
B. Yousefi
Department of Anatomical Sciences, Semnan University of Medical Sciences, Semnan, Iran
H. R. Samene
Department of Anatomical Sciences, Semnan University of Medical Sciences, Semnan, Iran
Pakistan Journal of Biological Sciences
Year: 2010 | Volume: 13 | Issue: 4 | Page No.: 185-189







ABSTRACT
This study was planned to determine the effects of All-Trans Retinoic Acid (ATRA) on the progenitors of White Blood Cells (WBC) and survey their outcomes in rat's embryo during both late-yolk sac and fetal liver stages of hematopoiesis. Single oral dose (100 mg kg-1) of ATRA was administered to rat on Gestation Day (GD) 10 and fetuses were observed on GD 18. The fetus's blood (from experimental group and control each, n = 24) were obtained directly from heart, as placental and mother circulation was continued and subsequently processed for Giemsa staining and followed by WBC counting and measuring. Statistical analysis was made by student t-test. A p-value<0.05 was considered significant. In the experimental embryos on GD 18, the mean number of WBC (29.2%), neutrophil, lymphocyte and monocyte were increased. There was a significant difference in WBC (p<0.0001) and neutrophil (p<0.001) between the groups in this regard. The mean diameter of neutrophil, lymphocyte and monocyte were compared in two groups. The results showed no significant change on experimental and control groups. We concluded that ATRA may have positive effects on proliferation, differentiation and maturation of neutrophil without having any significant effects on the diameter of cells throughout normal granulocyte differentiation in embryo during both late-yolk sac and fetal liver stages of hematopoiesis.
PDF Abstract XML References Citation
How to cite this article
B. Yousefi and H. R. Samene, 2010. The Effects of All-Trans Retinoic Acid on Leukocytes in Rat's Embryo. Pakistan Journal of Biological Sciences, 13: 185-189.
DOI: 10.3923/pjbs.2010.185.189
URL: https://scialert.net/abstract/?doi=pjbs.2010.185.189
DOI: 10.3923/pjbs.2010.185.189
URL: https://scialert.net/abstract/?doi=pjbs.2010.185.189
INTRODUCTION
The commitment of embryonic cells to hematopoietic fates begins in proximal region of egg cylinder at the mid-primitive streak stage with the simultaneous appearance of primitive erythroid and macrophage progenitors (Palis et al., 1999). First, the differentiation of leukocytes occurs and followed by erythrocytes differentiation (Tashiro et al., 2006). The first leukocytes that arise in the development of vertebrate embryos are the primitive macrophages, which differentiate in the yolk sac and then, quickly invade embryonic tissues (Le Guyader et al., 2008). It is apparent that a wide variety of external and internal stimuli influence and modulate the lineage choice and differentiation during hematopoiesis (Bedi and Sharkis, 1995). Hematopoiesis is a highly regulated process resulting in the formation of all blood lineages (Buitenhuis et al., 2008).
Retinoids are a group of natural and synthetic vitamin A analogues and exert important effects on the growth and differentiation of various cell types including hematopoietic progenitors (Douer and Koeffler, 1982). Also, there is a strong body of evidence that vitamin A (Kang et al., 2007) and retinoids have immune regulatory functions (Kang et al., 2007; Chen et al., 2009) and improve the numbers of various immune cell types (T helper 2) (Iwata et al., 2003). The fact that Retinoic Acid (RA) is a differentiating agent for granulocyte in culture has been well established for both normal and leukemic cells (Lawson and Berliner, 1999). However, the effects of exogenesis All-Trans Retinoic Acid (ATRA) throughout normal granulocyte differentiation during both late-yolk sac and fetal liver stages of hematopoiesis have so far not been investigated. Therefore, in this study, we have examined the effects of ATRA on the progenitors of White Blood Cells (WBC) and survey their outcomes.
