
M. Anoush
Department of Pharmacology and Toxicology, Faculty of Pharmacy, Tabriz University of Medical Sciences, Tabriz, Iran
M.A. Eghbal
Department of Pharmacology and Toxicology, Faculty of Pharmacy, Tabriz University of Medical Sciences, Tabriz, Iran
F. Fathiazad
Department of Pharmacognosy, Faculty of Pharmacy, Tabriz University of Medical Sciences, Tabriz, Iran
H. Hamzeiy
Department of Pharmacology and Toxicology, Faculty of Pharmacy, Tabriz University of Medical Sciences, Tabriz, Iran
N.S. Kouzehkonani
Drug Applied Research Center, Tabriz University of Medical Sciences, Tabriz, Iran
Pakistan Journal of Biological Sciences
Year: 2009 | Volume: 12 | Issue: 10 | Page No.: 765-771







ABSTRACT
Acetaminophen, the most commonly sold over-the-counter antipyretic analgesic, is capable of causing severe and sometimes fatal hepatic damage in humans and experimental animals. The incidence of liver injury due to acetaminophen overdose, either with suicidal intent or by accident, is increasing. Garlic is among those medicinal plants famous for its different health protective effects. In this study, the protective effects of garlic extract on acute acetaminophen-induced liver injury were investigated using freshly isolated rat hepatocytes. The hepatocytes were isolated from Sprague-Dawley male rats by a two step collagenase model. Formation of Reactive Oxygen Species (ROS) and Glutathione (GSH) depletion were studied after addition of acetaminophen to cell suspensions. The effects of garlic extract on prevention of ROS formation as well as GSH depletion was investigated and compared with the effects of N-Acetyl Cysteine (NAC) as the standard treatment. Reactive oxygen species formation was assessed by a spectrofluorometry method and garlic extract was shown to be as effective as NAC in decreasing ROS formation induced by acetaminophen. Glutathione (GSH) levels of hepatocytes were determined using HPLC. Garlic extract was effective in preventing GSH depletion significantly (p<0.05). It is concluded that garlic extract has an antioxidant effect and can protect hepatocytes from GSH depletion following NAPQI production.
PDF Abstract XML References Citation
How to cite this article
M. Anoush, M.A. Eghbal, F. Fathiazad, H. Hamzeiy and N.S. Kouzehkonani, 2009. The Protective Effects of Garlic Extract Against Acetaminophen-Induced Oxidative Stress and Glutathione Depletion. Pakistan Journal of Biological Sciences, 12: 765-771.
DOI: 10.3923/pjbs.2009.765.771
URL: https://scialert.net/abstract/?doi=pjbs.2009.765.771
DOI: 10.3923/pjbs.2009.765.771
URL: https://scialert.net/abstract/?doi=pjbs.2009.765.771
INTRODUCTION
For many centuries, garlic (Allium sativum) has been widely used as a flavor and as one of the nutrients which prevent different types of diseases. Earlier studies have shown that garlic possesses a wide range of biological activities, such as anti-atherosclerotic, antihypertensive, anticancer, immune-modulator, antimicrobial and potential anti-aging effects (Agarwal, 1996; Banerjee et al., 2003; Amagase, 2006). The main organosulfur compound in intact garlic bulbs, 7-glutamylcysteines, can be hydrolyzed and oxidized into alliin and then to allicin, which has been mistaken as the active component of garlic. Allicin is unstable and decomposed into the odorous compounds, i.e., Allyl Mercaptan (AM), diallyl sulfide (DAS), diallyl disulfide (DADS) and diallyl trisulfide (DATS) (Amagase et al., 2001; Xu and Simon Cho, 2000; Zeng et al., 2008).
It has been shown that high doses of the analgesic drug acetaminophen (APAP) produce centrilobular liver necrosis in human and other susceptible species (Ruepp et al., 2001; Sumioka et al., 2004; Aleksunes et al., 2008).
Acetaminophen is primarily metabolized by sulfation and glucuronidation, but with an increasing dose rate, these pathways become saturated and a greater proportion of the drug is available for oxidation by the microsomal cytochrome P-450 system (Amar and Schiff, 2007). N-Acetyl-P-benzoquinone Imine (NAPQI) is the product of this pathway which is thought to be responsible for the subsequent hepatic damage (Aleksunes et al., 2008).
