
K. Khucharoenphaisan
Department of Microbiology, Faculty of Science, Kasetsart University, Bangkok 10900, Thailand
S. Tokuyama
Department of Applied Biological Chemistry, Faculty of Agriculture, Shizuoka University, 836 Ohya, Shizuoka 422-8529, Japan
K. Ratanakhanokchai
School of Bioresources and Technology, King Mongkut�s University of Technology, Thonburi, Bangkok 10140, Thailand
V. Kitpreechavanich
Department of Microbiology, Faculty of Science, Kasetsart University, Bangkok 10900, Thailand
Pakistan Journal of Biological Sciences
Year: 2010 | Volume: 13 | Issue: 5 | Page No.: 209-215







ABSTRACT
The effect of carbon sources on the production of β-xylanase by Thermomyces lanuginosus TISTR 3465 was investigated. Xylan showed the highest inductive effect on the enzyme formation whereas xylobiose and xylooligosaccharides resulted in lesser effect. β-Xylanase was also produced at low level with xylose as well as other sugars tested. Xylan concentration at 15 g L-1 gave the maximum inductive effect on β-xylanase formation, whereas xylooligosaccharides and xylose were effective at a lower concentration of 2.5 g L-1. High concentrations of these sugars significantly repressed the enzyme formation. Crude enzyme from the supernatants, without and with other sugars produced a single xylanase band on non-denaturing PAGE gels. However, an intense xylanase activity band was observed from the supernatant of media with xylan, xylobiose and xylooligosaccharides as the carbon sources. An intense protein band of 24.9 kDa from the culture filtrate of xylan medium was observed. Xylan increased β-xylanase production by the fungus 16-fold when it was added to the xylose medium after cultivation for 3 days. In contrast, addition of xylose to the xylan medium decreased β-xylanase production 3-fold. A distinct appearance and disappearance of a 24.9 kDa protein and the activity band coincided with an increase and decrease of xylanase activity, respectively. This indicated the synthesis of xylanase by this strain was most induced by xylan. Moreover, the level of xylanase induction has no related to amino acid sequence of the enzyme.
PDF Abstract XML References Citation
How to cite this article
K. Khucharoenphaisan, S. Tokuyama, K. Ratanakhanokchai and V. Kitpreechavanich, 2010. Induction and Repression of β-Xylanase of Thermomyces lanuginosus TISTR 3465. Pakistan Journal of Biological Sciences, 13: 209-215.
DOI: 10.3923/pjbs.2010.209.215
URL: https://scialert.net/abstract/?doi=pjbs.2010.209.215
DOI: 10.3923/pjbs.2010.209.215
URL: https://scialert.net/abstract/?doi=pjbs.2010.209.215
INTRODUCTION
Microbial xylan-degrading enzymes are potentially important in the biobleaching process of pulp and paper, improving the quality of animal feed, increasing the quality of bread and the conversion of lignocellulose to fermentable sugars for the production of chemicals and fuels (Jäger et al., 1992; Viikari et al., 1993; Sunna and Antranikian, 1997; Beg et al., 2001). β-Xylanase (Endo 1,4-β-D-xylan xylanohydrolase, EC 3.2.1.8) is one of the important enzymes involved in xylan degradation working synergistically with other debranching enzymes (Yasui et al., 1984; Rapp and Wangner, 1986; Hrmova et al., 1991). Numerous bacteria, streptomycetes and fungi can degrade xylan by producing a range of enzymes (Abdeshahian et al., 2009). Thermomyces lanuginosus is a potent thermophilic strain which produces a single cellulose-free endo β-xylanase (Kitpreechavanish et al., 1984; Anand et al., 1990; Singh et al., 2000, 2003). Xylan, xylose, xylobiose and non-metabolizable β-xylosides known to be common inducers of xylanase and the xylanase produced by T. lanuginosus DSM 5826 was most stimulated by xylose (Purkarthofer and Steiner, 1995). Our early reported that strains of T. lanuginosus could be differentiated into two groups based on their ability to produce xylanase using xylan or xylose as sole of carbon source. One group of these strains produced high xylanase activity either in the medium using xylan or xylose as a sole carbon source. The xylanase production by T. lanuginosus ATCC 44008, THKU-11 and THKU-25, which were the representative members of this group, increased when xylose was added to the xylan medium. In contrast, there was another group that produced high xylanase activity only in the xylan medium. Addition of xylose to the xylan medium resulted decreasing of xylanase production in T. lanuginosus ATCC 46882, TISTR 3465 and THKU-85 that belonged to this group (Khucharoenphaisan et al., 2009). In this study, the induction and repression of β-xylanase production of T. lanuginosus TISTR 3465 in liquid culture and resting cell condition were examined. Furthermore, protein patterns and activity of β-xylanase on gel electrophoresis were investigated during the induction and repression of the enzyme.