MATERIALS AND METHODS
Animals: In this study, Wistar strain of rats (about 3 months of age) were used. They were housed in light (12:12 light: dark cycle) and temperature-controlled (21°C) rooms and maintained on laboratory chow and tap water provided ad libitum. Adult virgin females (n = 10) (180-200 g in weight) were mated overnight with males of the same stock. A vaginal plug and smear observed indicated day 0 of pregnancy. The experimental protocol was approved by the Ethics Review Committee for Animal Experimental of the Semnan University of Medical Sciences. This study was conducted from 2007 to 2009.
Drug: ATRA (Sigma-Aldrich, USA) were used in this study.
Preparation of solution: Single dose of 100 mg kg-1 of ATRA (Sigma-Aldrich, USA) suspended in alcohol; corn oil (1:9) mixture (Padmanabhan, 1998) (light-proof containers under yellow light) was administered. The drug was given by oral incubations on the morning of Gestation Day (GD), 10 to experimental group (n = 5). The control (n = 5) were vehicle treated. The animals were euthanized on GD 18 by ether.
Samples preparation: The fetus’s blood (experimental and control each, n = 24) were obtained directly from heart, as placental and mother circulation was continued. Blood cytology slides were made using the cytospin preparation (Akbari et al., 2007) and subsequently processed for staining and followed by counting and measuring. These measurements in each sample were done in ten slides and in the five areas for each section (using Randomization Implementation). The embryos weighed and fixed in 10% formalin, examined for external malformation. The blood cells were studied by light microscope and eye piece (x200 for WBC count and x1000 for diameter).
Statistical analysis: The values were expressed as Mean±SEM. Comparison between two groups was made by student t-test (Mohammadi et al., 2009), p<0.05 was considered as significant different.
RESULTS
The experimental group showed a marked malformation on external inspection. In the experimental embryos on GD 18, the number of WBC per field was 42.12±2.14 with 29.3% increase in this group, compared to control group (32.58±0.88) as was shown in Fig. 1. In the experimental group, the number of neutrophil per field was 12.8±1.50 with 85% increase in this group, compared to control group (6.9±0.30) as was shown in Fig. 1. In the experimental embryos, the number of lymphocyte per field was 20.6±1.32 with 16.4% increase as compared to control group (17.75±1.15), no significant difference was found between two groups. The number of monocyte per field in experimental group was 8.6±0.56 with 8.4% increase when compared to control group (7.91±0.24).There were significant relationships between two groups in WBC (p<0.0001) and neutrophil (p<0.001) counts. However, there were no significant changes in monocyte and lymphocyte numbers in two groups Fig. 1. The mean diameter of neutrophil, lymphocyte, monocyte, for experimental and control groups were: 6.88±0.18 and 7.26±0.13; 5.8±0.13 and 5.6±0.15; 8.35±0.24 and 8.18± 0.18, respectively (Fig. 2).
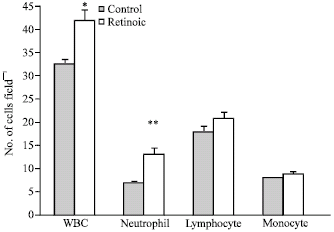 | |
| Fig. 1: | Effect of ATRA (100 mg kg-1) on WBC, neutrophil, lymphocyte and monocyte numbers of rat embryo on GD 18. *p< 0.0001, **p< 0.001 relative to values in experimental group. Results was presented as Mean±SEM for n = 24 embryo per group |
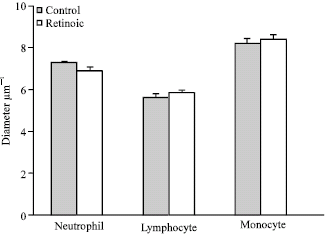 | |
| Fig. 2: | Effect of ATRA (100 mg kg-1) on the mean diameter of WBC in rat embryo on GD 18, in comparison with control group. No significant differences between groups were noted in this regard |
These data showed that the mean diameters of leukocytes were not markedly changed.
DISCUSSION
ATRA and diverse synthetic analogues affect differentiation of neoplastic and normal hematopoietic cells (Lanotte et al., 1991). In this study, a single dose of ATRA has been used to identify its effects during hematopoietic differentiation in rat’s embryos. The data indicated that ATRA enhances the proliferation and differentiation of myeloid compartment, especially neutrophils.
