
Y. Ameyapoh
Laboratoire de Microbiologie et de Contr�le de Qualite des Denrees Alimentaires, ESTBA-UL, Universite de Lome, Togo
Jean-Yves Leveau
Laboratoire de Microbiologie Industrielle, ENSIA, France
Simplice D. Karou
Laboratoire de Microbiologie et de Contr�le de Qualite des Denrees Alimentaires, ESTBA-UL, Universite de Lome, Togo
M. Bouix
Laboratoire de Microbiologie Industrielle, ENSIA, France
Seyram K. Sossou
Laboratoire de Microbiologie et de Contr�le de Qualite des Denrees Alimentaires, ESTBA-UL, Universite de Lome, Togo
C. De Souza
Laboratoire de Microbiologie et de Contr�le de Qualite des Denrees Alimentaires, ESTBA-UL, Universite de Lome, Togo
Pakistan Journal of Biological Sciences
Year: 2010 | Volume: 13 | Issue: 3 | Page No.: 132-137







ABSTRACT
The present study aimed to access for the physiochemical parameters of vinegar production through Togolese local variety Mangovi of mango Mangifera indica juice fermentation. The juice was fermented successively by Saccharomyces cerevisisae and acetic bacteria. The levels of ethanol and acetic acid in the juice during the production of vinegar were monitored by gas chromatography and titrimetry methods, respectively. The physiological state of the yeast Saccharomyces cerevisiae L2056 was determined by flow cytometry using a dual fluorescent labeling of diacetate carboxy-fluorescein (CFDA) and propidium iodide. The results indicated that 200 mL of mango juice, sugar content 20 Brix, set in alcoholic fermentation with 106 yeast cells produced 22.4 g L-1 ethanol in 72 h. Acetic fermentation transformed 93% of this ethanol to acetic acid in 288 h. Twenty-four hours after the beginning of alcoholic fermentation, 91% of cells were viable, 8.85% were stressed and 0.05% died. After 24 h of acetic fermentation, viable, stressed and dead cells were 45, 12 and 39%, respectively; corresponding to the passage of acetic vinegar level from 0.9 to 2.1°. At the end of the acetic fermentation, dead cells were estimated to 98% at and acetic acid to 4.7°. Using consecutive fermentations is suitable technique for vinegar production from mango juice. The application of the present results may contribute to avoid fruits post harvest losses.
PDF Abstract XML References Citation
How to cite this article
Y. Ameyapoh, Jean-Yves Leveau, Simplice D. Karou, M. Bouix, Seyram K. Sossou and C. De Souza, 2010. Vinegar Production from Togolese Local Variety Mangovi of Mango Mangifera indica Linn. (Anacardiaceae). Pakistan Journal of Biological Sciences, 13: 132-137.
DOI: 10.3923/pjbs.2010.132.137
URL: https://scialert.net/abstract/?doi=pjbs.2010.132.137
DOI: 10.3923/pjbs.2010.132.137
URL: https://scialert.net/abstract/?doi=pjbs.2010.132.137
INTRODUCTION
Post-harvest losses of seasonal fruits are considerable in tropical countries. In Togo, according to the ministry of agriculture, mango losses can be estimated to 50%. Mangoes are particularly biodegradable and highly perishable fruits, because of their high water content and the presence of carbohydrates. The sugars adhering to the mango fruit are ideal for fermentations. Several reports suggested the possibilities of alcohol and vinegar production from juice (Ethiraj and Suresh, 1990). The reduction of overproduction fruits of the local variety Mangovi in Togo, Mangifera indica by alcohol production resulted in high content of residual sugar at the end of the fermentation. Some reports demonstrated that, the production of vinegar needs some sugar in the juice and ethanol contents is not a limiting factor for the growth of acetic bacteria involved in this production (Gullo et al., 2005).