N-Acetyl-P-benzoquinone Imine (NAPQI) is a highly reactive electrophile and is detoxified in liver by either reduction to the parent compound, acetaminophen, or conjugation at the meta-position with glutathione, which both reactions consume GSH (Ruepp et al., 2001). The importance of glutathione in acetaminophen toxicity is further emphasized by the large body of evidence which indicates that interventions which increase GSH content can dramatically reduce acetaminophen and NAPQI-induced hepatic injury (Mitchell, 1977; Ruepp et al., 2001).
Oxidative stress is also considered to be involved in the induction of hepatotoxicity by APAP. Oxidation of APAP by CYPs may generate Reactive Oxygen Species (ROS). Hydrogen peroxide and superoxide are produced during metabolic activation of APAP in the mixed function oxidase system (Amimoto et al., 1995).
Some in vivo studies have mentioned the antioxidant and hepatoprotective effects for garlic (Park et al., 2005; Zeng et al., 2008; El-Shenawy and Hassan, 2008), but the cellular effects of garlic extract on the rat isolated hepatocytes with acetaminophen have not been studied before. Here, for the first time, we have shown that GE is able to prevent ROS formation and GSH depletion caused by APAP administration in hepatocytes freshly isolated from β-naphtoflavon-treated male Sprague-Dawley rats.
MATERIALS AND METHODS
This research project was conducted from 2007 to 2009, in the Drug Applied Research Center and Faculty of Pharmacy in Tabriz University of Medical Sciences.
Chemicals: Bovine Serum Albumin, collagenase A from clostridium histolyticum and HEPES were obtained from Roche diagnostics (Indianapolis, IN) acetaminophen (APAP; 4-acetamidophenol) from Sigma-aldrich (St. Louis, MO), N Acetyl-Cysteine (NAC) from Acros Pharmaceuticals, Di-Chloro-Fluorescin (DCF) and GSH from FLUKA, Beta-Naphto-Flavon (BNF), Heparin sodium salt grade 1-A, Trypan Blue 0.4% solution, Methanol, MgSO4 and other buffer salts, were obtained from Merck (Germany). All other chemicals were of the highest grade commercially available.
Animals: Male Sprague-Dawley rats (200-250 g) were obtained from the laboratory of Animals Research Center of Tabriz University of Medical Sciences. The rats were housed in an air-conditioned room, under controlled temperature of 23±1°C, relative humidity of 36±6% and 12 h light/12 h dark conditions for 1 week before starting the experiments. They were allowed to feed with standard laboratory chow and tap water ad libitum. Procedures involving animals and their care were conducted in conformity with the NIH guidelines for the care and use of laboratory animals. The whole procedure was approved by the Animal Ethics Committee and the Research Council of Vice-Chansellor for Research Affairs at Tabriz University of Medical Sciences, under the license number 5/4/6704 on 2007. 10.28.
Preparation of extract: Fresh garlic (Allium sativum L.) was purchased from a retail food store (Tabriz, Iran) and identified by botanists in the herbarium of Tabriz University. On the day of experiments, the garlic bulbs were peeled, weighed and ground to obtain a fine juice. It was then homogenized in deionized water. The homogenized mixture was filtered through cheesecloth. Garlic extracts of lower concentrations were prepared by dilution of this solution with media used for cell suspensions (Baluchnejadmojarad et al., 2003).
Determination and preparation of allicin from garlic extract: In order to determine and isolate the allicin content in garlic, aqueous extract was assayed using analytical HPLC (Vargas et al., 2008). Separation of allicin from extract was performed using a Spherisorb ODS2 column (4.6x250 mm, 5 μm, waters, Ireland) and methanol (75%) and water (phosphate buffer pH = 3), as a mobile phase, with a flow rate of 1 mL min-1, detecting allicin at the wavelength of 254 nm. Retention time for allicin was 5.8 min. Pure allicin was obtained on the basis of analytical HPLC from a further preparative HPLC. The achieved allicin solution was dried with freeze drier and the resulting allicin powder was used for all experiments. Allicin content of garlic bulbs was quantified with analytical HPLC mentioned above as 80 mg.