MATERIALS AND METHODS
Microorganism and cultivation: A thermophilic fungus, Thermomyces lanuginosus (formerly known as Humicola lanuginosa) (Domsch et al., 1993), used in this study was a generous gift from Prof. Tsutomu Morinaga. The strain is deposited in the MIRCEN Bangkok culture collection, Thailand as T. launginosus TISTR 3465. Xylanase was produced in 50 mL of basal medium in 250 mL Erlenmeyer flasks. The medium consisted of (per liter of distilled water) 2.0 g KH2PO4, 0.3 g MgSO4.7H2O, 0.3 g CaCl2, 5.0 g peptone, 3.0 g yeast extract and 10 g of each appropriate carbon sources; arabinose, cellobiose, glucose, galactose, xylose, xylobiose, xylooligosaccharide and oat spelt xylan. Initially, pH of medium was adjusted to 6.0 and autoclaved for 20 min at 121°C. The different mediums were inoculated with 105-106 spores of 10 days T. lanuginosus grown on wheat bran agar medium and then incubated at 45°C on reciprocal shaker at 120 strokes min-1 (JEIO TECH, model BS-30). Moreover, various concentrations of xylan, xylooligosaccharides and xylose were also used as carbon source.
Induction and repression on β-xylanase production: The fungus was grown in basal medium supplement with xylose or glucose as mention earlier. Oat spelt xylan suspension was added to the culture on the 3rd day of cultivation to make a final concentration of 10 g L-1 and further incubated. In parallel, the fungus grew in basal medium containing oat spelt xylan and then xylose or glucose was added on the 3rd of cultivation. The culture supernatant was collected by centrifugation and used as enzyme solution for determination of β-xylanase activity, soluble protein content, protein pattern and activity on gels by electrophoresis.
The freshly washed glutamic acid-grown mycelium were suspended in 50 mL sodium phosphate buffer (pH 6.0) and the aliquot (15 mL) to make a final concentration of mycelium was about 0.11 g dry weight. The inducers, oat spelt xylan or xylose was rapidly mixed with mycelium suspension (final concentrations are 10, 100, 500 and 1000 mg L-1). A control without any inducer was also set up. The mixtures were incubated at 50°C on a reciprocal shaker at 120 strokes min-1. Aliquots were taken at different time intervals (10 h) and filtrated supernatant was used as enzyme solution for determination of xylanase activity.
Sequencing of xylanase gene fragment: Total RNA of T. lanuginosus TISTR 3465 was isolated and used as template for complementary DNA synthesized by using Qiagen® One Step RT-PCR Kit. The PCR product was cloned to pGEM-T easy vector following standard procedures. The plasmid DNA was isolated from transformant by Quantum Prep Plasmid Miniprep Kit (Bio-Rad, USA). The PCR products were sequenced using Big Dye® Terminator V3.1 cycle sequencing kit (Applied Biosystems). Sequence gel electrophoresis was carried out and sequences obtained by a 5500-5 DNA sequencer (Hitachi). The nucleotide sequences were translated to amino acid and compared with other xylanase using the Genetyx version 5.0 program (Khucharoenphaisan et al., 2008). The phylogenetic tree was constructed by using the neighbor-joining method in MEGA version 4 software. The topology was evaluated by bootstrap analysis based on 1000 resamplings (Felsenstein, 1985).
Analysis
Determination of dry weight: Culture broths were filtered using Whatman filter paper No. 4 (Whatman International Ltd., USA). Mycelium retained on the Whatman filter paper was then washed and dried at 105°C to a constant weight.
