspension: Each blood sample was centrifuged (10-15 min) and the supernatant was discarded. The pellets were washed in 5-10 volumes of isotonic PBS with a pH of 7.4 for 3-5 times (until the supernatant appeared very clear). The washed erythrocytes were then resuspended in approximately 19 volumes of PBS with a pH of 7.4, in order to achieve an approximate 5% suspension. Then, 1 mL of the suspension was lysed with 14 mL of 0.1% aqueous solution of anhydrous sodium carbonate. The Optical Density (OD) of the clear lysate was measured against a water blank using a Beckman spectrophotometer/ Ultraspec II with a lcm cuvette at wavelength of 541 nm. An OD of 0.7 corresponds approximately to 1×109 RBC mL-1 of 5% cell suspension. The original suspension was then adjusted to 5% using the Kabat’s formula:
 |
These standardized erythrocyte suspensions were labeled and well-stoppered flasks and kept in the refrigerator at 4°C until required. Later on, 0.1% suspensions were prepared at the time of each assay according to procedure of Abdally (1996).
Running the assays
Dilution of the homogenates: Double dilutions of Midgut Homogenates (MG) were performed with PBS in microtitre plates (96 wells). In order to make sure that the assays were done correctly, the first experiment was done by using the lectin concanavalin A (Con A) from Ccmvalla ensiformis, obtained from Sigma. The Con A was used as a positive control by following the methodology of Msangi (1988) and Abdally (1996). It was reported that Con A was found to agglutinate 2% fresh human erythrocytes in 0.1 M PBS with a pH of 6.8 at 25 mg mL-1 (Sigma). However in the laboratory, it was found to agglutinate 0.1% suspensions of rabbit erythrocytes and human (ABO), sheep and horse erythrocytes in 0.1 M PBS with a pH of 7.4. The haemagglutination activities of MG samples were compared to Con A (positive control). Assays were carried out in 96 well plates (Dynatech). The first well from every sample row was used for the neat sample. In these assays dilution started from 2-1 (well 2) to 2-12 (well 12) for each sample. Five microliter of PBS was added to all the wells in the rows except well 1 in rows A, B, D, E, G and H in which 10 μL of the sample (neat) was added. Doubling the dilutions of the samples was then performed by the transference of 5 μL of the neat sample from well 1 to the diluent (PBS) in well 2. The solution was mixed thoroughly. After changing the tip, 5 μL of this diluted solution was transferred to the next 5 μL of the diluent (PBS) in well 3 and the process was repeated up until well 12. This process was performed for all samples. Every sample of HL and MG was tested in a single plate alone. The plates were covered and incubated at 25-26°C for 2 h after which the haemagglutination activity was determined.
Detection the degree of the agglutination activities: Equal volumes of the adjusted RBCs were added to all wells. The plates were then incubated at 25-26°C for two hours and examined for haemagglutination. The degree of agglutination was usually assessed using a seven score relative scale according to Cunningham and Vickerman (1992), Ingram and Molyneux (1988) and Abdally (1996). 100% agglutination (3+), 75% (3+/2), 50%(2+/l), 25%(2+/+), IO-2O%(1+), trace (tr) and no agglutination (0). The dilutions which just failed to give visible agglutination were regarded as the end points (titres). They were expressed as -Iog2 2-n. The controls comprised PBS alone against each sample and all the experiments were duplicated. The results were treated by using the Analysis of Variance (ANOVA) technique.
RESULTS AND DISCUSSION
The results are expressed in Table 1 (for larval stages), Table 2 (adult males fed and unfed) and Table 3 (adult females fed and unfed).
Most susceptible RBC types: The results indicated (Table 1-3) that the rabbit RBCs are the most sensitive cells (susceptible indicators) as these gave the highest titers (p<0.05) followed by human RBCs group B, human RBCs group O, horse RBCs, human RBCs group A and AB and the sheep RBCs (the lowest titers).
Effect of age, sex and nutrition
Larval stages: There were no significant differences in the activities of L1, L2 and L3 larvae (fed and unfed) of each stage against each RBC indicator (Table 1). However, it was found that the activity increased in the advanced larval stages. This is clearly shown in case of rabbit RBCs where, L1(3), L2(5) and L3(5) and human RBCs group B where L1 (2), L2 (4) and L3 (4). While, assays against the other RBC types almost gave less titers.