Preparation of hepatocytes: Hepatocytes were isolated from male Sprague-Dawley rats by a two-step collagenase perfusion, as described previously (Moldeus et al., 1978; Eghbal et al., 2004). The first step involves the perfusion of a calcium-free buffer. The second step is circulation of a calcium-supplemented buffer containing collagenase. The initial perfusion facilitates desmosomal cleavage and further dispersion of liver cells. The addition of Ca2+ to the enzyme solution ensures adequate collagenase activity. After isolation, the cells were suspended (106 cells mL-1) in Krebs-Henseleit buffer containing 12.5 mM HEPES and incubated under a stream of 95% O2 and 5% CO2 in continuously rotating round-bottomed 50 mL flasks at 37°C. Cell viability was measured by Trypan blue exclusion method. The hepatocytes used in this study were at least 85-90% viable immediately after isolation.
Determination of ROS formation: The dichlorofluorescein-diacetate (DCFH-DA) was dissolved in methanol and was added to the cell suspensions at the same time as APAP. The final concentration of DCFH-DA in cell suspensions was 1 μM. The samples were taken at special time intervals and the fluorescence intensity was measured at the excitation wavelength 485 nm and the emission wavelength of 530 nm. Results were expressed as the fluorescent intensity per 106 cells.
Measurement of intracellular GSH: Intracellular GSH in isolated hepatocytes was measured in deproteinized samples (5% metaphosphoric acid) after derivatization with iodoacetic acid and 1-fluoro-2,4-dinitrobenzene, by HPLC (Reed et al., 1980), using a μBondapak NH2 column (Water associates, Milford, MA).
The procedure is based upon the initial formation of s-carboxymethyl derivatives of free thiols with iodoacetic acid followed by conversion of free amino groups to 2, 4-dinitrophenyl derivatives by reaction with 1-fluoro-2,4-dinitrobenzene (FDNB). Determination of nanomole levels of GSH is possible with this method. Briefly, 0.8 mL of the cell suspension was spun at 50 g for 40 sec and the cell pellet was resuspended in 0.8 mL of fresh medium. 0.2 mL of 25% metaphosphoric acid was added to the sample and after 10-60 min the sample was centrifuged at 100 g for 5 min. 0.5 mL of supernatant and 0.05 mL of iodoacetic acid were mixed in 200-300 mg of sodium bicarbonate. The mixture was sealed and left in the dark and room temperature for 1 h. Then 0.5 mL of FDNB solution (1.5% v/v in ethanol) was added to the sample and the sample was sealed. The sample was left in the dark for 4 h at room temperature and then analyzed by HPLC (Khan and O’Brien, 1997; Eghbal et al., 2004).
Experimental protocols: The animals of test group received 3 I.P injections of BNF (80 mg kg-1) during 72 h before starting the experiments; while the animals of positive control and negative control groups received Corn oil and no chemicals, respectively (Marvasi et al., 2006). On the day of experiments, after induction of anesthesia using sodium pentobarbital, the liver cells were isolated via a two step model. The cells were allowed to get adapted with the incubation conditions for 20 min before the addition of compounds to the incubation mixture. The cells were then exposed to different concentrations of GE and/or NAC 30 min before, at the same time with APAP and 30 min after APAP addition. Aliquots of the cells were taken at different time points (0, 60, 120, 180 and 0, 30, 60, 90, 120 and 150 min) for determination of ROS formation and GSH levels respectively.
Statistical analysis: All the data were expressed as Mean±SEM statistical software SPSS14.0 was used for statistical analysis. All the data were analyzed using one-way Analysis of Variance (ANOVA) for models with repeated measurements, followed by TUKEY’s post hoc tests. The differences were considered significantly at p<0.05 level.
RESULTS
The viability of hepatocytes treated with all chemicals used in this study was examined by trypan blue exclusion method. The results showed that none of the treatments were hepatotoxic, except for APAP which was cytotoxic, when added to the hepatocytes isolated from BNF pre-treated rats, while it did not show any significant hepatotoxicity in rats without pretreatment with BNF. Hepatocytes were incubated in Krebs-Henseleit solution, pH 7.4 at 37°C under the atmosphere of 95% O2/5% CO2. The samples were taken at mentioned time intervals and cell death was assessed by trypan blue exclusion test.
