
D. Yonli
Institut de l�Environnement et de Recherches Agricoles, 04 B.P. 8645 Ouagadougou 04, Burkina Faso
H. Traore
Institut de l�Environnement et de Recherches Agricoles, 04 B.P. 8645 Ouagadougou 04, Burkina Faso
P. Sereme
Institut de l�Environnement et de Recherches Agricoles, 04 B.P. 8645 Ouagadougou 04, Burkina Faso
D. E. Hess
Merry Lea Environmental Center of Goshen College, Wolf Lake, IN, USA
P. Sankara
Universite de Ouagadougou, Unite de Formation et de Recherche en Sciences de la vie et de la terre, B.P. 7021, Ouagadougou, Burkina Faso
Pakistan Journal of Biological Sciences
Year: 2010 | Volume: 13 | Issue: 5 | Page No.: 201-208







ABSTRACT
Striga hermonthica (Del.) Benth. is an important constraint to cereal crop production in Burkina Faso, of which sorghum (Sorghum bicolor L. Moench) is the most important component. Native Fusarium species to use as bio-control agents to S. hermonthica has been investigated. Fifty one Fusarium isolates obtained from diseased plants of S. hermonthica were evaluated for their pathogenicity against Striga under controlled environmental conditions. Of 51 Fusarium isolates, 14 were pathogenic to S. hermonthica but their virulence differed. These 14 isolates were evaluated for their effects on Striga seed germination in the laboratory and their ability to kill emerged Striga plants growing in greenhouse pots. Spores of Fusarium sp. isolates 150a-M, 125b-Za, 6-Fa, Fusarium equiseti isolates 5-Kou, 31-Kom, 32-Or, 13-Ba and Fusarium oxysporum isolate 34-Fo reduced Striga germination by 78 to 96% compared to the untreated control. The study showed that at the rate of 33 mg mL-1, metabolites of Fusarium sp. isolates 125b-Za, 6-Fa, F. equiseti 5-Kou and F. oxysporum 34-Fo prevented Striga seed germination. In addition to these four isolates, Fusarium sp. isolates 141b-O, 150a-M and F. equiseti isolate 32-Or were effective at 67 mg mL-1. Percentage of Striga mortality ranged from 17-37% between 14 and 28 days after inoculation with spores of F. oxysporum 34-Fo and F. equiseti 5-Kou. Striga dry biomass was reduced by 84 and 78% for the respective isolates compared to the untreated control with Striga. Sorghum yield was improved by 84 and 99% with Fusarium sp. 6-Fa and F. oxysporum 34-Fo, respectively, compared to the control without Striga. The use of Fusarium spores and metabolites against Striga offers different possibilities of bio-herbicides formulation that can be combined with other controls methods in the integrated Striga management. Further studies will be carried out under field conditions to assess the efficacy and safety of these Fusarium isolates to environment and humans and evaluate low cost strategies for transfer to subsistence farmers.
PDF Abstract XML References Citation
How to cite this article
D. Yonli, H. Traore, P. Sereme, D. E. Hess and P. Sankara, 2010. Pathogenicity of Fusarium isolates to Striga hermonthica in Burkina Faso. Pakistan Journal of Biological Sciences, 13: 201-208.
