
M. Abdulhai
General Commission for Scientific Agricultural Research, Aleppo Center, P.O. Box 4195, Aleppo, Syria
M. El-Bouhssini
ICARDA, P.O. Box 5466, Aleppo, Syria
M. Jamal
Faculty of Agriculture, Damascus University, Damascus, Syria
A. N. Trissi
Faculty of Agriculture, Aleppo University, Aleppo, Syria
Z. Sayyadi
ICARDA, P.O. Box 5466, Aleppo, Syria
M. Skinner
Entomology Research Laboratory, University of Vermont, Burlington, Vermont, USA 05405-0105
B. L. Parker
Entomology Research Laboratory, University of Vermont, Burlington, Vermont, USA 05405-0105
Pakistan Journal of Biological Sciences
Year: 2010 | Volume: 13 | Issue: 21 | Page No.: 1052-1056







ABSTRACT
The objectives of this study were to evaluate the efficacy of Sunn pest entomopathogenic fungi collected from wheat fields and overwintering sites in Syria and characterize them under different temperature regimes. Eleven isolates of Beauveria bassiana (Bals.) Vuil. were included in this study: five were isolated from overwintered adults of Sunn Pest, Eurygaster integriceps Puton and six from nymphs and new adults in wheat fields. Growth rate and conidial production were tested at 15, 20, 25, 30 and 35°C. Results showed that highest growth rate of tested fungal isolates was at 25°C, except for the two isolates SPDR-1 and SPDR-2 which exhibited highest growth rate at 20°C. The highest production of conidia of tested fungal isolates was at 20°C, except for isolates SPSR-1, SPSQ and SPSS which exhibited higher production at 25°C. Virulence test of the tested fungal isolates on overwintered adults Sunn Pest showed that percent mortality after 14 days ranged between 86-100%.
PDF Abstract XML References Citation
Received: July 07, 2010;
Accepted: September 09, 2010;
Published: October 12, 2010
How to cite this article
M. Abdulhai, M. El-Bouhssini, M. Jamal, A. N. Trissi, Z. Sayyadi, M. Skinner and B. L. Parker, 2010. Beauveria bassiana Characterization and Efficacy vs. Sunn Pest, Eurygaster integriceps Puton (Hemiptera: Scutelleridae). Pakistan Journal of Biological Sciences, 13: 1052-1056.
DOI: 10.3923/pjbs.2010.1052.1056
URL: https://scialert.net/abstract/?doi=pjbs.2010.1052.1056
DOI: 10.3923/pjbs.2010.1052.1056
URL: https://scialert.net/abstract/?doi=pjbs.2010.1052.1056
INTRODUCTION
Sunn Pest, Eurygaster integriceps Puton, is one of the major constraints to the production of wheat in Central and West Asia and East Europe (El-Bouhssini et al., 2009). It causes yield losses of 20-30% in barley and 50-90% in wheat. While feeding they inject chemicals into the grain that destroys the gluten and reduces the baking quality of flour (Hariri et al., 2000; Javahery, 1995).
In Syria, in 2005, ~282,000 ha were sprayed with insecticides to control Sunn Pest (Anonymous, 2005). In 2009, IPM strategies were used and the areas sprayed by insecticides were reduced to 87.000 ha (Anonymous 2009). Beauveria bassiana (Deuteromycotina: Hyphomycetes) is an entomopathogenic fungus that has shown great potential for the management of various insect pests (Feng et al., 1994; Inglis et al., 2001; Jaronski and Goettel, 1997) and it is widely used as a microbial control agent in many countries. It is easy to mass produce and has minimal impact on most vertebrates (Boucias and Pendland, 1998). Parker et al. (2000, 2003) collected entomopathogenic fungi of Sunn Pest from Syria, Turkey, Iran, Uzbekistan, Kazakhstan, The Kyrghyz Republic and Russia. Jordan and Pascoe (1996) collected them from N. Iraq. Isolates from the genera Beauveria, Paecilomyces (some sp. recently placed in the genus Isaria) and Verticillium (some sp. recently placed in the genus Lecanicillium) were collected (Humber, 2010). Beauveria bassiana was the most commonly recovered species, suggesting that this pathogen may hold the most promise for management.
Many studies have been made on the response of B. bassiana to a wide range of temperatures in vitro (Ekesi et al., 1999; Fargues et al., 1997; James et al., 1998; Shimazu, 2004; Sivasankaran et al., 1998). Parker et al. (2003) tested isolates of B. bassiana at 20, 25, 30 and 35°C for growth and sporulation. Usually, the optimal temperature for growth of entomopathogenic fungi ranges between 20-25°C, but this also depends on species and strain (Glare and Milner, 1991; Goettel and Inglis, 1997).