Effects of garlic extract and NAC on acetaminophen-induced ROS formation: We measured the production of ROS in isolated hepatocytes by measuring DCF fluorescence. The probe 2’, 7’-dichlorofluorescin has been used as an indicator of reactive oxygen species formation and oxidative stress (Eghbal et al., 2004). The principle of this assay is that DCFH-DA diffuses through the cell membrane and is enzymatically hydrolyzed by intracellular esterases to nonfluorescent dichlorofluorescin (DCFH). In the presence of ROS, this compound is rapidly oxidized to highly fluorescent dichlorofluorescein (DCF) (LeBel et al., 1992; Eghbal et al., 2004). In order to determine, the possible effects of the chronologic treatments, three protocols were applied. Briefly, after preparation and incubation of cell suspensions, garlic extract and/or NAC was added to suspensions 30 min before, at the same time with and 30 min after addition of APAP solution (500 μM).
The results show significant differences between APAP and control group (Fig. 1, 2) which means APAP could induce a high amount of ROS formation. Figure 1 indicates the protective effect of GE against APAP induced ROS formation; especially when the extract added 30 min before and at the same time with APAP. It can be seen from Fig. 3 that NAC, as the standard treatment, is completely protective even if administered 30 min after APAP.
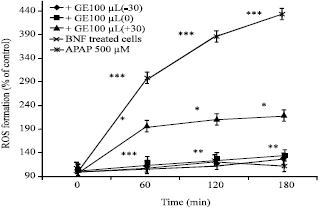 | |
| Fig. 1: | The effect of Garlic Extract on acetaminophen-induced ROS formation (three protocols), ROS formation induced in isolated hepatocytes by APAP. Garlic extracts were added at 3 different time intervals according to the time point of APAP addition. ROS formation was measured at 4 time points. The APAP addition time was equal to 0time point. Values represent Mean±SEM and are at least from 3 independent experiments, *Shows significant difference (p<0.05) with APAP group, **Shows significant difference (p<0.001) with APAP group, ***Shows significant difference (p<0.001) with control group (BNF treated cells) |
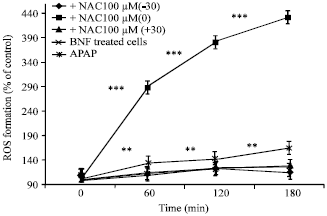 | |
| Fig. 2: | The effects of NAC on acetaminophen-induced ROS formation (three protocols), ROS formation induced by APAP in isolated rat hepatocytes pretreated with BNF. Hepatocytes maintained under the constant flow of 95% O2/5% CO2 .Garlic extracts were added at 3 different time intervals according to the time point of APAP addition. ROS formation was measured at 4 time points. The APAP addition time was equal to 0 time point. Values represent Mean±SEM and are at least from 3 independent experiments, **Shows significant difference (p<0.001) with APAP group, ***Shows significant difference (p<0.001) with control group (BNF treated cells) |
 | |
| Fig. 3: | The effects of APAP, GE and NAC on GSH levels of isolated rat hepatocytes, APAP-induced GSH depletion in isolated rat hepatocytes and protective effects of Garlic extract as well as NAC. Hepatocytes (106 cells mL-1) were isolated from rats pretreated with BNF and maintained under the constant flow of 95% O2/5% CO2 atmosphere. GSH levels were determined at 6 time points according to the time point of APAP addition by the method of Reed et al. (1980). Values represent Mean±SEM and are at least from 3 independent experiments, **Shows significant difference (p<0.05) with APAP group, ***Shows significant difference (p<0.001) with APAP group |
Effects of garlic extract and NAC on acetaminophen-induced GSH depletion: In order to determine the effect of APAP on GSH depletion, GSH content of the hepatocytes was assessed. The standard calibration curve (Fig. 3) was obtained using GSH depleted cells (treated with 200 μM Bromoheptane). Glutathione standard solutions of 4 different concentrations were added to the cell suspensions 30 min after their incubation with bromoheptane. Concentrations used for GSH were 10, 20, 40 and 80 nM.