DOI: 10.3923/pjbs.2010.201.208
URL: https://scialert.net/abstract/?doi=pjbs.2010.201.208
DOI: 10.3923/pjbs.2010.201.208
URL: https://scialert.net/abstract/?doi=pjbs.2010.201.208
INTRODUCTION
Among parasitic weeds, Striga hermonthica (Del.) Benth. is the most widespread and destructive in sub-Saharan Africa (Haussmann et al., 2000). Striga hermonthica is virulent on sorghum (Sorghum bicolor L. Moench), pearl millet (Pennisetum glaucum L. R.Br.) and maize (Zea mays L.), the main staple food crops in many parts of the world (Parker and Reid, 1979; Hess et al., 1992). Striga causes severe losses in crops and more than 75% of its overall damage occurs to the host before parasite emergence from the soil (Parker and Riches, 1993). In sub-saharan Africa, yield losses due to Striga in sorghum, pearl millet and maize vary from 8 110 000 to 8 520 000 t, corresponding to 39-45% of total production (Gressel et al., 2004). In West Africa, yield losses from damage by S. hermonthica range from 24 to 27% (6 555 000 to 6926 000 t) in sorghum and millet whereas, in Burkina Faso they are estimated to 35-40% (710 000-820 000 t) (Gressel et al., 2004). Yield losses due to Striga are so tremendous that they reach 100% in heavily infested fields (Ejeta and Butler, 1993). Severe Striga infestation correlated to poor soil fertility, low rainfall and lack of production inputs contribute to important yield losses and then constitute big issues for subsistence farmers (Vogler et al., 1996). Striga species are difficult to eradicate because of the unique adaptation of the parasite to its environment and the complexity of the host-parasite-environment relationship (Vogler et al., 1996).
Striga species produce a large number of seeds with prolonged viability and special germination requirements (Ejeta and Butler, 1993; Siame et al., 1993).
Several control methods have been evaluated against Striga. These methods include the use of cultural practices, fertilizers, resistant crop varieties (Ejeta and Butler, 1993), herbicides (Carson and Kunjo, 1991; Lagoke et al., 1991; Traoré et al., 2001). However, it is still difficult to find cultivars with complete resistance (Ejeta and Butler, 1993). The availability and cost of the fertilizers and herbicides are the main constraints to their widespread use in Africa. Moreover, used alone, none of these methods has given satisfactory suppression of the parasite (Ciotola et al., 1995). Striga management strategies should lead to destruction of both the soil seed bank and emerged Striga and before the parasite produces seed (Haussmann et al., 2000; Schaub et al., 2006).
Spores of some fungi were reported to prevent Striga germination (Sauerborn et al., 1996; Kroschel et al., 1996) and this inhibition can be mechanical and/or chemical. Inhibition of Striga seed germination by phytotoxic compounds of some Fusarium species was reported by Zonno et al. (1996), Sugimoto et al. (2002) and Idris et al. (2003). In Nigeria, Striga infestation was reduced by application of spores of F. oxysporum isolates DCP and PSM-197, F. acuminatum, F. equiseti and Fusarium sp. isolate PSM-297 (Marley et al., 1999). A bio-control approach should give preference to a biological endogenous agent because of its adaptation to the ecological conditions and in order to reduce the risks associated with introducing organisms from other areas. However, there is need for improved bio-herbicide formulations and delivery systems to enhance the potential role of bio-control in integrated management of Striga in the subsistence farmer’s level.
The objectives of this study were to: (1) evaluate the pathogenicity of Fusarium isolates against S. hermonthica and (2) determine methods of application.
MATERIALS AND METHODS
The collecting of plant material started in 1998. Two year (2001-2002) study was conducted at the Kamboinsé Research Station (01°33' E, 12°28' N and 300 m altitude) of Environmental and Agricultural Research Institute in Burkina Faso.
S. hermonthica seeds: Striga hermonthica seeds were harvested in 1998 from a sorghum field located at the Kouaré Agricultural Research Station (11°95'03''’N and 0°30'58'' E) in Eastern Burkina Faso, air dried and stored in the laboratory at ambient temperature (30°C). Seeds were surface sterilized in 70% ethanol for 3 min, followed by 5 min in 3% sodium hypochlorite (NaOCl) with Tween 80, before use in germination tests.