Determination of β-xylanase activity: β-xylanase was assayed by determination of reducing sugars liberated from oat spelt xylan when incubated with the diluted enzyme solutions at pH 6.0 and 50°C for 10 min. The reducing sugars were measured using the DNS reagent (3,5-dinitrosalisylic acid) (Miller, 1959). One unit of β-xylanase activity was defined as the amount of enzyme that produced 1 μmole of xylose in 1 min.
Determination of protein: Protein was determined using the method of Lowry et al. (1951) with bovine serum albumin as a standard.
Determination of molecular weight and xylanase activity band: Molecular weight of proteins in supernatants was determined by Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE) following the method of Laemmli (1970). Protein and xylanase activity patterns were performed with Polyacrylamide Gel Electrophoresis (PAGE) (Mishra and Maheshwari, 1996) and stained with Coomassie brilliant blue R-250. For active zymogram analysis, the gels were incubated in a 1% oat spelt xylan solution for 1 h at 50°C. The gels were stained with 1% Congo red (Sigma Co., USA) for 15 min and destained by 1 M NaCl until excess dye was removed from the xylanase activity band. The gels were next immersed in 0.5% acetic acid and the background turned dark blue and the xylanase activity bands appeared clear colorless areas (Kyu et al., 1994).
Chemicals: All chemicals used were analytical grade. The xylan used for induction studies was oat spelt xylan (Sigma Co., USA). Xylooligosaccharides (Suntory Limited Osaka, Japan Lot. No. 92072511) contained 22.2% xylose, 55.4% xylobiose, 20.3% xylooligotriose, 2.1% glucose and 24.6% moisture content.
Place and during time: This study was conducted from 2008 to 2009 at Enzyme Technology Laboratory of Faculty of Science, Kasetsart University, Thailand.
RESULTS AND DISCUSSION
Effects of various sugars on β-xylanase production from T. lanuginosus TISTR 3465: The effect of the sugars such as arabinose, cellobiose, glucose, galactose, xylose, xylobiose, xylooligosaccharides and oat spelt xylan on the production of β-xylanase was investigated. Oat spelt xylan stimulated the highest level of β-xylanase production compared to xylooligosaccharides, xylobiose and xylose which had a lesser effect (Table 1). A low level of β-xylanase activity (<1U mL-1) was detected in the media containing other sugars as a carbon source and a control without addition of sugar. Furthermore, an intense band of xylanase activity was observed on zymogram of the culture supernatant containing xylan, xylobiose and xylooligosaccharides.
It is clearly indicates that oat spelt xylan is the best carbon source for stimulate the production of extracellular xylanase by this fungus. However, low level of β-xylanase activity was detected in the presence of other inducing substances and also in non carbon source and the presence of easily metabolizable substrates such as glucose, cellobiose, arabinose and galactose. This finding leads to the conclusion that the β-xylanase was constitutively produced at low level by T. lanuginosus TISTR 3465 and could be induced by xylan.
| Table 1: | β-Xylanase activities of T. lanuginosus TISTR 3465 produced on different carbon sources after 5-day cultivation |
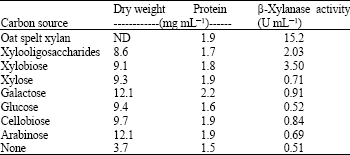 | |
| ND: Not-detected | |
The results are agreement with the earlier reports where it was found that xylanase production from T. lanuginosus DSM 5826 (Purkarthofer and Steiner, 1995) and Trichoderma harzianum (Ahmed et al., 2003) were induced by xylan and repressed by glucose.
Effect of xylan, xylooligosaccharides and xylose concentration on β-xylanase production: The effect of xylan concentration and its sugar derivatives on enzyme production was investigated. It was found that β-xylanase activity gradually increased with the increase of xylan concentrations up to 15 g L-1 but it was decreased when high concentration (Fig. 1). Low concentrations of xylooligosaccharides and xylose also had a similar effect on xylanase production but in less extent. Maximum β-xylanase activity was obtained when concentration of xylooligosaccharides and xylose was at 2.5 g L-1 (Fig. 1).
It was possible that a small amount of sugars in which was released from the hydrolysis of xylan played a role on induction but this appeared to be depend on its concentration. High concentrations of the polymer repressed synthesis of enzyme which might have resulted from the accumulation of hydrolysis products resulting in feedback inhibition of enzyme production. At the beginning of the culture, the constitutive level of xylanase activity was produced, which lead to the liberation of xylooligomers from xylan. Xylanase formation in xylan culture was extended, thus leading to a high level of enzyme activity. The reaction of this effect was believed to be the longer availability of inducing molecules that are slowly and continuously liberated. This mechanism was also suggested by Ghosh and Nanda (1994) to explain the induction of xylanase in Aspergillus sydowii MG49 by xylose.