Adult insects: The results in Table 1 and 2 clearly indicated, especially with rabbit and human B RBCs, that the old insects gave higher activities (p<0.05) than the young insects (p>0.01). Similarly, the activities of males (p<0.05) were higher than the females (p>0.01). It was also observed that the activities showed increases when the insects were well fed (p<0.05).
| Table 1: | The agglutination activity titration (-log 2-n) of MG of Red Palm Weevil (Rhynchophorus ferrugineus) larval stages (fed and unfed) against mammalian RBC used in the investigation |
 | |
| L: Larval stage (both fed and unfed larval stages gave the same results) | |
| Table 2: | The effects of age and nutritional status on agglutination activity titration (-log 2-n) of MG from adult male Red Palm Weevil (Rhynchophorus ferrugineus) fed and unfed against mammalian RBC used in the investigation |
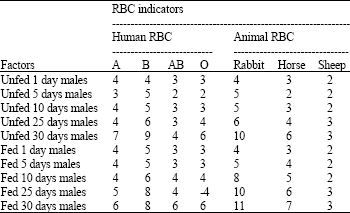 | |
| Table 3: | The inflwenc of age and nutrion on the agglutination activity titration (-log2-n) of MG from female Red Palm Weevil (Rhynchophorus ferrugineus) against mammalian RBC used in the investigation |
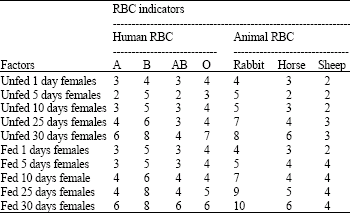 | |
This behavior is clear especially in the results of the assay against the rabbit RBCs (indicators) (Table 2, 3).
The results of this study showed that there are substances in the midgut (MG) of the Red Palm Weevil (RPW) which are responsible for the agglutination of the Red Blood Cells (RBCs) and these might be lectins. The rabbit RBCs were the most agglutinating indicators (p<0.05). These RBCs were similar to those found in stable flies S. calcitrans haemolymph (HL) midgut (MG) (Abdally, 1996). It has been observed in various insects that if the HL and MG lectins were from the same insect, they would have the same RBCs indicators and played significant roles in that insect immunity (Abdally, 1996). Therefore, for this reason, the rabbit RBCs were strongly recommended for use only in investigating the HL lectins of the Red Palm Weevil (RPW), Rhynchophorus ferrugineus. The strength of the hemagglutination activities of an insect lectins might be influenced by its food types and the tissue extractions. This was seen clearly in case of the blood sucking stable fly, S. calcitrans, where the maximum agglutination activity was observed with homogenates taken only from the reservoir zone of the midgut (MG), while the whole midgut (MG) extracts containing the opaque and the lipoid zones caused less agglutination than the reserve zone. This is besides the hemolysis the tested RBCs (Abdally, 1996). This was observed also with tsetse flies Glossina morsitans morsitans, G. palpalis gambiensis and G. tachinoides (Ingram and Molyneux, 1988). However, in the present work, the activity of whole MG tissues of RPW was higher than the blood parasitic flies. This could be attributed to the food types of RPW. In fact, the RPW feeds on tissues of palms containing a lot of sugars and they need this agglutinins to accumulate in order to be digested, while blood sucking flies need them to prevent them from pathogenic organisms found in ingested blood as innate immunity (Boulanger et al., 2002), beside helping them in digestion (Abdally, 1996). This is not surprising as there were other reports of hemolysis occurring with the use of whole midgut extracts (Spates and Deloach, 1980; Spates, 1981). Lehane (1976) reported that the opaque zone midgut cells of S. calcitrans were responsible for the production and release of proteolytic digestive enzymes into the gut lumen via Membrane Bound Vesicles (MBV). Later, Blakemore et al. (1993) reported that the trypsin enzyme in the opaque zone was stimulated by cyclic AMP but not cyclic GMP. Therefore, it was well known that the peritrophic membrane (pm) functions as a physical barrier (a primary barrier) which separates blood contents and parasites from the midgut epithelial cells (Dunn, 1986; Lehane, 1991). However, it was reported that trypanosomes can penetrate the peritrophic membrane 9-11 days after ingestion of infective blood (Ellis and Evans, 1977). This indicates that the peritrophic membrane is not a physical barrier to the penetration of trypanosomes to the haemocoel of the infected insects (Lehane and Msangi, 1991). Generally, the lectins are known to play an important role as humoral defence factors in the immune mechanisms of insects (Dunn, 1986; Lackie, 1988). They were reported in the haemolymph of G. m. morsitans and had anti-parasitic agglutination activity against T. brucei, T. vivax and T. congolense in vitro (East et al., 1983). The agglutination activity was reported also in the haemolymph mid and hind guts of G. m. morsitans and G. austeni against T. brucei and RBCs of several animal species. In some studies, the insect lectins (agglutinins) were found to have a selective reactivity against