Fusarium isolates: Diseased S. hermonthica plants were collected in 1999 from 15 locations in three agro-ecological zones (Sahelian, North-sudanian and South-sudanian) of Burkina Faso. Pieces of Striga plants showing disease symptoms were washed with sterile distilled water and surface sterilized with 90% ethanol for 45 sec. They were then rinsed with sterile distilled water, air-dried on sterile Whatman No. 1 filter paper and plated on potato-dextrose agar amended with 0.1 g L-1 of streptomycin (PDAS). After incubation at 25-30°C for 3 to 4 days, fungi showing typical morphology of Fusarium were transferred to PDAS for purification. From the collection, 42 isolates of fungi with morphological features of the genus Fusarium (colour and shape) were obtained. Nine out of these 42 isolates were identified by the laboratory of the Danish Seed Health Centre for Developing Countries (Royal Vet. and Agricultural University, Copenhagen Thorvaldsensvej 40, DK-1871 Frederiksberg C, Denmark): Fusarium oxysporum (34-Fo) and Fusarium equiseti (5-Kou, 42-Ko, 31-Kom, 13-Ba, 21-Or, 32-Or, 17-Fo, 7-Fo). An additional nine isolates collected by Abbasher et al. (1998) were included, giving a total number of 51 isolates evaluated in this study.
Spores production: Fusarium isolates were grown on PDAS in petri dishes for 14 days. Fifteen milliliter of sterile distilled water was added to mycelium collected from each Petri dish and this mixture was ground with a mixer (Janke and Kunkel, IK labortechnik, Ultra-turrax T25). The suspension was filtered through a 75 μm sieve and adjusted with sterile distilled water to 106 spore mL-1 using a haemocytometer of Malassez (HERKA®, depth = 0.2 mm, area = 0.0025 m2).
Metabolites production: Metabolites were extracted from a liquid culture of different Fusarium isolates. From a pure culture of each isolate on PDAS, three plugs (4 mm diameter) were placed in a 250 mL glass beaker with 100 mL of Potato Dextrose Broth as growth medium. The glass beaker was stirred on an agitator (Edmund Bühler, 7 400 Tübingen, SM 25) at 120 rpm for 30 days and each solution was then filtered through a Whatman® filter paper. The filtrate was freeze-dried.
Evaluation of the bioherbicidal activity of Fusarium isolates
Application of Fusarium spores: Spores of 51 Fusarium isolates were placed on Striga seeds using the technique described by Kroschel et al. (1996). Sterilized Striga seeds (95% germination rate) were pre-conditioned in a flask with sterile distilled water for 4 days. About 100 preconditioned Striga seeds were placed on a 1 cm2 glass microfibre filter paper disc (Whatman GF/A) in a sterile 9-cm petri dish lined with two Whatman No. 1 filter paper discs.
For each isolate, a suspension of 10 mL (106 spore mL-1) was used to condition pre-conditioned Striga seeds in the petri dish. Seeds were then sprinkled with 0.05 g of sorghum flour and the Petri dish sealed with Parafilm M® barrier film (Pechiney Plastic Packaging, Chicago, IL, 60631). Dishes were wrapped in aluminum foil and black polyethylene and incubated in darkness at 28°C for 10 days.
After incubation for 10 days, Striga seeds were washed with sterile distilled water through two sieves, the first (315 μm) allowing separation of seeds from the mycelium and the second (80 μm) allowing separation of seeds from spores. Stimulation of germination of Striga seeds was evaluated in a modification of the technique of Berner and Williams (1998). Striga seeds placed on 8 glass microfibre filter discs (1 cm2) (Whatman GF/A); 20 to 30 seeds disc-1. The eight discs were arranged in 4 lines around an aluminum foil ring centered in the Petri dish lined with a double Whatman No. 1 filter paper. Fourteen day-old root pieces (1 g) of sorghum variety S29 (obtained from the national breeding program and susceptible to S. hermonthica) were placed in the ring to stimulate Striga germination. The root pieces in the ring were watered with 3 mL of sterile distilled water to help diffuse root exudates. Dishes were then wrapped in aluminum foil and black polyethylene and incubated at 30°C for 48 h. The germination rate of Striga seeds conditioned in sterile distilled water (control) was then compared to those of seeds conditioned in the spore suspension of the 51 isolates. A Completely Randomized Block design (CRB) was used with three replications. The experiment was repeated three times.