The increase in the neutrophils population, was seen here, shows that exogenous ATRA is able to improve the proliferation and differentiation of their progenitors during both late -yolk sac and fetal liver stages of hematopoiesis in embryo. These results are in line with those of Leelasiri et al. (2005), which reported an increase in neutrophil number after ATRA treatment, as well as with a reports showing induced granulocytic differentiation of acute myeloblastic, promyelocytic leukemia cell lines (Valtieri et al., 1991) and acute promyelocytic leukemia blasts in vivo (Warrell et al., 1991). In addition, this finding is consistent with results from previous studies showing that ATRA enhances the Hematopoietic Progenitor Cells (HPCs) in vitro (Collins et al., 2001; Tobler et al., 1986). In the hematopietic system, RARα is expressed at significant level in granulocyte (Labrecque et al., 1998) and stimulates differentiation in response to exogenous RA. This receptor can bidirectionally modulate granulopoiesis as a differentiation factor when liganded to RA or as an inhibitor in the absence of ligand (Kastner et al., 2001). Our data about lymphocyte and monocyte were in line with the results of Seguin-Devaux et al. (2005), who determined that RA directly enhances the number of T lymphocytes in the peripheral blood. Although these enhancing effects were not significant. It was contradicted with previously reports showing that RA decreases proliferation of lymphocyte (Stosic-Grujicic and Ejdus, 1994) and human myeloid leukemia cells in vitro (Douer and Koeffler, 1982).
Macrophage progenitors (Mac-CFC) increase in numbers in the yolk sac until GD 9.5 and are preferentially localized to the tail and para aortic splanchopleura aorta-gonad-mesonephros region prior to the development of the liver. These progenitors increase within the fetal liver between GD10.5 to 11.5. GM-CFC could also be detected within the yolk sac on GD 8.5. Low numbers of them persisted in both the yolk sac and embryo proper until GD 11.5, at which time that significant numbers were present in both blood and fetal liver. These progenitors gave rise to small to medium size colonies that contain cells with both neutrophils and macrophage morphology (Palis et al., 1999). It should be noted that when ATRA was given, the progenitors of leukocyte, certainly were Mac-CFC and bipotential granulocyte/macrophage (GM-CFC) in the embryo.
The apparent difference in the responses of neutrophil, lymphocyte and monocyte to ATRA may be as under: It is hypothesized that the inhibitory effects of RA on erythroid, monocyte and high-proliferative potential colony-forming cells originated through activation of either RAR/RXR of D3R/D3R-D3R/RXR, while, the stimulatory activity on granulocytic colony formation could theoretically be described only to activation of RAR/RXR (Tocci et al., 1996). The exogenous RA is able to redirect erythroid-monocyte, or eosinophil-committed progenitors toward a granulocyte fate (Tobler et al., 1986; Tocci et al., 1996; Paul et al., 1995). In addition, it possibly induces an HPC shift from erythroid to granulocytic neutrophilic-differentiation pathway (Labbaye et al., 1994). It also appears to orient the differentiation of pluripotent hematopietic progenitors toward the granulocyte lineage (Tocci et al., 1996). Thus, it may be involved as a commitment factor (Paul et al., 1995) and fate specification for this lineage (Kastner et al., 2001; Seguin-Devaux et al., 2005) with different effective concentrations in vitro and in vivo.
After data analysis, we found that ATRA not to be of significant influence on cell diameter. These finding are in contrast with those of Kinoshita et al. (2000), who reported a decrease in mast cell’s diameter with this agent. The size of a cell depends on intrinsic and extrinsic factors and is regulated by Ribosome biogenesis in multicellular organisms (Hafen and Stocker, 2003). That is also a consequence of correlation between the rates of cell-cycle progression and growth, but four different general processes modulate the two rates (Rupes, 2002). Although, the mechanisms responsible for the differences in cell size among different red cell lineages are not well understood, they may relate to differential rates of cell division (Kingsley et al., 2004). This issue needs future investigations.
