
W. Pitakdantham
Center for Agricultural Biotechnology (CAB), Kasetsart University, Kamphang Saen Campus, Nakhon Pathom 73140, Thailand
T. Sutabutra
Department of Plant Pathology, Faculty of Agriculture, Kasetsart University, Bangkok 10900, Thailand
P. Chiemsombat
Department of Plant Pathology, Faculty of Agriculture, Kasetsart University, Kamphang Saen Campus, Nakhon Pathom 73140, Thailand
C. Pitaksutheepong
National Center for Genetic Engineering and Biotechnology (BIOTEC), 113 Phaholyothin Road, Klong 1, Klong Luang, Pathumthani 12120, Thailand
Pakistan Journal of Biological Sciences
Year: 2010 | Volume: 13 | Issue: 20 | Page No.: 1000-1005







ABSTRACT
To isolate and characterize chalcone synthase gene in anthocyanin biosynthetic pathway during flower development of Dendrobium Sonia Earsakul. The gene was isolated from floral tissues of the orchid by reverse transcriptase polymerase chain reaction. Characterization of the gene considered to its relatedness to chalcone synthase gene in other orchid plants elucidated by construction of a neighbor-joining phylogenetic tree. Gene expression pattern related to flower development and pigmentation was investigated by relative quantification real time polymerase chain reaction. A complete coding sequence was obtained and sequence analysis revealed that the gene of Dendrobium Sonia Earsakul consisted of 1,188 bp. Blast analysis and multiple alignments showed that the chalcone synthase gene of Dendrobium Sonia Earsakul shares high homology to chalcone synthase gene of Dendrobium genus particularly Dendrobium hybrid Uniwai prince. Phylogenetic tree revealed that chalcone synthase of Dendrobium genus are highly conserved. The chalcone synthase gene of Dendrobium Sonia Earsakul was highly expressed in young flower bud with no pigmentation and the expression was sharply decreased when young flower bud started accumulation of pigments. Expression of chalcone synthase gene was then maintained at the same level until young bud developed into fully opened flowers.
PDF Abstract XML References Citation
Received: June 24, 2010;
Accepted: August 16, 2010;
Published: September 24, 2010
How to cite this article
W. Pitakdantham, T. Sutabutra, P. Chiemsombat and C. Pitaksutheepong, 2010. Isolation and Characterization of Chalcone Synthase Gene Isolated from Dendrobium Sonia Earsakul. Pakistan Journal of Biological Sciences, 13: 1000-1005.
DOI: 10.3923/pjbs.2010.1000.1005
URL: https://scialert.net/abstract/?doi=pjbs.2010.1000.1005
DOI: 10.3923/pjbs.2010.1000.1005
URL: https://scialert.net/abstract/?doi=pjbs.2010.1000.1005
INTRODUCTION
Anthocyanin is a type of flavonoid that is a major factor in flower pigmentation. It contributes a range of colors from red to margenta, orange to red and purple (Tanaka et al., 2005). Biosynthesis of these pigments is controlled by a series of enzymes. Chalcone synthase gene (chs) encodes chalcone synthase (CHS) which is an important enzyme in anthocyanin biosynthetic pathway. CHS supplies chalcone, the precursor of anthocyanin molecules, via processes of decarboxylation, condensation, cyclization and aromatization reaction of p-coumaroyl and three molecules of malonyl-CoA (Ferrer et al., 1999).
Chalcone synthase was investigated in many plant species. Their flower colors were successfully manipulated through modification this structural gene such as Torenia Petunia and Gentian (Fukusaki et al., 2004; Hanumappa et al., 2007; Nakatsuka et al., 2008; Suzuki et al., 2000; Tanaka et al., 1998). In orchids, there are few data reported in this anthocyanin biosynthetic gene such as Bromheadia (Liew et al., 1998) Phalaenopsis (Han et al., 2006), while only one publication was reported in Dendrobium hybrids (Mudalige-Jayawickrama et al., 2005).
Dendrobium Sonia Earsakul is widely cultivated for commercial production as cut flower orchid in Thailand. Its flower color is purple. To gained insight in color production of this orchid plants. We isolated cDNA clone of chs gene. BLAST analysis and phylogenetic tree was performed to see their relationship to those of orchids. Expression profile of gene in floral tissue at five developmental stages was investigated by real time PCR analysis.
MATERIALS AND METHODS
RNA extraction: Flower tissues of D. Sonia Earsakul grown at five stages as shown in Fig. 1 were pooled and utilized for total RNA extraction by modified CTAB method described by Sambrook et al. (1989). For relative quantification real time PCR, total RNA of flower tissues grown in different developmental stages (Fig. 1) were separately extracted as described above.