Application of Fusarium metabolites: Thirty to Forty sterilized Striga seeds were placed on glass microfibre filter paper discs (6 mm diameter) in Petri dishes (7 cm) lined with double Whatman No. 1 filter papers. Six milliliter of solution of each treatment was used to condition Striga seeds and treatments were as follows:
| • | (1) Control (-), seeds conditioned with sterile distilled water |
| • | (2) Control (+), seeds conditioned with 67 mg mL-1 of growth medium filtrate freeze-dried |
| • | (3) seeds conditioned with 1 mg mL-1 of metabolite of each Fusarium isolate |
| • | (4) seeds conditioned with 10 mg mL-1 of metabolite of each Fusarium isolate |
| • | (5) seeds conditioned with 33 mg mL-1 of metabolite of each Fusarium isolate |
| • | (6) seeds conditioned with 67 mg mL-1 of metabolite of each Fusarium isolate |
Sealed petri dishes were then incubated in darkness at 27°C for 14 days. On the fifteenth day, seeds were placed on discs and transferred into new petri dishes (9 cm) lined with double Whatman No. 1 filter papers. In each petri dish, discs were arranged in 4 lines (5 discs per radius) around the aluminum foil ring. Fourteen-day old root pieces (1 g) of sorghum variety S29 were placed in the ring and moistened with 3 mL of sterile distilled water. Dishes were again sealed and incubated in the darkness at 30°C for 48 h. After 48 h, germination of Striga seed was determined. Three replications were carried out using a Completely Randomized Block design (CRB) and the experiment was repeated three times.
Leaf inoculation with Fusarium spores: Sorghum variety S29 was sown in plastic pots (26×28 cm) in soil medium made up of two parts unsterilized soil and one part of steam sterilized sand. Pots had been artificially infested with Striga seeds as described by Marley et al. (1999) in order to have an infestation rate of 5×103 viable Striga seeds per pot. Seventy Days after Sowing (DAS), emerged Striga plants were inoculated by spraying a spore suspension (1×106 spore mL-1) of Fusarium. Striga plants were inoculated in a growth chamber at 25-28°C and 100% HR and incubated there for 24 h; they were then transferred into a greenhouse.
The experimental design was a randomized complete block with three replications and the following treatments: (1) non-infested soil + sorghum, (2) soil infested with Striga seeds + sorghum ; 3 to 16) soil infested with Striga seeds + sorghum + 10 mL suspension (1×106 spore mL-1) of each isolate spread on Striga plants 70 DAS.
The following observations were made
| • | Percentage of dead Striga plants 14, 21 and 28 Days after Inoculation (DAI) of spores |
| • | Counts of emerged Striga plants flowering or bearing seed capsules |
| • | A visual estimation of Striga vigor recorded 14, 21 and 28 DAI using a rating scale of 0 to 9 (Haussmann et al., 2000), where 0= No emerged Striga plant, 9 = Mean height of Striga plants > 40 cm, with more than 10 branches |
| • | Weight of Striga dry biomass at sorghum harvest |
| • | Weight of sorghum dry biomass (stalks and roots) and grain yield |
Statistical analysis Percent germination of Striga and percentage of dead Striga were arc sine transformed (Gomez and Gomez, 1984) before performing ANOVA (SAS Institute. Cary. NC) followed by separation of means using the LSD’s test. Number of flowering Striga, vigor of Striga plants, Striga dry biomass and sorghum yields were subjected to an Analysis of Variance (ANOVA) and to separation of means using LSD’s test.
RESULTS
Effect of Fusarium spores on Striga germination: Of the 51 isolates evaluated, 14 were found to reduce significantly Striga germination in comparison to the control. Indeed, the 14 isolates led to a reduction by more than 50% of potential germination rate of Striga (Fig. 1). The spores of F. oxysporum (34-Fo), F. equiseti (5-Kou, 32-Or, 31-Kom, 13-Ba) and Fusarium sp. (6-Fa, 125b-Za, 150a-M) were more effective reducing Striga germination by 78 to 96%.