Acetic bacteria are of important industrial interest as well as lactic bacteria and yeast. Since, the acetic bacteria are involved in the production or spoilage of food, their species identification is lead information for the technologist trying to control a bioprocess in industry (Trcek, 2005). The production of traditional balsamic vinegar uses a selection of yeasts and acetic bacteria (De Vero et al., 2006). According to Ndoye et al. (2006) acetic acid bacteria are Gram negative, strictly aerobic and commonly found in nature on different plants (fruits, grains, herbs etc.). They have the ability to oxidize the different types of alcohol in major biotechnological process food products such as vinegar (Trcek, 2005). In recent years, there have been major advances in understanding their taxonomy, molecular biology and physiology and in methods for their isolation and identification (Raspor and Goranoviccaron, 2008). However, problems related to environmental conditions such as temperature variations and process technology limited the industrial applications in tropical regions (Ndoye, 2007). The successive fermentations: alcoholic and acetic fermentation during the production of vinegar could lead to change physiological status of yeast. Flow cytometry allows the following up of the physiological state of the yeast during the production and control in bioreactors (Kacmar et al., 2006). Kacmar et al. (2004) reported that cytometric system indicates not only the quantity of yeast cells and their viability but also provides a reproducible detail on culture dynamics. Thus, increasing the acetic acid content of the wort from the ethanol oxidation would influence the viability, stress and disintegration of yeast.
Our work therefore, aimed access for the microbiological and physico-chemical parameters of vinegar production.
MATERIALS AND METHODS
Plant material: The mango Mangifera indica (L.) Merr. (Bromeliaceae), local variety mangovi was used in this study. Mango fruits, average weight between 120 and 165 g, were purchased from Ahanoukope market, in Lomé town, in Togo, in April 2006.
Microorganisms: The yeast strain Saccharomyces cerevisiae L2056 used for alcoholic fermentation was provided by Laboratory of Microbiology ENSIA Massy France. The acetic bacteria strain used for alcoholic fermentation was isolated from a red wine after exposure at ambient temperature. Indeed, 250 mL of red wine was exposed to ambient temperature in open air for 72 h. Afterwards, the wine was used to inoculate Muller Hinton Petri dishes. After 24 h incubation in air at 30°C acetic bacteria were isolated from pure single colonies. The identification was made on the basis of morphological character and biochemical profiles such as gram staining, respiratory metabolism and acid production from ethanol, catalase and oxidase tests. Catalase test was performed by adding one drop H2O2 on a pure microbial colony. The presence of catalase is revealed by the production of bubbles. Oxidase test was performed using oxidase disk (Bio Rad France) as recommended by the manufacturer. Respiratory metabolism of isolated bacteria was assayed by sowing the beef-liver agar semi solid (BioRad, France) then incubated at 30°C for 24 h The acetic acid production from ethanol was carried out in the mid of yeast extract and phenol red (BioRad, France) by addition of ethanol. The identification of gender was completed by the tests of oxidase and catalase.
Production of mango vinegar: The production of mango vinegar was assayed according to the diagram in Fig. 1.
It was carried out in two successive stages Alcoholic fermentation at 30°C for a total period of 144 h using the yeast strain Saccharomyces cerevisiae L2056 and acetic fermentation at 30°C using the acetic bacteria isolated from wine. Briefly, the mango fruit was washed to eliminate the dirt; afterwards the fruits were peeled off. Juice was obtained from the peeled mangoes by mechanic pressure.
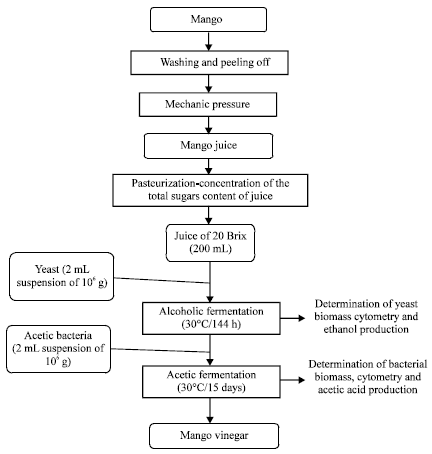 | |
| Fig. 1: | Diagram of mango vinegar production |
The juice was heated at 80°C in order to prevent microbial contaminations and to concentrate sugar until 20 Brix. This was determined using a refractometer (Europe, HP type 0-32). After cooling at room temperature, the juice was distributed in 200 mL sterile bottles. One bottle was used as control to check environmental bacterial contamination. The other bottles were seeded with 2 mL of 106 cfu yeast suspension for 144 h alcoholic fermentation at 30°C. One of these seeded bottles was used for the determination of yeast biomass and alcohol concentration at anytime during the fermentation. After 72 h, the culture was seeded again with 2 mL of 106 cfu acetic bacteria for acetic fermentation. This fermentation was performed at 30°C for a total period of 15 days to have mango vinegar.