As shown in Fig. 3, APAP (500 μM) caused a time-dependent GSH depletion in hepatocytes isolated from BNF-pretreated rats with a significant decrease in GSH levels after 60 min of addition. Normal hepatocytes which were isolated from rats without any pretreatment did not show GSH depletion in presence of APAP. Furthermore, there were not any significant differences between 3 timed protocols (described for ROS formation) in GSH levels (data not shown). GE prevented GSH depletion with a high percentage, especially after 60 min which was comparable to the effect of NAC.
DISCUSSION
APAP has been used effectively and safely by a large number of patients for its analgesic and antipyretic effects. At its regular dose in the body, APAP is mainly detoxified either by glucuronide conjugation or sulfation. At overdose, it is also metabolized by CYP2E1 to form NAPQI that depletes GSH. Saturation of the detoxification pathways which causes excess of NAPQI to be formed consequently leading to extensive depletion of GSH and therefore cell death (Burke et al., 2006). Present finding is in accordance with these results as APAP was not able to induce toxicity without the induction of CYP2E1 with BNF. Glutathione (GSH) plays an important role in protecting cells from electrophilic compounds and free radicals such as reactive oxygen species generated during cellular metabolism. Reduced glutathione can act as a reductant, reducing hydrogen peroxide and lipid hydroperoxides directly to H2O, a reaction catalyzed by GSH-Px.
Depletion of intracellular GSH, under conditions of continuous intracellular oxidative stress, leads to oxidation and damage of lipids, proteins and DNA by the reactive oxygen species (Kaplowitz, 2000; Nordberg and Arner, 2001). Oxidative stress (along with nitrosative stress) is one of the proposed mechanisms (Jaeschke, 1990) and depletion of cellular GSH in the liver cells is known to play an important role in APAP toxicity (Mitchell et al., 1973). Glutathione (GSH) is used by cells to detoxify reactive intermediates and is critical for elimination of many drugs, such as APAP. Depletion of GSH can lead to tumor cell death in vitro, especially in cells generating high levels of oxyradicals (Wolchok et al., 2003). It is also well known that NAC is used as an antidote in acetaminophen induced toxicity mainly because it restores the GSH content in the liver (Vina et al., 1980; Mitchell et al., 1985,). In a earlier reported study, Mitchell et al. (1985) investigated GSH status to cellular unity in cultured hepatocytes over time after chemical exposure. Their findings indicate that depletion of GSH to 20% of normal levels qualifies cells for significant drug/chemical induced injury implying that GSH depletion precedes cell toxicity. Previous studies have shown that GSH content decreases after APAP overdose in animal livers (Jaeschke, 1990; Amimoto et al., 1995).
Present results support this idea, because APAP addition caused hepatocytes to lose their GSH content after 30 min. This GSH loss was significantly prevented by GE (p<0.05) and NAC (p<0.001) (Fig. 3). Difference between GE and NAC might be due to the presence of different ingredients in GE which necessitates the isolation and purification of the active compounds. It should be mentioned that, the control group which was just pre-treated with BNF did not show significant decrease in GSH levels during 3 h. This study examines the hypothesis that the toxicity of acetaminophen might occur as a result of ROS formation as well as GSH depletion and that; garlic extract could protect the hepatocytes from these processes. In the present study, administration of AAP resulted in decreased GSH and increased ROS levels indicating that an oxidative stress is present in the cells exposed to AAP.
ROS formation is an important mechanism in various cytotoxicities. The one-electron oxidation of APAP by CYPs may generate Reactive Oxygen Species (ROS). Hydrogen peroxide and superoxide are produced during metabolic activation of APAP in the mixed function oxidase system (Sumioka et al., 2004). It has also been reported that APAP overdose results in a significant decreases in antioxidant enzyme activities such as catalase and glutathione peroxidase (Sumioka et al., 2004). Thus, ROS formation could have a critical role in manifestation of acetaminophen overdose sequelae.
In this study, the amount of ROS formation was shown to increase significantly 60 min after APAP inclusion which is completely inhibited by NAC and to a lesser amount by GE (Fig. 1, 2).
Allicin which is one of the major but unstable components of GE (Amagase et al., 2001) was shown to be cytotoxic (data in-press). Compounds such as allicin which are present in garlic extract can cover the effects of pure antioxidant components of GE like DATS (Zeng et al., 2008) and allyl mercaptan which possess antioxidant activities. The antioxidant and hepatoprotective activities of allyl mercaptan have been studied by our research group and the results will be published in the near future.