Effect of Fusarium metabolites: ANOVA revealed that the 4 doses (1, 10, 33 and 67 mg mL-1) of metabolites of the 14 isolates significantly reduced Striga germination. The dose of 1 mg metabolites mL-1 of the 14 isolates slightly influenced Striga germination. A reduction by more than 50 % was recorded with 10 mg metabolites mL-1 of the isolates F. oxysporum 34-Fo, F. equiseti (5-Kou, 6-Fa) and Fusarium sp. 125b-Za. Striga germination rate was 0% with four isolates at the dose of 33 mg metabolite mL-1 whereas at the dose of 67 mg mL-1, Striga germination was inhibited with seven isolates (Fig. 2).
The results showed a highly significant positive correlation (r = 0.88) between Striga germination rates for seeds treated with Fusarium spores and Striga germination rates when seeds were treated with Fusarium metabolites.
Effect of Fusarium spores on Striga infestation: Striga infestation of sorghum was significantly reduced 14, 21 and 28 Days after Inoculation (DAI) of the emerged parasite with Fusarium spores (Table 1). Striga death rates greater than 11% were recorded with four isolates 14 DAI and isolate 34-Fo was the most effective with a 22% kill rate. A Striga death rate greater than 11% was recorded with 8 isolates 21 and 28 DAI; isolates of F. oxysporum 34-Fo, F. equiseti (5-Kou, 32-Or) and Fusarium sp. (6-Fa, 150a-M) were the most effective for both dates (Striga death rate varying from 17 to 37%). Number of flowering Striga and Striga bearing seed capsules were significantly reduced 21 and 28 DAI with Fusarium spores (Table 2).
| Table 1: | Effect of inoculation with Fusarium spores on Striga death rate in greenhouse |
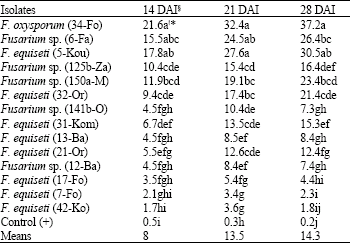 | |
| §: Days after inoculation, !: Means are back-transformations of percentage of dead Striga, *: Means within a column followed by the same letter are not significantly different (p = 0.01) according to LSD’s test | |
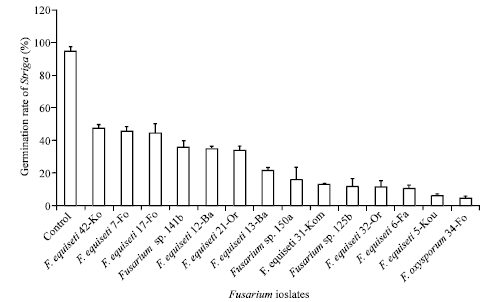 | |
| Fig. 1: | Effect of spores of Fusarium isolates on Striga hermonthica seeds germination. Vertical lines indicate standard errors of the means. Control: Striga seeds conditioned in sterile distilled water |
Seven and 10 isolates significantly reduced number of flowering Striga and Striga bearing fruits, respectively, 21 and 28 DAI.
Effect of Fusarium spores on Striga growth: Fusarium isolates significantly reduced Striga vigor 21 and 28 DAI (Table 2). Striga vigor was highly reduced 21 DAI with three isolates whereas at 28 DAI, all the isolates (except F. equiseti 42-Ko) reduced Striga vigor. The most effective were isolates of F. oxysporum 34-Fo and Fusarium sp. 6-Fa. The fourteen Fusarium isolates significantly reduced Striga biomass compared to the control and thirteen led a reduction by more than 48%. F. oxysporum 34-Fo was the most efficient at reducing Striga growth (84% biomass reduction) followed by Fusarium sp 6-Fa (78% biomass reduction).
Effect of Fusarium spores on Striga infestation and sorghum yield: Application of Fusarium spores significantly reduced number of emerged Striga plants in pots. As a result, the dry biomass of sorghum stalks and roots and sorghum grain yield were improved. Striga infestation only reduced sorghum root biomass on the control. Seven fungal isolates (F. oxysporum 34-Fo, F. equiseti (5-Kou, 31-Kom, 32-Or) and Fusarium sp. (150a-M, 6-Fa, 125b-Za)) all alleviated Striga effect with respect to stalk biomass (Fig. 3).