Ethanol production: Ethanol contents was determined every 24 h during the alcoholic and acetic fermentations of mango juice by gas chromatography using a Head-Space GC (PERKIN ELMEIR Autosystems XL ®) with a column PEWAX 30 m long, 0.32 mm in diameter. The flow rate of helium column was 3 mL min-1 and the flow rate of carrier gas (hydrogen) Split FLOX was 20 mL min-1.
Acetic acid production: The production of acetic acid was determined every 24 h by titration of 1 mL sample with sodium hydroxide 0.1 N, using phenolphthalein as indicator. The acidity of vinegar expressed in degrees of acetic acid was defined as the mass in gram of acetic acid in 100 g pure vinegar (Lotong et al., 1989).
Physiological state of yeast by flow cytometry: The physiological state of yeast was accessed at the beginning, the middle and at the end of alcoholic and acetic fermentations during the production of vinegar. Theassay is based on the use of fluorescent diacetate carboxy-fluorescein (cFDA) (CHEMUNEX laboratory) and propidium iodide HPLC grade (SIGMA Aldrich Chimie, Saint Quentin and Falladier, France) double-labeling. Propidium iodide penetrates cells through their membrane to color their DNA in red orange. The cFDA is hydrolyzed by cells esterase and their membranes become fluorescent, therefore taking a green color in fluorescent microscope (Sekavova et al., 2005). For the assay, 1 mL fermented juice was centrifuged at 14.000 rpm for 90 sec using a centrifuge EPPENDORF (model Minispin Plus). The pellet was dissolved in a pH4 buffer solution. This solution was divided into two tubes, one receiving 10 μL propidium iodide and being incubated in a Bain Marie (FISHER SCIENTIFIC brand, model Polystat 36) at 40°C for 20 min. Afterwards, the two tubes were then incubated in a Bain Marie at 40°C for further 10 min after introducing of 10 μL of 1/10 cFDA. The observation of cells was carried out on fluorescence microscope (ZEISS brand, model HB050 more AxioStar) equipped with camera (IMASYS mode Scan HD, Wen and PCLK out Synchrogreen) whose images were processed by the software Turbochrom Navigator 4 (CR4). The enumeration of yeast was carried out using a flow cytometer (Dikinson BECTON, FAC Scan model) using the Cell Quest Pro software. The viable cells were stained in green color, stressed cells in yellow and dead cells in red (Hatzis and Porro, 2006).
Statistical analysis: The results of the development of ethanol and acetic acid during the production of vinegar were analyzed by correlation test using the software Excel. Statistical significance was set at p<0.05.
RESULTS
Acetic bacterium identification: After 72 h exposition in air, the cultivation of the red wine on Muller Hinton agar yielded pure single colonies. The Gram staining revealed that the microorganisms were Gram-negative bacilli. The respiratory metabolisms assay revealed that the microorganisms were strictly aerobic and producing acid from ethanol. Catalase and oxidase tests showed that they were catalase and oxidase positive. These characters are the main characters of Acetobacter genus. Thus, the isolated strain was affiliated to this genus and was coded Acetobacter sp. ASV02.
Ethanol and acetic acid productions: The dosage of ethanol during the production of vinegar revealed that, the alcoholic fermentation yielded 1.0 to 22.4 g L-1 ethanol in 72 h (Fig. 2). The monitoring of ethanol level in bottles not seeded with acetic bacteria revealed that, this amount deceased to 14.2 g L-1 after 144 h. Indeed 72 h corresponded to the time to have maximum ethanol in the medium. This time was selected as the starting point for acetic fermentation since ethanol will be used as the substrate for this second fermentation.