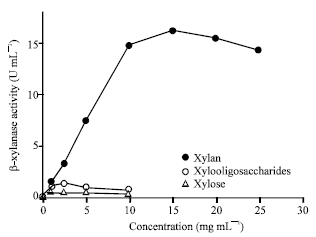 | |
| Fig. 1: | Effect of various concentrations of xylan, xylooligosaccharides and xylose on production of β-xylanase at 5-day cultivation |
These results are also similar to findings with Bacillus circulans B6 reported by Kyu et al. (1994) with the highest xylanase production observed with 10 g L-1 xylan and repression at high concentrations. The highest concentration of xylan (0.333 g L-1) induced xylanase in resting cells of T. lanuginosus DSM 5826 (Purkarthofer and Steiner, 1995). The highest xylanase activity was detected with 2.5 g L-1 of xylan in Cellulomonas flavigena (Hidalgo-Lara and Farrés, 2005). In a similar manner, the effect of concentration of xylooligosaccharides on xylanase production was almost identical to that with xylose as shown Fig. 1. Xylooligosaccharides and xylose also had an inductive effect on enzyme production at the low concentrations, 2.5 g L-1 (Fig. 1). Nevertheless, the inductive effects were less pronounced in which were compared to xylan. This indicated that readily utilized sugars such as xylose and xylooligosaccharides could be inducers at low concentrations. However, there is a certain ambiguity about whether xylooligosaccharides are direct inducers of β-xylanase. Xylooligosaccharides are hydrolyzed to xylose by β-xylosidase during transport across the cell membrane or within the cell and that the inducing effect of xylooligosaccharides and xylobiose are an effect of xylose at the molecular level.
Induction and repression effect on β-xylanases production by T. lanuginosus: A low β-xylanase activity was found when the fungus was grown in the medium containing xylose or glucose as the carbon source. However, addition of xylan to both media at the 3rd day of culture could effectively increase β-xylanase formation 16-fold (Fig. 2).
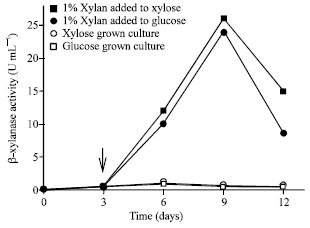 | |
| Fig. 2: | Effect of xylan addition to the xylose and glucose grown culture on the β-xylanase production by T. lanuginosus. 1% xylan added to xylose grown culture, 1% xylan added to glucose grown culture, xylose grown culture, glucose grown culture. Arrow indicated the addition of xylan |
In contrast, the β-xylanase formation dramatically decreased about 3-fold when xylose or glucose was added to the 3rd day xylan-grown culture (Fig. 3). The SDS-PAGE revealed that the main protein in the supernatant had a molecular weight of 24.9 kDa. This main protein exhibited a β-xylanase activity on zymogram in which was corresponded to main protein on SDS-PAGE and PAGE. Addition of xylose or glucose to xylan grown cultures decreased the intensity of the protein band on SDS-PAGE (Fig. 4a, lanes 2 and 3), PAGE (Fig. 4b, lanes 2 and 3) and xylanase activity bands on zymogram (Fig. 4c, lanes 2 and 3). In contrast, intensity of main protein band was increase when xylan was added to the xylose or glucose grown cultures, which detected on SDS-PAGE (Fig. 4a, lanes 5 and 7) and zymogram (Fig. 4c, lanes 5 and 7).
Effect of xylan and xylose concentration on β-xylanase induction in resting cell: At various time intervals, the xylanase activity was determined in the incubated buffer. Xylan and xylose were able to induce xylanase formation in the T. lanuginosus TISTR 3465. Xylanase were increased when the concentration of xylan or xylose was increased (Fig. 5a-c). In the control of experiments showed very low xylanase activity due to the constitutive formation of xylanase. The xylanase formation was faster in the presence of xylose than xylan but xylan show higher pronounced on xylanase induction than xylose (Fig. 5). The result was different from the report of Purkarthofer and Steiner (1995) who reported that xylose was the best inducer for xylanase production in resting cell of T. lanuginosus DSM 5826.