Cloning the chs gene: The first strand cDNA was synthesized from total RNA using MMLV reverse transcriptase (Fermentas, Canada). OligodT including M13 reverse sequences, 5′CAG GAA ACA GCT ATG ACC ATG TTT TTT TTT TTT TTT TT 3′, was used as a primer. Degenerate primers, 3′endF 5′TAT CCG GAY TAC TAC TTC AGR ATT ACC A 3′ and 5′endR 5′GGC GTT GTT CTC GGC GAG GTC TTT GGC3′, were designed from a conserved region of chs genes from many plant species and employed to amplify partial chs gene. Degenerate and specific primers, 5′endF 5′ GAA TAG GGA GGG AGT TAA TTA ATG GC 3′ and 5′endR were used to amplify 5′end of the chs gene while a pair of primers 3′endF and M13R 5′CAG GAA ACA GCT ATG ACC ATG 3′ were used to amplify the 3′end of the gene. The amplified fragments were ligated to pDrive cloning vector and used for transformation of E.coli DH5α. All positive clones containing the inserts were subjected to sequencing analysis.
Sequence analysis: Nucleotide and amino acid sequence were analyzed using BLAST SEARCH (http://blast.ncbi.nlm.nih.gov/Blast.cgi) and clustalW (http://www.ebi.ac.uk/Tools/clustalw2/). Protein analysis system (http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) was used to predict the simple physical properties of the sequence and functional domains of the protein.
Phylogenetic tree analysis: Multiple alignment of amino acid sequences were compared among chs from various orchids and other plant species using clustalW program. The phylogenetic tree was constructed using neighbor joining method.
Relative quantification real time PCR analysis: The expression of chs gene at different growth stages of flower buds was investigated by relative real time PCR. The 18S rRNA was used as the reference control. Relative changed in chs gene expression was calculated by using the 2-ΔΔCt method (Livak and Schmittgen, 2001). Flower tissues of D. Sonia Earsakul were separated into five growth stages (Fig. 1). The total RNA of each stage was used as the RNA templates. The reaction of 15 μL consisted of 7.5 μL RNA direct MASTER with SYBR I dye chemistry, 0.5 μM of each primer (a primer set of forward primer 5'AAT CGA ATC ATG CTT TAC CAA CAA GGC TGC 3'and reverse primer 5'TGA ACA AAC GAC GAG AAC TCG 3' were used for partial chs gene amplification and a primer set of forward primer 5'GCT ACT CGG ATA ACC GTA GT 3'and reverse primer 5'ACC AGA CTT GCC CTC CAA TG 3' were employed to amplification of 18S rRNA gene) and 150 ng of RNA template. RT-PCR was performed by denaturation at 95°C for 1 min and followed by reverse transcription at 61°C for 20 min. PCR was immediately done after reverse transcription was completed by denaturation the DNA at 95°C for 1 min followed by 45 cycles of 95°C for 20 s, 58°C for 20 sec and 72°C for 45 sec. DNA melting was performed after amplification had completed to allow fluorescence measurements of non specific products and analyze of expected DNA product by melting curve analysis.
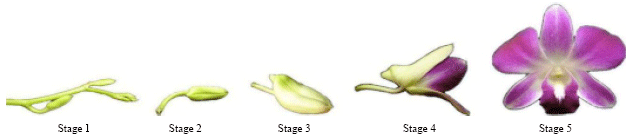 | |
| Fig. 1: | Dendrobium Sonia Earsakul flower was categorised into five growth stages. Stage 1 is the early stage of flower budding. Stage 2 is the closed bud without pigmentation. Stage 3 is the near-open bud with slight pigmentation. Stage 4 is the opened bud. Stage 5 is the fully opened flower |
This research was carried out at Center for Agricultural Biotechnology, Kasetsart University Nakhon Pathom, Thailand. Dendrobium Sonia Earsakul was cultivated in nursery at Plant Unit, National Center for Genetic Engineering and Biotechnology (BIOTEC), Thailand.
RESULTS
Isolation of chs gene from D. Sonia Earsakul: To isolate a chs gene, first strand cDNA of D. Sonia Earsakul was synthesized by reverse transcription using oligodtM13R primer. Subsequently, a conserved region of chs gene sequence was amplified using primers named 3'endF and 5'endR (Fig. 2) and a fragment of about 440 bp was obtained. Sequence analysis revealed that the gene fragment showed high homology to chs genes of orchid plants. To obtain a full-length coding sequence, the 3' and 5' ends of the chs gene were PCR-amplified using primers as detailed in Fig. 2. Fragments of 663 and 552 bp were obtained after amplification of the 3' and 5' ends. Gene assembly of the three fragments using SeqMan software (DNA Star, Inc.) showed that a complete coding sequence of chs gene was obtained. The gene comprised of 1,188 bp and encodes 395 amino acids with the calculated molecular mass of 43.04 kDa and isolectric point (pI) of 6.22. The complete cDNA and deduced amino acid sequences were deposited in GenBank with accession No. AM490639 and CAM32716, respectively.