The evolution of acetic acid and ethanol during different stages of vinegar production. showed an increase in the production of acetic acid from 0.9 to 4.7° (Fig. 3). This production was inversely proportional to the decline in ethanol from 22.4 to 1.25 g L-1 in the juice of mango after 288 h of acetic fermentation. The stabilization of the alcohol content to 1.25 g L-1 and that of acetic acid to 4.7 degree marked the end of the acetic fermentation. This acetic fermentation transformed 93% (w/w) of ethanol to acetic acid in 15 days. The residual ethanol in vinegar product was maintained at 1.25 g L-1.
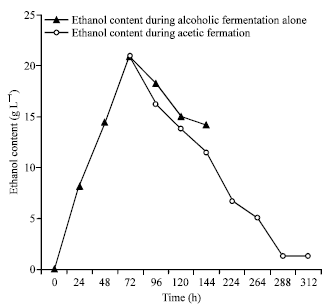 | |
| Fig. 2: | Evolution of ethanol content during mango vinegar production. Ethanol content during alcoholic fermentation alone. The medium is seeded with 106 cfu of Saccharomyces cerevesiae only. Ethanol content during acetic fermentation. The medium is seeded with 106 cfu of Saccharomyces cerevesiae after 72 h the medium is seeded again with 106 cfu of Acetobacter ASV02 |
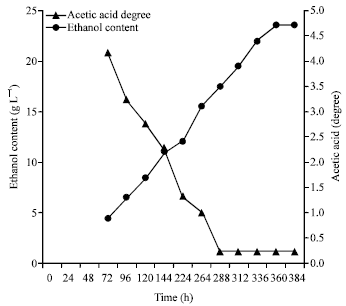 | |
| Fig. 3: | Evolution of the acetic acid and the ethanol content during the production of mango vinegar |
Cytometry of yeast cells: The yeast physiological state monitoring during the production of vinegar was assayed by flow cytometry. The results indicated that 90 to 92% of yeasts were viable 24 h after the alcoholic fermentation (Fig. 4-6). The percentage of viable yeasts decreased from 45% (corresponding to the end of alcoholic fermentation) to zero at the end of the acetic fermentation (360 h after the start of the fermentation).
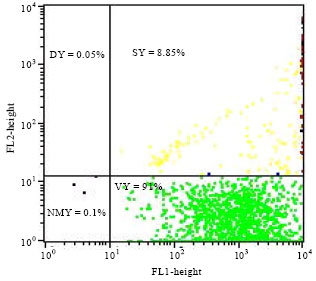 | |
| Fig. 4: | Physiological state of yeast Saccharomyces cerevisiae L2056 population after 20 h of alcoholic fermentation. VY: Viable yeast, DY: Died yeast, SY: Stressed yeast, NMY: No marked yeast FL1: Green Fluorescence intensity and FL2: Red Fluorescence intensity |
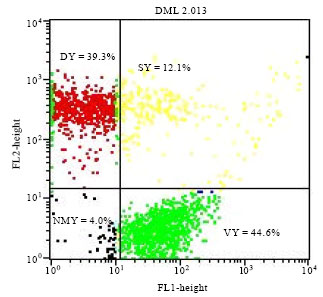 | |
| Fig. 5: | Physiological state of yeast Saccharomyces cerevisiae L2056 population after 96 h of alcoholic fermentation and 24 h of acetic fermentation. VY: Viable yeast, DY: Died yeast, SY: Stressed yeast, NMY: No marked yeast FL1: Green Fluorescence intensity and FL2: Red Fluorescence intensity |
Stressed yeasts decreased from 8.85% after 24 h of fermentation acetic 12.10% to 1.51% after 96 and 360 h, respectively. The decrease of viable yeasts was followed by the increase of dead yeasts.