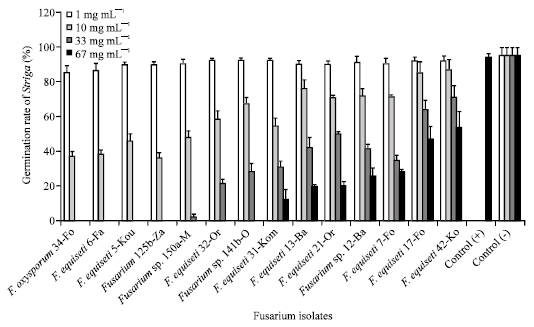 | |
| Fig. 2: | Effect of metabolites of Fusarium isolates on Striga hermonthica seeds germination. Vertical lines indicate standard errors of the means. Control (-): Striga seeds conditioned with sterile distilled water; Control (+): seeds conditioned with 67 mg mL-1 of growth medium filtrate freeze-dried |
| Table 2: | Effect of Fusarium spores on number of flowering Striga, Striga vigor and dry biomass in greenhouse |
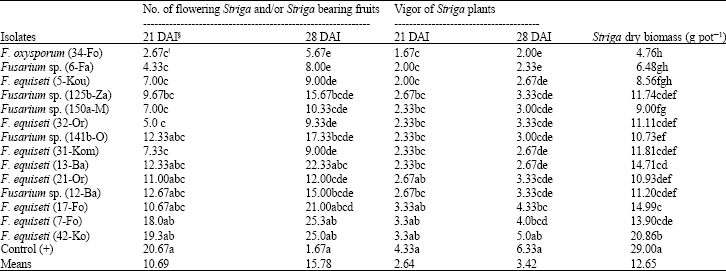 | |
| §: Days after inoculation. !: Means within a column followed by the same letter are not significantly different (p = 0.05) according to LSD’s | |
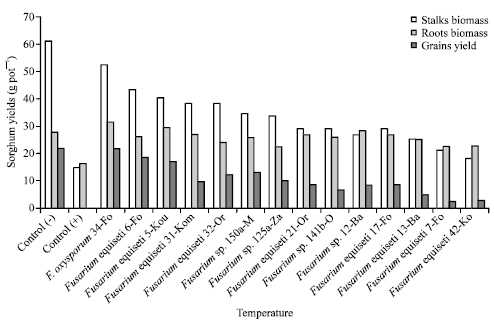 | |
| Fig. 3: | Sorghum stalks, roots dry biomass, Sorghum grains yield following upon application of Fusarium spores on Striga hermonthica plants. Vertical lines indicate standard errors of the means. Control (+): Sorghum with Striga, Control (-): Sorghum without Striga |
Striga depressive effect on sorghum grain yield was alleviated by three isolates (F. oxysporum 34-Fo, F. equiseti 5-Kou and Fusarium sp. 6-Fa), for which we recorded, respectively, 78, 84% and 99% of yield of the control without Striga. Sorghum grain yield recorded with the other isolates were between 12% and 60% of that of the control (Fig. 3).
DISCUSSION
The results of these investigations confirm the potential pathogenicity of Fusarium sp. on S. hermonthica as reported by Kroschel et al. (1996), Abbasher et al. (1998), Elzein and Kroschel (2004) and Elzein et al. (2009). However, the pathogenicity varied significantly between Fusarium isolates collected in the same location. In addition, bioherbicidal activity of Fusarium isolates from different locations should be influenced by the method of inoculation with Fusarium (Kroschel et al., 1996; Marley et al., 1999).