Present results are in accordance with the other studies showing garlic to be protective in vivo against hepatotoxicity induced by acetaminophen (Hu et al., 1996) carbon tetrachloride (Fanelli et al., 1998), or cyclophosphamide (Das et al., 1993). Present data indicate that GE is able to protect the cells against consequences of APAP induced cytotoxicity. This protection might be as a result of an ability of garlic components to inhibit phase 1 enzymes or induce phase 2 enzymes, GE antioxidant activity and/or its interaction with NAPQI to protect cellular GSH. Further research is needed to clarify the subject.
ACKNOWLEDGMENTS
This study was supported by Drug Applied and Biotechnology research centers in Tabriz University of Medical Sciences, Tabriz, Iran. The authors want to thank Dr. S. Dastmalchi for his kind supports.
REFERENCES
- Agarwal, K.C., 1996. Therapeutic actions of garlic constituents. Med. Res. Rev., 16: 111-124.
PubMedDirect Link - Aleksunes, L.M., S.N. Campion, M.J. Goedken and J.E. Manautou, 2008. Acquired resistance to acetaminophen hepatotoxicity is associated with induction of multidrug resistance-associated protein 4 (Mrp4) in proliferating hepatocytes. Toxicol. Sci., 104: 261-273.
Direct Link - Amagase, H., B.L. Petesch, H. Matsuura, S. Kasuga and Y. Itakura, 2001. Intake of garlic and its bioactive components. J. Nutr., 131: 955S-962S.
PubMedDirect Link - Amagase, H., 2006. Clarifying the real bioactive constituents of garlic. J. Nutr., 136: 716S-725S.
PubMedDirect Link - Amar, P.J. and E.R. Schiff, 2007. Acetaminophen safety and hepatotoxicity--where do we go from here? Exp. Opin. Drug Saf., 6: 341-355.
PubMed - Amimoto, T., T. Matsura, S.Y. Koyama, T. Nakanishi, K. Yamada and G. Kajiyama, 1995. Acetaminophen-induced hepatic injury in mice: The role of lipid peroxidation and effects of pretreatment with coenzyme Q10 and alpha-tocopherol. Free Radic. Biol. Med., 19: 169-176.
CrossRef - Banerjee, S.K., S. Sood, A.K. Dinda, T.K. Das and S.K. Maulik, 2003. Chronic oral administration of raw garlic protects against isoproterenol-induced myocardial necrosis in rat. Comp. Biochem. Physiol. C Toxicol. Pharmacol., 136: 377-386.
PubMed - Das, T., A. Royshoudhury, A. Sharma and G. Talukder, 1993. Modification of clastogenicity of three known clastogens by garlic extract in mice in vivo. Environ. Mol. Mutagen., 21: 383-388.
CrossRefDirect Link - El-Shenawy, S.M.A. and N.S. Hassan, 2008. Comparative evaluation of the protective effect of selenium and garlic against liver and kidney damage induced by mercury chloride in the rats. Pharmacol. Rep., 60: 199-208.
PubMed - Fanelli, S.L., G.D. Castro, E.G. De Toranzo and J.A. Castro, 1998. Mechanisms of the preventive properties of some garlic components in the carbon tetrachloride-promoted oxidative stress. Diallyl sulfide; diallyl disulfide; allyl mercaptan and allyl methyl sulfide. Res. Commun. Mol. Pathol. Pharmacol., 102: 163-174.
PubMedDirect Link - Hu, J.J., J.S. Yoo, M. Lin, E.J. Wang and C.S. Yang, 1996. Protective effects of diallyl sulfide on acetaminophen-induced toxicities. Food Chem. Toxicol., 34: 963-969.
PubMedDirect Link - Jaeschke, H., 1990. Glutathione disulfide formation and oxidant stress during acetaminophen-induced hepatotoxicity in mice in vivo: The protective effect of allopurinol. J. Pharmacol. Exp. Ther., 255: 935-941.
PubMedDirect Link - Khan, S. and P.J. O'Brien, 1997. Rapid and specific efflux of glutathione before hepatocyte injury induced by hypoxia. Biochem. Biophys. Res. Commun., 238: 320-322.