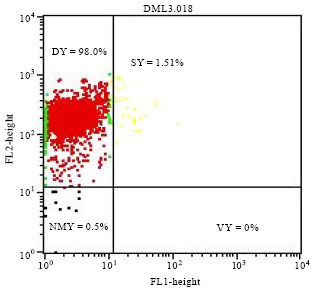 | |
| Fig. 6: | Physiological state of yeast Saccharomyces cerevisiae L2056 population after 360 h of acetic fermentation. VY: Viable yeast, DY: Died yeast, SY: Stressed yeast, NMY: No marked yeast FL1: Green Fluorescence intensity and FL2: Red Fluorescence intensity |
These dead yeasts were estimated at 0.05, 39.30 and 98.00%, respectively after 24, 96 and 360 h acetic fermentation.
DISCUSSION
The present study aimed to access for the physiochemical parameters of vinegar production from mango juice. The exposition of red wine in ambient air allowed us to isolate an acetic bacterium that was able to use ethanol as substrate to produce acetic acid. This bacterium was a Gram-negative bacillus, strictly aerobic, catalase and oxidase positive. According to previous criteria described in several reports the bacterium could belong to Acetobacter genus (Ndoye et al., 2006). However, we could not access for the species, thus the bacterium was coded Acetobacter sp. ASV02. About 17 acetic bacteria have been isolated from tropical and sub-Saharan fruits and used in industrial production of vinegar. Their main characteristic is to produce acetic acid from ethanol. Among them 60% of were identified as Acetobacter though 16S RNA sequencing. The remaining strains were identified as Gluconobacter genus. The two genus differ in biochemical characteristics; Acetobacter is oxidase positive whereas Gluconobacter is oxidase negative.
Vinegar was produced by two consecutive fermentations. During the growing phase of alcoholic fermentation, ethanol production may be proportional to the volume and concentration of sugar juice. The rapid and significant reduction of the ethanol content of wine after fermentation may be due to oxidation of ethanol by dehydrogenases during the acetic fermentation, which constitutes the second stage of the production of vinegar (Frebortova et al., 1997).
The second fermentation by Acetobacter sp ASV02 resulted in 1.25 g L-1 ethanol and 4.7 degree acetic acid contents. According to Valli et al. (2006), the genus Acetobacter commonly used in industrial vinegar production process was involved in biological oxidation of ethanol to acetic acid by the combination of two types of enzymes: an alcohol dehydrogenase (ADH) and aldehyde dehydrogenase (ALDH). Our results indicate that the acetic fermentation of mango juice requires 12 to 15 days for 4.7 degree vinegar. These results were in accordance with those of Ethiraj and Suresh (1990) who, found that the acetic fermentation of the juice of mango peelings produces 4.65 degrees vinegar in 12 days. The process of two consecutive fermentations is rapid than the fermentation by the simple batch that requires 4 to 5 weeks for a complete fermentation (Ethiraj and Suresh, 1990). The study of the evolution of acetic acid and ethanol in the juice Mangovi during the production of vinegar showed the existence of a non-linear relationship (r = -0.98) between both parameters. This suggested independence between the evolution of acetic acid and the evolution of ethanol in the mango juice.
Cytometry study enabled to access to changes in the physiological state of yeast during the production of vinegar. Stressed yeast cells died during the acetic fermentation. The state of stress remains a transitional stage leading to the death of the yeast in the majority of cases. Certainly, alcohol induced stress in S. cerevisiae by causing the flocculation (Claro et al., 2007). However, the stress of yeast is more related to acetaldehyde which is the first product of biological oxidation of ethanol by Acetobacter. That acetaldehyde disturbs the enzymatic activity of yeast (Aranda and Delolmo, 2003). Acetic acid oxidation causes a change of extracellular pH and affects the intracellular pH of the yeast (Valli et al., 2005), reducing considerably its production of ethanol as soon as the acid concentration reached 3% (Graves et al., 2007). Cell death and production of acid during the production of mango vinegar were highly correlated (r = -0.98) suggesting the death of the yeast was linked to the high content of acetic acid.