The objectives of the research herein were to characterize fungal temperature responses and evaluate the efficacy of isolates collected from Sunn Pest wheat field populations and from populations in diapauses in overwintering sites in Syria.
MATERIALS AND METHODS
Insect collection and fungal isolation: Isolates of B. bassiana were collected in the winter in 2006-2007 from overwintering sites of Sunn Pest. Adults that exhibited signs of mycelial growth were put in separate plastic bags lined with paper towels to absorb excess moisture and brought to the laboratory. Isolates were also collected in the summer from infested wheat fields in all provinces in Syria. Three sites in each province were randomly selected and 50 nymphs and new adults of Sunn Pest were hand-collected from each site. They were placed (by site) in plastic bags and returned to the laboratory. Sunn Pest were killed by placing them at -20°C for two hours; surface sterilized by immersion in 70% EtOH for 15 sec, rinsed in distilled water for 10 sec; followed by a 1 min immersion in 1% sodium hypochlorite; rinsed for 10 sec in 3 changes of sterile distilled water and blot-dried with sterile filter paper (Lacey and Brooks, 1997). Insects were placed in 9 cm diam plastic Petri dishes on sterile filter paper moistened with 0.001 g L-1 penicillin G (25 million units g-1) and 0.005 g L-1 streptomycin sulfate. The dishes were sealed with parafilm and held at 22±2°C for 2 weeks. for fungal outgrowth. Fungal isolations were made on ¼ SDAY supplemented with penicillin (0.03%) and streptomycin sulfate (0.08%) (Goettel and Inglis, 1997). Isolates were subcultured several times to ensure purity. Fungi were identified under 400x magnifications using the keys of Humber (1997).
Fungal test material: Isolates for bioassay were cultured on ¼ SDAY at 22±2°C for 14 days. Fresh conidia were harvested from the plates by scraping and then suspended by vortexing in 0.01% Tween 80. The suspension was filtered through eight layers of cheesecloth to remove the media and mycelia. Conidia were enumerated using a hymocetometer and their density adjusted to the target concentration by dilution (Goettel and Inglis, 1997). Conidial viability for each isolate was determined by inoculating a plate of ¼ SDAY with 0.1 mL of a conidial suspension containing 1×106 conidia mL-1. After incubation at 20°C for 24 h, the rate of spore germination was determined by counting 100 spores in 4 different fields of view (400 spores per plate). A spore with a germ tube longer than its width was considered germinated (Hywell-Jones and Gillespie, 1990). Suspension concentration was adjusted according to conidial viability when necessary to compensate for lower viability.
Isolate characterization: Isolates were characterized according to their growth and sporulation on ¼ SDAY medium at 15, 20, 25, 30 and 35°C (±1°C). Five microliters of a 1×106 conidial suspension were pipette onto a 0.64-cm diam. desk of filter paper and placed in the center of 5 Petri dishes containing 20 mL of media. Colony growth was marked on the dish bottom at the outer edge of the fungal growth at 5, 10, 15 and 20 day after inoculation and measured from the center of the dish to the mark. Conidial production was determined from four 5 mm diam discs cut from the colony on the last sampling day. The first disc was taken close to the leading edge of new growth and the remaining discs were taken within the previously marked 5 day growth zones for days 5, 10 and 15. The four discs from each plate were combined in the test tube containing 10 mL 0.01% Tween 80 in sterile distilled water and 12 glasses beads. Tubes were agitated for 60 sec to dislodge conidia. The numbers of conidia per unit volume were determined using a hemocytometer under 400x magnifications. The data were converted to numbers of conidia per unit surface area.
Insect assays: Sunn pest adults were collected from overwintering sites beneath the litter of pine trees at Tel Hadya, Syria 1 day prior to testing and held in plastic vented containers (25×15×15 cm) at 5°C until used. All insects were inspected before testing to ensure they were alive and only those that moved when probed with a blunt needle were used in the trial. Single-concentration assays (at 1×106 conidia/mL 0.01% Tween 80) were conducted in February and March 2009. A control, 0.01% Tween 80 suspension, was included in each assay. Each treatment was replicated five times within the trial. Ten adults were fixed on adhesive tape – ventral surface up and 5 μL of 1×106 conidia/mL 0.01% Tween 80 suspension was pipetted on the sternum and allowed to dry. Adults were then grouped in a clear, screw top plastic jar (5.5 cm diam 5.7-cm hgt) containing fresh wheat leaves. The jars were held in the laboratory at room temperature (22±2°C) under natural light conditions. Mortality counts were taken 5, 10, 12 and 14 days post treatment. Adults were considered dead if they failed to move 2 min following slight probing. Sunn Pest adults are known to feign death.