CONCLUSION
This study shows that administration of exogenous ATRA during both late-yolk sac and fetal liver stages of hematopoiesis may be improve proliferation, differentiation and maturation of neutrophils, without having any deleterious effects on the dimenation of WBC.
ACKNOWLEDGMENT
This study was supported by grant from Semnan University of Medical Sciences and Health Services.
REFERENCES
- Mozdarani, H., S. Akhlaghpoor, A.A. Pourfatollah, M. Soleimani and A.A.M. Akbari, 2007. Evaluation of the homing of human CD34+ cells in mouse bone marrow using clinical MR imaging. Pak. J. Biol. Sci., 10: 833-842.
CrossRefPubMedDirect Link - Bedi, A. and S.J. Sharkis, 1995. Mechanisms of cell commitment in myeloid cell differentiation. Curr. Opin. Hematol., 2: 12-21.
PubMed - Chen, X., R.S. Welner and P.W. Kincade, 2009. A possible contribution of retinoids to regulation of fetal B lymphopoiesis. Eur. J. Immunol., 39: 2515-2524.
PubMed - Collins, S.J., J. Ulmer, L.E. Purton and G. Darlington, 2001. Multipotent hematopoietic cell lines derived from C/EBPalpha(-/-) knockout mice display granulocyte macrophagus-colony-stimulating factor, granulocyte-colony-stimulating factor and retinoic acid-induced granulocytic differentiation. Blood, 98: 2382-2388.
PubMed - Douer, D. and H.P. Koeffler, 1982. Retinoic acid enhances growth of human early erythroid progenitor cells in vitro. J. Clin. Invest., 69: 1039-1041.
PubMed - Hafen, E. and H. Stocker, 2003. How are the sizes of cells, organs and bodies controlled?. PLoS. Biology., 1: E86-E86.
PubMed - Iwata, M., Y. Eshima and H. Kagechika, 2003. Retinoic acids exert direct effects on T cells to suppress Th1 development and enhance Th2 development via retinoic acid receptors. Int. Immunol., 15: 1017-1025.
CrossRefPubMedDirect Link - Kang, S.G., H.W. Lim, O.M. Andrisani, H.E. Broxmeyer and C.H. Kim, 2007. Vitamin A metabolites induce gut-homing FoxP3+ regulatory T cells. J. Immunol., 179: 3724-3733.
PubMedDirect Link - Kastner, P., H.J. Lawrence, C. Waltzinger, N.B. Ghyselinck, P. Chambon and S. Chan, 2001. Positive and negative regulation of granulopoiesis by endogenous RARalpha. Blood, 97: 1314-1320.
PubMed - Kingsley, P.D., J. Malik, K.A. Fantauzzo and J. Palis, 2004. Yolk sac-derived primitive erythroblasts enucleate during mammalian embryogenesis. Blood, 104: 19-25.
PubMed - Kinoshita, T., K. Koike, H.H. Mwamtemi, S. Ito and S. Ishida et al., 2000. Retinoic acid is a negative regulator for the differentiation of cord blood-derived human mast cell progenitors. Blood, 95: 2821-2828.
PubMedDirect Link - Labbaye, C., M. Valtieri, U. Testa, A. Giampaolo and E. Meccia et al., 1994. Retinoic acid downmodulates erythroid differentiation and GATA1 expression in purified adult-progenitor culture. Blood, 83: 651-656.
PubMedDirect Link - Labrecque, J., D. Allan, P. Chambon, N.N. Iscove, D. Lohnes and T. Hoang, 1998. Impaired granulocytic differentiation in vitro in hematopoietic cells lacking retinoic acid receptors alpha1 and gamma. Blood, 92: 607-615.
PubMed - Lanotte, M., V. Martin-Thouvenin, S. Najman, P. Balerini, F. Valensi and R. Berger, 1991. NB4, a maturation inducible cell line with t (15; 17) marker isolated from a human acute promyelocytic leukemia (M3). Blood, 77: 1080-1086.