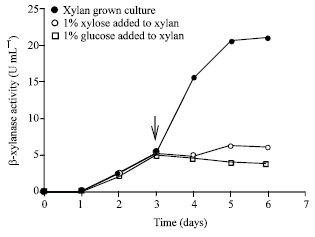 | |
| Fig. 3: | Effect of xylose or glucose addition to the xylan grown culture on the β-xylanase production by T. lanuginosus. Xylan grown culture, 1% xylose added to xylan grown culture, 1% glucose added to xylan grown culture. Arrow indicated the addition of xylose or glucose |
 | |
| Fig. 4: | Gels of SDS-PAGE (A) and Native-PAGE (B) after stained with coomassie brilliant blue R-250 and zymogram for xylanase activity (C) of crude T. lanuginosus TISTR 3465 β-xylanases produced in culture medium. Lanes: 1: xylan grown culture, 2: 1% xylose added to xylan grown culture, 3: 1% glucose added to xylan grown culture, 4: xylose grown culture, 5: 1% xylan added to xylose grown culture, 6: 1% glucose added to xylose grown culture, 7: xylan added to glucose grown culture and M: Molecular mass standards |
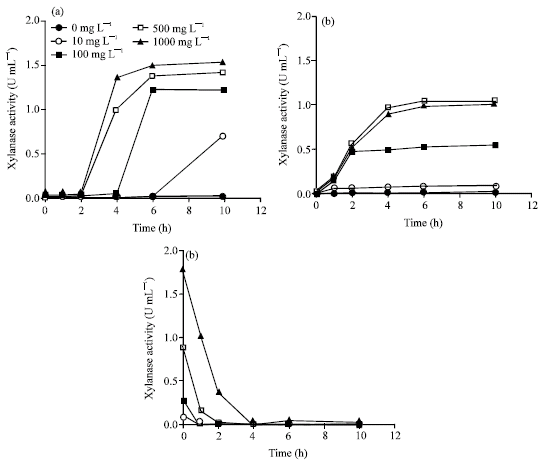 | |
| Fig. 5: | Time course of extracellular xylanase production by washed glutamic acid-grown mycelium of T. lanuginosus TISTR 3465 during incubation in (a) 50 mM phosphate buffer (pH 6.0) supplied with separate oat spelt xylan and (b) xylose at concentrations of 0 mg L-1, 10 mg L-1, 100 mg L-1, 500 mg L-1 and 1000 mg L-1. Concentrations of fungal mycelium are in range of 0.11 g dry weight/experiment. (c) The consumption of xylose from xylose-supplemented condition was also shown |
This indicated that these two strains might have different induction mechanism. However, we suggested that oat spelt xylan acted as the premier inducer of xylanase in strains of T. lanuginosus TISTR 3465.
 | |
| Fig. 6: | Alignment of amino acid sequence of xylanase from T. lanuginosus TISTR 3465 and DSM 5826. The alignment was done using the genetyx (version 5.0) alignment program. Alignment characters are indicated as follows: *Indicates position with a conserved amino acid residue and .: indicates position with a different amino acid residue |
 | |
| Fig. 7: | Phylogenetic tree of amino acid sequence analysis of xylanase of thermophile, mesophile and thermophilic fungi constructed by Neighbor-joining method from MEGA 4 program. Scale bar shown distance values under the tree means 0.05 substitutions per amino acid position. Bootstrap analyses were performed with 1000 re-samplings and percent values are shown at the branching points |
Xylanase gene sequencing analysis: The complementary DNA of xylanase genes from T. lanuginosus TISTR 3465 was sequenced. The nucleotide sequences were translated to amino acid sequence and aligned with sequence of xylanase from T. lanuginosus DSM 5826 (accession No. U35436) as shown in Fig. 6. Amino acid sequence of xylanase from strain TISTR 3465 was completely similar to that of T. lanuginosus DSM 5826. Furthermore, the sequences alignment revealed that the active site of xylanase of both strains of T. lanuginosus were the position of E117 and E209 corresponding to the previous reported of Lin et al. (1999). The phylogenetic tree showed that xylanase from TISTR 3465 strain is closed to DSM 5826 strain and related to each other thermophilic fungi (Fig. 7). This concluded that amino acid sequence has no effect on xylanase induction. It appears that other factors are dominant in induction of xylanase than that of amino acid sequence. However, xylanase production using xylose as a carbon source was related to RAPD pattern with UBC 241 primer (Khucharoenphaisan et al., 2009).