Statistical analyses: Data on rates of fungal colony growth (mm day-1) over the 20 day experiment and on the effect of temperature on conidial production (conidia cm-1) were analyzed as completely randomized designs using a two-way ANOVA (no blocking). Prior to analysis, a natural log transformation was performed on the conidial count data to improve homogeneity of variance.
The percentages of mortality were arcsine-transformed and analyzed using ANOVA. The means and the standard errors reported in the tables are based on data before transformation.
RESULTS
Insect collection and fungal isolation: Five isolates of B. bassiana were isolated from overwintering adults of Sunn Pest in overwintering sites.
| Table 1: | B. bassiana isolates collected from fields and overwintering sites in Syria |
 | |
| Table 2: | Growth rate (mm day-1) of B. basiiana isolates cultured at different temperatures for 20 days |
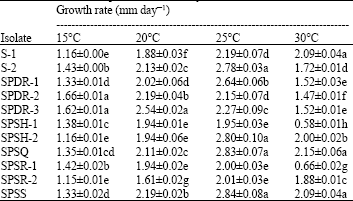 | |
| Values are as Mean±SE of 5 replicates. Numbers followed by the same letters within a column are not significantly different (p<0.05) | |
From 5850 nymphs and new adults of Sunn Pest collected from wheat fields, six isolates of B. bassiana were discovered and were called summer isolates (Table 1).
Fungal characterization: None of the isolates grew at 35°C and all were dead after 20 day, These temperature data were excluded from the analyses. There were significant differences among the isolates in their growth rate at each temperature and among the temperatures for each isolate (Table 2) (p<0.001 for Temperature, Isolate and Isolate *Temperature). The highest growth of 8 isolates was at 25°C, while two isolates, SPDR-1 and SPDR-2 exhibited their highest growth at 20°C. The growth of SPSH1 did not differ at 20 and 25°C. Growth rate was reduced for all isolates at >25°C or it decreased at <20°C.
Production of conidia for all isolates stopped at 35°C and for SPSH-1 at 30°C. More conidia were produced at 20°C for 8 of the isolates, while SPSR-1, SPSQ and SPSQ, all summer isolates produced more conidia at 25°C (Table 3). However, fungal isolates and temperatures significantly interacted (p<0.001) in their effect on conidial production. S-1 was the highest conidial producer at 20°C and 15°C. At 25 and 30°C, S-1 was the third conidial producer after SPSS and SPSQ, indicating that this isolate is not sensitive to the lower and higher temperatures.
| Table 3: | Number of conidia produced by entomopathogenic fungus B. bassiana when cultured at 15, 20, 25 and 30°C |
 | |
| Values are as Mean±SE of 5 replicates. Numbers followed by the same letters within a column are not significantly different (p<0.05) | |
| Table 4: | Percent mortality of Eurygaster integriceps when treated with 11 isolates of B. bassiana after 5, 10, 12 and 14 days |
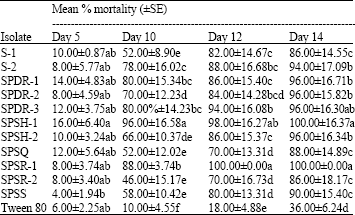 | |
| Values are as Mean±SE of 5 replicates, Tween 80 (control) means no fungal treatment. Numbers followed by the same letters within a column are not significantly different (p<0.05) | |
At 25°C SPSS was the highest conidial producer, while at 30°C SPSQ was the highest.
Insect assays: At 5 day post treatment there were no differences in mortality between isolates and the Tween control, while at 10, 12 and 14 day isolates had mortality significantly greater than the Tween control (Table 4). At day 10 SPSH-1 caused greater mortality than the other isolates and reached 96%. At day 12 SPSR-1 caused greater mortality (100%) than the other isolates. At day 14, mortality ranged between 86 and 100%. The highest mortality was caused by SPSR-1 and SPSH-1. Fungal isolates and days post treatment significantly interacted (p<0.001) in their effect on Sunn Pest mortality.
DISCUSSION
The lack of colony growth and the ultimate mortality of all isolates tested at 35°C corroborates earlier findings. The growth rates obtained in this study in response to different temperatures are similar to that reported previously (Parker et al., 2003; Fargues et al., 1997; Hallsworth and Magan, 1999; Mietkiewski et al., 1994; Roddam and Rath, 1997; Shimazu, 2004; Tong-Kwee et al., 1989). The two isolates SPDR-1 and SPDR-2 that exhibited the highest growth rate at 20°C were collected form overwintering site from Rhankos, Reef Demashk with an elevation of 1500 m above sea level and where temperatures could reach -30°C in winter. The strong interactions observed between temperatures and isolates showed the necessity to characterize isolates for their sensitivity to temperature before their use in different environments. (Bidochka et al., 2001; Fargues et al., 1997).