The present study allowed us to isolate an efficient Acetobacter strain that is under investigation for molecular characterization. The study also allowed the production of vinegar by two consecutive fermentations of local variety mango juice. The process is rapid and non expensive method and easily applicable. The application of the present results may be an alternative for the transformation of local fruits to avoid the post harvest loses.
ACKNOWLEDGMENT
The authors thank MM. K.T. ANANI and K. SONCY of Microbiology Laboratory and Quality Control of Foodstuffs (Togo) for their technical contribution to the achievement of this work.
REFERENCES
- Aranda, A. and M. Delolmo, 2003. Response to acetaldehyde stress in the yeast Saccharomyces cerevisiae involves a strain-dependent regulation of several ALD genes and is mediated by the general stress response pathway. Yeast, 20: 747-759.
PubMed - Claro, F.B., K. Rijbrak and E.V. Soarese, 2007. Flocculation onset in Saccharomyces cerevisiae: Effect of ethanol heat and osmotic stress. J. Applied Microbiol., 102: 693-700.
CrossRef - Ethiraj, S. and E.R. Suresh, 1990. Studies on the utilisation of mango processing wastes for production of vinegar. J. Food Sci. Technol., 29: 48-50.
Direct Link - Frebortova, J., K. Matsushita and O. Adachi, 1997. Effect of growth substrates on formation of alcohol dehydrogenase in Acetobacter methanolicus and Acetobacter aceti. J. Fermen. Bioeng, 83: 21-25.
CrossRef - Hatzis, C. and D. Porro, 2006. Morphologically-structured models of growing budding yeast populations. J. Biotechnol., 124: 420-438.
CrossRef - Kacmar, J., A. Zamamiri, R. Carlson and N.R. Abu-absi and F. Srienc, 2004. Single-cell variability in growing Saccharomyces cerevisiae cell populations measured with automated flow cytometry. J. Biotechnol., 109: 239-254.
PubMed - Kacmar, J., R. Carson, S.J. Balogii and F. Srienc, 2006. Staining and quantification of poly-3-hydroxybutyrate in Saccharomyces cerevisiae and Cupriavidus necator cell populations using automated flow cytometry. Cytometry, 69: 27-35.
PubMed - Lotong, N., W. Malapan and A. Boongorsrang, 1989. Production of vinegar by acetobacter cells fixed on a rotating disc reactor. Applied Microbiol. Biotechnol., 32: 27-31.
CrossRef - Ndoye, B., F. Weekers, B. Diawara, T.A. Guiro and P. Thonart, 2007. Survival and preservation after freeze-drying process of thermoresistant acetic acid bacteria isolated from tropical products of Subsaharan Africa. J. Food Eng., 79: 1374-1382.
CrossRef - Ndoye, B., S. Lebeccque, R. Dubois-Dauphin, L. Tounkanra, A.T. Guiro, C. Kere, B. Diawara and P. Thonart, 2006. Thermoresistant properties of acetic acids bacteria isolated from tropical products of Sub-Saharan Africa and destined to industrial vinegar. Enz Microb. Technol., 39: 916-923.
CrossRef - Raspor, P. and D. Goranovic, 2008. Biotechnological applications of acetic acid bacteria. J. Critic. Rev. Biotechnol., 28: 101-124.
CrossRefPubMedDirect Link - Sekavova, B., K. Melzoch, L. Paulova and M. Rychtera, 2005. Application of flow cytometry to Saccharomyces cerevisiae peculation analysis. Chimia, 59: 745-748.
Direct Link - Valli, M., M. Sauer, P. Branduardi, N. Borth, D. Porro and D. Mattanovich, 2006. Improvement of lactic acid production in Saccharomyces cerevisiae by cell sorting for high intracellular pH. Applied Environ. Microbiol., 72: 5492-5499.
PubMed - Valli, M., M. Sauer, P. Branduardi, N. Borth, D. Porrot and D. Mattanovich, 2005. Intracellular pH distribution in Saccharomyces cerevisiae cell populations, analyzed by flow cytometry. Applied Environ. Microbiol., 71: 1515-1521.
CrossRefDirect Link