CONCLUSION
Present finding was confirmed that the formation of β-xylanase by T. lanuginosus TISTR 3465 could be induction by xylan and its sugar derivatives. However, the mechanisms have to further clarify to make more understanding whether these are different from other T. lanuginosus strains.
ACKNOWLEDGMENT
This study was financially supported by the Thailand Research Fund through the Royal Golden Jubilee Ph.D. Program and Student Exchange Support Program (Scholarship for Short-term Study in Japan)-JASSO.
REFERENCES
- Abdeshahian, P., N. Samat and W.M. Wan Yusoff, 2009. Xylanase production by Aspergillus niger FTCC 5003 using palm kernel cake in fermentative bioprocess. Pak. J. Biol. Sci., 12: 1049-1055.
CrossRefPubMedDirect Link - Ahmed, S., Qurrat-ul-Ain, A. Nighat, N. Saima, Sajjad-ur-Rahman and J. Amer, 2003. Induction of xylanase and cellulase genes from Trichoderma harzianum with different carbon sources. Pak. J. Biol. Sci., 6: 1912-1916.
CrossRefDirect Link - Anand, L., S. Krishnamurthy and P.J. Vithayathil, 1990. Purification and properties of xylanase from the thermophilic fungus, Humicola lanuginosa (Griffon and Maublanc) Bunce. Arch. Biochem. Biophys., 276: 546-553.
CrossRef - Beg, Q.K., M. Kapoor, L. Mahajan and G.S. Hoondal, 2001. Microbial xylanases and their industrial applications: A review. Applied Microbiol. Biotechnol., 56: 326-338.
CrossRefDirect Link - Felsenstein, J., 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39: 783-791.
CrossRefDirect Link - Ghosh, M. and G. Nanda, 1994. Physiological studies on xylose induction and glucose repression of xylanolytic enzymes in Aspergillus sydowii MG49. FEMS Microbiol. Lett., 117: 151-156.
Direct Link - Hidalgo-Lara, M.E. and G.S.A. Farres, 2005. β-Methyl- xyloside: Positive effect on xylanase induction in Cellulomonas flavigena. J. Ind. Microbiol. Biotechnol., 32: 345-348.
CrossRefDirect Link - Hrmova, M., E. Petrakova and P. Biely, 1991. Induction of cellulose and xylan-degrading enzymes of Aspergillus terreus by homo and heterodisaccharides composed of glucose and xylose. J. Gen. Microbiol., 137: 541-547.
CrossRefDirect Link - Khucharoenphaisan, K., S. Tokuyama, K. Ratanakhanokchai and V. Kitpreechavanich, 2009. A comparative study of Thermomyces lanuginosus strains on thermostable xylanase production. Afr. J. Biotechnol., 8: 1608-1614.
Direct Link - Khucharoenphaisan, K., S. Tokuyama and V. Kitpreechavanich, 2008. Characterization of the thermostability of xylanase produced by new isolates of Thermomyces lanuginosus. Sci. Asia, 34: 187-192.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Purkarthofer, H. and W. Steiner, 1995. Induction of endo xylanase in the fungus Thermomyces lanuginosus. Enzyme Microbiol. Technol., 17: 114-118.
CrossRef - Rapp, P. and F. Wangner, 1986. Production and properties of xylan-degrading enzymes from Cellulomonas uda. Applied Environ. Microbiol., 51: 746-752.
PubMed - Singh, S., B. Pillay and B.A. Prior, 2000. Thermal stability of β-xylanases produced by different Thermomyces lanuginosus strain. Enzyme Microbiol. Technol., 26: 502-508.
CrossRef - Sunna, A. and G. Antranikian, 1997. Xylanolytic enzymes from fungi and bacteria. Crit. Rev. Biotechnol., 17: 39-67.
CrossRef - Yasui, T., B.T. Nguyen and K. Nakanishi, 1984. Inducer for xylanase production by Cryptococcus flavus. J. Ferment. Technol., 62: 353-359.
Direct Link