PubMed - Lawson, N.D. and N. Berliner, 1999. Neutophil maturation and the role of retinoic acid. Exp. Hematol., 27: 1355-1367.
PubMed - Le-Guyader, D., M.J. Redd, E.Colucci-Guyon, E. Murayama and K. Kissa et al., 2008. Origins and unconventional behavior of neutrophils in developing zebrafish. Blood, 111: 132-141.
PubMed - Leelasiri, A., T. Numbenjapol, W. Prayoonwiwat, W. Mongkolsritrakul and C. Srisawat, 2005. Successful treatment of retinoic acid syndrome with dexamethasone: A case report. J. Med. Assoc. Thai., 88: 302-310.
PubMed - Mohammadi, S.S., E. Shahbazian, G. Shoeibi and F. Almassi, 2009. Effect of scalp infiltration with bupivacaine on early hemodynamic responses during craniotomy under general anesthesia. Pak. J. Biol. Sci., 12: 603-606.
CrossRefPubMedDirect Link - Padmanabhan, R., 1998. Retinoic acid-induced caudal regression syndrome in the mouse fetus. Reprod. Toxicol., 12: 139-151.
PubMed - Palis, J., S. Robertson, M. Kennedy, C. Wall and G. Keller, 1999. Development of erythroid and myeloid progenitors in the yolk sac and embryo proper of the mouse. Development, 126: 5073-5084.
PubMed - Paul, C.C., S. Mahrer, M. Tolbert, B.L. Elbert and I. Wong et al., 1995. Changing the differentiation program of hematopoietic cells: Retinoic acid-induced shift of eosinophil-committed cells to neutrophils. Blood, 86: 3737-3744.
PubMed - Seguin-Devaux, C., D. Hanriot, M. Dailloux, V. Latger-Cannard and F. Zannad et al., 2005. Retinoic acid amplifies the host immune response to LPS through increased T lymphocytes number and LPS binding protein expression. Mol. Cell. Endocrinol., 245: 67-76.
PubMed - Stosic-Grujicic, S. and L. Ejdus, 1994. Modulation of in vitro T cell alloreactivity by synthetic retinoids. Immunopharmacology, 27: 87-92.
PubMed - Tashiro, S., A. Sedohara, M. Asashima, Y. Izutsu and M. Maeno, 2006. Characterization of myeloid cells derived from the anterior ventral mesoderm in the Xenopus laevis embryo. Dev. Growth. Differ., 48: 499-512.
PubMed - Tobler, A., M.I. Dawson and H.P. Koeffler, 1986. Retinoids. Structure-function relationship in normal and leukemic hematopoiesis in vitro. J. Clin. Invest., 78: 303-309.
PubMed - Tocci, A., I. Parolini, M. Gabbianelli, U. Testa and L. Luchetti et al., 1996. Dual action of retinoic acid on human embryonic/fetal hematopoiesis: Blockade of primitive progenitor proliferation and shift from multipotent/erythroid/monocytic to granulocytic differentiation program. Blood, 88: 2878-2888.
PubMed - Valtieri, M., G. Boccoli, U. Testa, C. Barletta and C. Peschle, 1991. Two-step differentiation of AML193 leukemia line: Terminal maturation is induced by positive interaction of retinoic acid with granulocyte colony-stimulating factor (CSF) and vitamin D3 with monocyte CSF. Blood, 77: 1804-1812.
PubMed - Warrell, R.P., S.R.J. Frankel, W.H.J. Miller, D.A. Scheinberg and L.M. Itri et al., 1991. Differentiation therapy of acute promyelocytic leukemia with tretinoin (all-trans-retinoic acid). N. Engl. J. Med., 324: 1385-1393.
PubMedDirect Link - Buitenhuis, M., L.P. Verhagen, H.W. van Deutekom, A. Castor and S. Verploegen et al., 2008. Protein kinase B (C-akt) regulates hematopoietic lineage choice decisions during myelopoiesis. Blood, 111: 112-121.
PubMed