Spores of 51 fungi isolates were evaluated for inhibition of Striga seed germination. Of these 51 isolates, 14 were found to reduce Striga germination by more than 50%. Isolates of F. equiseti (13-Ba, 150a-M, 31-Kom, 32-Or, 5-Kou), Fusarium sp. (125b-Za, 6-Fa) and F. oxysporum 34-Fo were the most effective with a reduction of Striga germination by more than 78%. Kroschel et al. (1996) also demonstrated that spores of two F. oxysporum isolates (Fo1, Fo2) and one F. solani isolate (Fs1) used to condition Striga seeds reduced Striga germination by more than 90%. A suspension of 10 mL F. nygamai at a rate of 4x106 spore mL-1 used to inoculate Striga seeds in roots chamber reduced germination rate by 88% (Abbasher and Sauerborn, 1992). Reduction of Striga germination by spores of Fusarium isolates is probably the result of penetration of mycelium into Striga seeds which are then killed (Sauerborn et al., 1996).
Application of fungal metabolites to parasite seed reduced or inhibited Striga germination. In this study, no germination of Striga was observed with 33 mg mL-1 metabolites of isolates Fusarium sp. (125b-Za, 6-Fa), F. equiseti 5-Kou and F. oxysporum 34-Fo. Three other isolates (Fusarium sp. (141b-O, 150a-M) and F. equiseti 32-Or) showed a complete inhibition of Striga germination with 67 mg mL-1 metabolites. The bio-herbicide effect of these seven isolates is probably due to metabolites of the filtrates. Four toxic metabolites (fusaric acid, 9,10-acid dehydrofusaric, 2 ester-methyl) produced in vitro by F. nygamai and completely inhibiting Striga germination were identified (Zonno et al., 1996; Idris et al., 2003). Four other metabolites of tricothecene group (acuminatin, neosolaniol, 8-acetylneosolaniol and tetraacetoxy T-2 tetraol (neosolaniol diacetate)) produced by F. solani (Sud 96) lead to inhibition of S. hermonthica seed germination (Sugimoto et al., 2002). The high positive correlation between Striga germination rates with Fusarium spores and metabolites indicates that when evaluating Striga germination rate with Fusarium spores, it appears unnecessary to determine Striga germination rates with Fusarium metabolites.
Application of Fusarium spores led to a Striga death rate more than 20% with five isolates. In this study, the highest Striga death rate due to application of Fusarium spores was lower than 40% whereas Striga death rates of 44 to 89% were reported with local isolates from Nigeria (Marley et al., 1999). Fusarium isolates affected Striga vigor and consequently Striga biomass was reduced by more than 48% with all isolates except F. equiseti 42-Ko. The great reduction of number of flowering and fruiting Striga recorded with 10 Fusarium isolates 28 DAI, may lead to reduce Striga seeds production. Thus, the application of Fusarium spores on Striga in the field could limit the increase of the soil seed bank. The increase of sorghum dry biomass and yield was the result of effectiveness of Fusarium isolates to protect the host plants.
We are currently investigating the metabolites/toxins responsible for Striga death. A potential use of Fusarium isolates by farmers to control Striga infestations in the field could be through crop seed coating with fungal spores before sowing. So, use of Fusarium isolates could be a biological component in the integrated Striga management in West Africa.
ACKNOWLEDGMENTS
We are grateful to International Foundation of Science (IFS) and INTSORMIL which funded this study. We also thank the Institut de l’Environnement et des Recherches Agricoles (INERA) of Burkina Faso for the infrastructure allowing us to carry out this study.
REFERENCES
- Abbasher, A.A., D.E. Hess and J. Sauerbom, 1998. Fungal pathogens for biological control of Striga hermonthica or Sorghum and pearl millet in West Africa. Afr. Crop Sci. J., 6: 179-188.
Direct Link - Berner, D.K. and O.A. Williams, 1998. Germination of Striga gesnerioides seeds stimulated by hosts and non-hosts. Plant Dis., 82: 1242-1247.
Direct Link - Ciotola, M., A.K. Watson and S.G. Hallet, 1995. Discovery of an isolate of Fusarium oxysporum with potential to control Striga hermonthica in Africa. Weed Res., 35: 303-309.