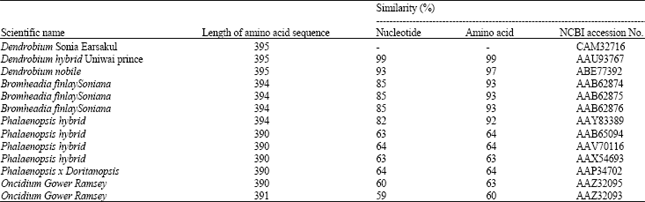
Characterization of chs gene: Protein-protein Blast analysis of deduced amino acid sequence revealed that CHS of D. Sonia Earsakul has high homology to those of Dendrobium genus. The CHS contains putative conserved domains which are the characteristics of chalcone synthase. Multiple alignment of orchid CHS using ClustalW confirmed that the CHS of D. Sonia Earsakul shares high degree of identity to those of CHS isolated from Dendrobium orchids like D. hybrid Uniwai prince (99%) and D. nobile (97%) (Table 1). The CHS of D. Sonia Earsakul also showed high similarity to those of Bromheadia genus (93%). However, the CHS of D. Sonia Earsakul shares less similarity to those isolated from Oncidium and Phalaenopsis orchids, except for one cultivar of Phalaenopsis (AAY83389).
Phylogenetic analysis of chalcone synthase: A phylogenetic tree of plant CHS proteins was constructed based on amino acid sequences (Fig. 3). The tree showed two clades of CHS proteins.
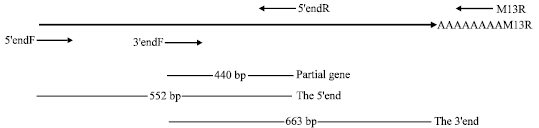 | |
| Fig. 2: | The schematic representation of chs gene fragments of D. Sonia Earsakul isolated by RT-PCR. AAAAAAM13R was used as a primer for first strand cDNA was synthesized. Primers and 5'endR were used to amplify 440 bp. partial chs gene. Primers 5'endF and 5'endR were used to amplify 552 bp. 5'end of the chs gene. Primers 3'endF and M13R were used to amplify 663 bp the 3'end of the gene |
| Table 1: | Percentage of similarity at nucleotides and amino acid sequences of chs cDNA of D. Sonia Earsakul comparison with other orchid species |
 | |
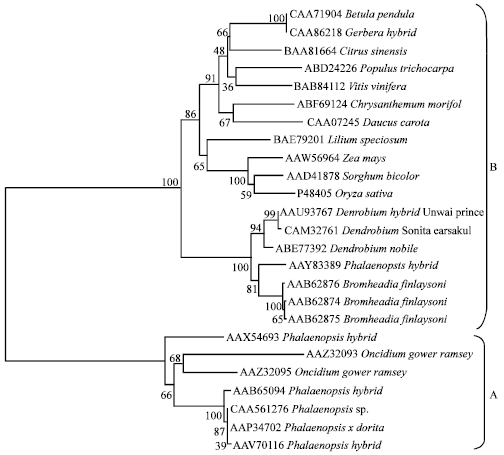 | |
| Fig. 3: | The phylogenetic tree of plant CHS. The tree of CHS proteins was generated using clustalW and MEGA4. Numerals next to the branch represent percentage of bootstap values from 1,000 replications. The bar indicates an evolutionary distance of 0.02%. Accession number in the database of each chs protein was placed in front |
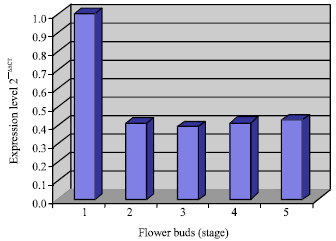 | |
| Fig. 4: | Relative quantification of the chs gene expression from flower buds of D. Sonia earsakul |
The CHS of D. Sonia Earsakul was grouped with those of Dendrobium, Bromheadia, a Phalaenopsis plant and those of other monocots and dicots whereas the CHS of Oncidium and Phalaenopsis orchids fell into the other clade.
Expression of chs gene in flower of D. Sonia Earsakul: Relative quantification real time PCR analysis revealed that the chs gene was highly expressed in young flower buds (stage 1, Fig. 1) with no pigmentation (Fig. 4). The expression was sharply decreased when young flower buds started to develop and accumulation of pigments was seen. From this developmental stage, the level of expression was maintained at nearly the same level throughout to the end of flower development.