D(+)-glucosamine (Ibrahim et al., 1984, Mohamed et al., 1992). Hemagglutination activity was reported in gut and hemolymph extracts of G. morsitans, G. palpalis gambiensis and G. tachinoides (Ingram and Molyneux, 1988) and hemolymph of G. fuscipes fuscipes (Ingram and Molyneux, 1990) against human erythrocytes (RBC) of ABO (H) blood groups. These findings supported our investigation on the immunity of the RPW MG lectins against all the tested RBCs where the highest titers were, seen in case of the rabbit RBCs, 2-5 -2-12 (p<0.05). Therefore, the rabbit RBCs should be used alone for detection of RPW HL and Mg lectins besides detecting their specific carbohydrate inhibitors. There is another thing which is very important in the digestion and defence of insects which should be investigated in this weevil. It is the peritrophic membrane (pm). Abdally (1996) investigated this in stable flies following the same method and found results similar to those of Lehane and Msangi (1991), who studied extracts of whole gut, peritrophic membrane and midgut remains in order to investigate the hemagglutination activity towards human erythrocytes of AB Rh -ve blood group. They found that there was no activity in the extracts of whole gut of newly-emerged G. m. morsitans flies and only a trace of agglutination was observed in flies aged 3-8 days post-emergence (p.e.) (titres neat -2°) whilst insects aged 14 days p.e. had the activity of 50% (2+) and titres ranged neat -2-8. The same RBC type was found to be agglutinated by midgut (MG) extracts of G. m. morsitans (titre 2-7), G. palpalis gambiensis (titres 2-3) (Ingram and Molyneux, 1988). However, larvae and young adult (1-10) RPW showed some agglutination activities (p>0.01) almost against all the RBCs used. This might be due to the weevil’s food types which are obtained from plant tissues (e.g., Rice, sugar canes, Palm trees etc.) that are used as typical sources of carbohydrates for men and animals. In fact this will be more clear if the investigation of the pretrophic membrane of the weevil gut and the most specific inhibitor sugars of its agglutinins (lectins) of MG and HL were done. Because, it will elaborate the defense mechanisms of this economical important insect especially if these studies are carried with the biological insecticides containing the entomopathogenes (the natural enemies of the insects) beside the chemical insecticides in order to determine their integrated effects against the RPW. It will also support the Integrated Management (IM) of the Red Palm Weevil in Saudi Arabia and other Middle East Countries.
ACKNOWLEDGMENTS
This cooperative study was carried by The Date Palm Research Center (Center of Excellence) of King Faisal University (KFU) and Al-Hasa National Date Palm Research Center, Ministry of Agriculture under the Memorandum of Understanding between KFU and The ministry. Thanks are extended to the Hematology Department, King Fahd Hospital, Hofuf for providing the healthy human blood types. The authors appreciate the support and encouragement received from Mr. Adnan Al-Afaleg, Director, Al-Hasa National Date Palm Research Centre. Thanks are also extended to Prof. Dr. S. Al-Eid, Director, Date Palm Research Center (Center of Excellence), King Faisal University and Dr. Abdul Kader Sallam and their colleagues for their supports. Authors also appreciate the useful information and cooperation received from Dr. Jameel Al-Khayri (Member of Shura Council), Prof. Dr A. AL-Ajlaan (KFU) and Dr. J. R. Faleiro (Entomology ICAR Research Complex For Goa, India). Finally, they wish to express their great appreciation to their colleagues Prof. Dr. M. Abdul-Azeeze and Dr. M.J. Al-Hajar (KFU) for their help in statistical analysis and useful comments for improving this manuscript.
REFERENCES
- Dunn, P.E., 1986. Biochemical aspects of insect immunology. Ann. Rev. Entomol., 32: 321-339.
CrossRef - Ibrahim, E.A., G.A. Ingram and D.H. Molyneux, 1984. Haemagglutinins and parasite agglutinins in haemolymph and gut of Glossina. Tropenmedezin unde Parasitol., 35: 151-156.
PubMedDirect Link - Kershaw, M.J., E.R. Moorhouse, R. Bateman, S.E. Reynolds and A.K. Charnley, 1999. The role of destruxins in the pathogenicity of Metarhizium anisopliae for three species of insect. J. Invertebrate Pathol., 74: 213-223.
CrossRefDirect Link - Mohamed, H.A., G.A. Ingram and D.H. Molyneux, 1992. Carbohydrate-binding specificities of anti-erythrocyte lectins (haemagglutinins) in Anopheles gambiae gut extracts and haemolymph. Med. Vet. Entomol., 6: 217-224.
PubMed - Mohan, M. and G.T. Gujar, 2002. Geographical variations in larval susceptibility of the diamondback moth, Plutella xylstella (Lepidoptera: Plutellidae) to Bacillus thuriniensis spore-crystal mixture and purified crystal proteins and associated resistance development in India. Bull. Entomol. Res., 92: 488-498.