CrossRef - LeBel, C.P., H. Ischiropoulos and S.C. Bondy, 1992. Evaluation of the probe 2',7' dichlorofluorescin as an indicator of reactive oxygen species formation and oxidative stress. Chem. Res. Toxicol., 5: 227-231.
PubMedDirect Link - Marvasi, L., A. Zaghini, P.G. Geravasi, E. Vaccaro and V. Chirulli, 2006. Induction of cytochrome P450 1A by beta-naphto-flavon within the pig liver and central nervous system. Vet. Res. Commun., 30: 333-336.
CrossRef - Mitchell, J.R., D.J. Jollow, W.Z. Potter, J.R. Gillette and B.B. Brodie, 1973. Acetaminophen-induced hepatic necrosis: IV. Protective role of glutathione. J. Pharmacol. Exp. Ther., 187: 211-217.
PubMed - Mitchell, D.B., D. Acosta and J.V. Bruckner, 1985. Role of Glutathione depletion in the cytotoxicity of Acetaminophen in a primary culture system of rat hepatocytes. Toxicology, 37: 127-146.
PubMedDirect Link - Nordberg, J. and E.S.J. Arner, 2001. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system. Free Radic. Biol. Med., 31: 1287-1312.
CrossRefPubMedDirect Link - Moldeus, P., J. Hogberg and S. Orrenius, 1978. Isolation and use of liver cells. Methods Enzymol., 52: 60-71.
PubMedDirect Link - Park, E.Y., S.H. Ki, M.S. Ko, C.W. Kim, M.H. Lee, Y.S. Lee and S.G. Kim, 2005. Garlic oil and DDB, comprised in a pharmaceutical composition for the treatment of patients with viral hepatitis, prevents acute liver injuries potentiated by glutathione deficiency in rats. Chem. Biol. Interact., 155: 82-96.
PubMed - Reed, D.J., J.R. Babson, P.W. Beatty, A.E. Brodie, W.W. Ellis and D.W. Potter, 1980. High-performance liquid chromatography analysis of nanomole levels of glutathione, glutathione disulfide and related thiols and disulfides. Anal. Biochem., 106: 55-62.
CrossRef - Ruepp, S.U., R.P. Tonge, J. Shaw, N. Wallis and F. Pognan, 2001. Genomics and proteomics analysis of acetaminophen toxicity in mouse liver. Toxicol. Sci., 65: 135-150.
PubMed - Sumioka, I., T. Matsura and K. Yamada, 2004. Acetaminophen induced hepatotoxicity: Still an important issue. Yonago Acta Med., 47: 17-28.
Direct Link - Vargas-Arispuro, I., C. Corrales-Maldonado and M.A. Martynez-Tellez, 2008. Compounds derived from garlic as bud induction agents in organic farming of table grape. Chilean J. Agric. Res., 68: 94-101.
Direct Link - Wolchok, J.D., L. Williams, J.T. Pinto, M. Fleisher, S.E. Krown, W.J. Hwu, P.Q. Livingston, C. Chang and P.B. Chapman, 2003. Phase I trial of high dose paracetamol and carmustine in patients with metastatic melanoma. Melanoma Res., 13: 189-196.
PubMed - Xu, S. and B.H. Simon Cho, 2000. Allyl mercaptan, a major metabolite of garlic compounds, reduces cellular cholesterol synthesis and its secretion in Hep-G2 cells. J. Nutr. Biochem., 10: 654-659.
PubMed - Zeng, T., C.L. Zhang, Z.P. Zhu, L.H. Yu, X.L. Zhao and Q.K. Xie, 2008. Diallyl trisulfide (DATS) effectively attenuated oxidative stress-mediated liver injury and hepatic mitochondrial dysfunction in acute ethanol-exposed mice. Toxicology, 252: 86-91.
PubMed
Fakour Ahmadiani Reply
This is an acceptible paper in PJBS. I will be glad to send papers to PJBS with the qualified papers like this one
abd akrom Reply
good, qualified and applicable for us
sadollah mohammadi Reply
I studied this basic and experimental article. In my opinion, it is a useful and applied research in this simple but life- threatening subject about Acetaminophen toxicity.and ,I hope ,with continuity trying and indefatigable preservance of article authors and other researchers , for solving an important problem at the medical and therapeutics events ,the results of this research could enter as Pharmaceutical Products to the drug marketing in near future