DISCUSSION
A complete cDNA of chs gene of D. Sonia Earsakul was obtained. The gene comprised of 1188 bp and encodes 395 amino acids. The CHS of D. Sonia Earsakul shared high degree of homology to those of Dendrobium species and to a lesser extend to those of Bromheadia orchid. Alignment of CHS of D. Sonia Earsakul to those of Phalaenopsis and Oncidium plants revealed that they shared less similarity. Phylogenetic tree of CHS proteins revealed that the CHS of D. Sonia Earsakul was categorized into the same clade as Dendrobium CHS whereas the CHS proteins of Phalaenopsis and Oncidium orchids were grouped in the other clade.
Sequence analysis showed that the deduce amino acid sequence contains feature domains of which are characteristics of chalcone synthase as shown in Fig. 3. This suggested that the CHS could be function in the processes of decarboxylation, condensation, cyclization and aromatization reaction of p-coumaroyl and three molecules of malonyl-CoA (Ferrer et al., 1999). Gene expression pattern of chs gene in D. Sonia Earsakul is similar to that in other plant such as Oncidium (Chiou and Yeh, 2008) Phalaenopsis (Han et al., 2005) and Gentian (Nakatsuka et al., 2005). The expression pattern of chs gene showed that the gene was expressed in all flower development stages. This finding is consistent with the fact that chs gene is a common enzyme necessary for flavone and anthocyanin biosynthesis (Martin et al., 1991; Nakatsuka et al., 2005). The expression of chs gene is highest at the early stage of flower formation. This might reflect the fact that chs gene is one of early biosynthetic genes of flavone and anthocyanin pathways (Davies, 2004; Martin and Gerats, 1993; Nakatsuka et al., 2005; Pelletier et al., 1999). Its product is subsequently required for production of structural genes and enzymes downstream in all flavone and anthrocyanin biosynthesis (Nakagawa et al., 2008).
The most important basic for manipulating orchid flower color is to gain information of the key enzymes in anthocyanin synthetic pathway. In this study, the data of chs gene from D. Sonia Earsakul through gene isolation, characterization and gene expression during flower development could be a tool for flower color manipulation in this plant.
ACKNOWLEDGMENTS
This research is supported by the Center for Agricultural Biotechnology, Postgraduate Education and Research Development Office, Commission on Higher Education, Ministry of Education. And it was partially supported by thesis and dissertation support fund, Graduate school, Kasetsart University.
REFERENCES
- Chiou, C.Y. and K.W. Yeh, 2008. Differential expression of MYB gene (OgMYB1) determines color patterning in floral tissue of oncidium gower ramsey. Plant Mol. Biol., 66: 379-388.
CrossRef - Han, Y.Y., F. Ming, W. Wang, J.W. Wang, M.M. Ye and D.L. Shen, 2006. Molecular evolution and functional specialization of chalcone synthase superfamily from phalaenopsis orchid. Genetica, 128: 429-438.
CrossRefDirect Link - Livak, K.J. and T.D. Schmittgen, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 25: 402-408.
CrossRefDirect Link - Martin, C. and T. Gerats, 1993. Control of pigment biosynthesis genes during petal development. Plant Cell, 5: 1253-1264.
Direct Link - Martin, C., A. Prescott, S. Mackay, J. Bartlett and E. Vrijlandt, 1991. Control of anthocyanin biosynthesis in flowers of Antirrhinum majus. Plant J., 1: 37-49.
PubMed - Nakatsuka, T., M. Nishihara, K. Mishiba and S. Yamamura, 2005. Temporal expression of flavonoid biosynthesis-related genes regulates flower pigmentation in gentian plants. Plant Sci., 168: 1309-1318.
CrossRef - Nakatsuka, T., K.I. Mishiba, Y. Abe, A. Kubota, Y. Kakizaki, S. Yamamura and M. Nishihara, 2008. Flower color modification of gentian plants by RNAi-mediated gene silencing. Plant Biotechnol., 25: 61-68.
CrossRefDirect Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 1st Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
Direct Link - Suzuki, K.I., H.M. Xue, Y. Tanaka, Y. Fukui and M. Fukuchi-Mizutani et al., 2000. Flower color modifications of torenia hybrida by cosuppression of anthocyanin biosynthesis genes. Mol. Breed., 6: 239-246.
CrossRef - Tanaka, Y., Y. Katsumoto, F. Brugliera and J. Mason, 2005. Genetic engineering in floriculture. Plant Cell. Tissue Organ Cult., 80: 1-24.
CrossRefDirect Link