The sporulation of most isolates at tested temperatures corroborates earlier findings showing no sporulation of B. bassiana isolates at or above 30°C. Present results showing highest production of conidia at 20°C for 8 isolates do not support those of Parker et al. (2003) who reported highest production of conidia of B. bassiana isolates at 25°C. However, conidial production of the three summer isolates (SPSR-1, SPSQ and SPSS) was similar to that reported by Parker et al. (2003).
Results of this study will be helpful in selecting the appropriate fungal isolate for use in an integrated pest management program against Sunn Pest in the target environment.
REFERENCES
- Bidochka, M.J., A.M. Kamp, T.M. Lavender, J. Deconing and J.N.A. Cross, 2001. Habitate association in two genetic groups of the insect-pathogenic fungus Metarhizium anisopliae: Uncovering cryptic species. Applied Environ. Microbiol., 67: 1334-1342.
CrossRef - Ekesi, S., N.K. Maniania and K. Ampong-Nyarko, 1999. Effect of temperatures on Germination, radial growth and virulence of Metarhizium anisopliae and Beauveria bassiana on Megalurothrips sjostedti. Biocont. Sci. Technol., 9: 177-185.
Direct Link - El-Bouhssini, M., K. Street, A. Joubi, Z. Ibrahim and F. Rihawi, 2009. Sources of wheat resistance to Sunn pest, Eurygaster integriceps Puton, in Syria. Genet. Resour. Crop Evol., 56: 1065-1069.
Direct Link - Fargues, J., M.S. Goettel, N. Smits, A. Ouedraogo and M. Rougier, 1997. Effect of temperature on vegetative growth of Beauveria bassiana isolates from different origins. Mycologia, 89: 383-392.
Direct Link - Feng, M.G., T.J. Poprawski and G.G. Khachatourians, 1994. Production, formulation and application of the entomopathogenic fungus Beauveria bassiana for insect control: Current status. Biocont. Sci. Technol., 4: 3-34.
CrossRefDirect Link - Hallsworth, J.E. and N. Magan, 1999. Water and temperature relation of growth of the entomogenous fungi Beauveria bassiana, Metarhizium anisopliae and Paecilomyces farinosus. J. Invertebrate Pathol., 74: 261-266.
Direct Link - Hariri, G., P.C. Williams and F.J. El-Haramein, 2000. Influence of pentatomid insects on the physical dough properties and two-layered flat-bread baking quality of syrian wheat. J. Cereal Sci., 31: 111-118.
CrossRef - Hywel-Jones, N.L. and A.T. Gillespie, 1990. Effect of temperature on spore germination in Metarhizium anisopliae and Beauveria bassiana. Mycol. Res., 94: 389-392.
Direct Link - Inglis, G.D., M.S. Goettel, T.M. Butt and H. Strasser, 2001. Use of Hyphomycetous Fungi for Managing Insect Pests. In: Fungi as Biocontrol Agents, Butt, T.M., C. Jackson and N. Magan (Eds.). CAB International, Wallingford, UK., pp: 23-69.
Direct Link - James, R.R., B.A. Croft, B.T. Shaffer and B. Lighthart, 1998. Impact of temperature and humidity on host-pathogen interactions between Beauveria bassiana and a coccinellid. Environ. Entomol., 27: 1506-1513.
Direct Link - Jaronski, S.T. and M.S. Goettel, 1997. Development of Beauveria bassiana for control of grasshoppers and locusts. Mem. Entomol. Soc. Can., 171: 225-237.
Direct Link - Parker, B.L., M. Skinner, S.D. Costa, S. Gouili, W. Rreid and M. El Bouhssini, 2003. Entomopathogenic fungi of Eurygaster integriceps puton (Hemiptera: Scutelleridae) collection and characterization for development. J. Biol. Contr., 27: 260-272.
CrossRefDirect Link - Shimazu, M., 2004. Effects of temperature on growth of Beauveria bassiana F-263, a strain highly virulent to the Japanese pine sawyer, Monochamus alternatus, especially tolerance to high temperatures. Applied Entomol. Zool., 39: 469-475.
Direct Link - Tong-Kwee, L., R. Muhamad, G.C. Fee and C.C. Lan, 1989. Studies on Beauveria bassiana isolated from the cocoa mirid, Helopeltis theobromae. Crop Protec., 8: 358-362.
CrossRef