CrossRefDirect Link - Ejeta, G. and L.G. Butler, 1993. Host-parasite interactions throughout the Striga life cycle and their contributions to Striga resistance. Afr. Crop Sci. J., 1: 75-80.
CrossRefDirect Link - Elzein, A. and J. Kroschel, 2004. Fusarium oxysporum Foxy 2 shows potential to control both Striga hermonthica and S. Asiatica. Weed Res., 44: 433-438.
CrossRefDirect Link - Elzein, A., J. Kroschel, P. Marley and G. Cadisch, 2009. Does vacuum-packaging atmosphere or co-delivered amendments enhance shelf-life of Striga-mycoherbicidal products containing Fusarium oxysporum f.sp. strigae during storage? Biocontrol Sci. Technol., 119: 349-367.
Direct Link - Gressel, J., A. Hanafi, G. Head, W. Marasas and A.B. Obilana et al., 2004. Major heretofore intractable biotic constraints to african food security that may be amenable to novel biotechnological solutions. Crop Prot., 23: 661-689.
CrossRefDirect Link - Haussmann, B.I.G., D.E. Hess, H.G. Welz and H.H. Geiger, 2000. Improved methodologies for breeding Striga-resistant sorghums. Field Crops Res., 66: 195-211.
CrossRef - Hess, D.E., G. Ejeta and L.G. Butler, 1992. Selecting sorghum genotypes expressing a quantitative biosynthetic trait that confers resistance to Striga. Phytochemistry, 31: 493-497.
CrossRefDirect Link - Idris, A.E., M.A. Abouzeid, A. Boari, M. Vurro and A. Evidente, 2003. Identification of phytotoxic metabolites of a new Fusarium sp. Inhibiting germination of Striga hermonthica seeds. Phytopathol. Med., 42: 65-70.
Direct Link - Kroschel, J., A. Hundt, A.A. Abbasher and J. Sauerborn, 1996. Pathogenicity of fungi collected in northern Ghana to Striga hermonthica. Weed Res., 36: 515-520.
CrossRefDirect Link - Marley, P.S., S.M. Ahmed, J.A. Shebayan and S.T.O Lagoke, 1999. Isolation of Fusarium oxysporum with potential for biocontrol of the witchweed Striga hermonthica in the Nigerian Savanna. Biocontrol. Sci. Technol., 9: 159-163.
CrossRefDirect Link - Sauerborn, J., I. Dorr, A.A. Abbasher, H. Thomas and J. Kroschel, 1996. Electron microscopic analysis of the penetration process of Fusarium nygamai, a hyperparasite of Striga hermonthica. Biol. Control, 7: 53-59.
Direct Link - Schaub, B., P. Marley, A. Elzein and J. Kroschel, 2006. Field evaluation of an integrated Striga hermontica management in Sub-Saharan Africa: Synergy between Striga-mycoherbicides (biocontrol) and sorghum and maize resistant varieties. J. Plant Dis. Prot., 20: 691-699.
Direct Link - Siame, B.A., Y. Weerasuriya, K. Wood, G. Ejeta and G.L. Butler, 1993. Isolation of Strigol, a germination stimulant for Striga asiatica, from host plants. J. Agric. Food Chem., 41: 1486-1491.
CrossRefDirect Link - Sugimoto, Y., N.E. Ahmed, N. Yasuda and S. Inanaga, 2002. Trichothecene inhibitors of Striga hermonthica germination produced by Fusarium solani. Weed Sci., 50: 658-661.
Direct Link - Vogler, R.K., G. Ejeta and L.G. Butler, 1996. Inheritance of low Production of Striga germination stimulant in sorghum. Crop Sci., 36: 1185-1191.
Direct Link - Zonno, M.C., M. Vurro, R. Capasso, A. Evidente, A. Cutignano, J. Sauerborn and H. Thomas, 1996. Phytotoxin metabolites produce by Fusarium nygamai from Striga hermonthica. Proceedings of the 9th International Symposium on Biological Control of Weeds, Jan. 19-26, University of Cape Town, South Africa, pp: 223-226.